Antigenic and Pathogenic Characteristics of QX-Type Avian Infectious Bronchitis Virus Strains Isolated in Southwestern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Eggs
2.2. Phylogenetic and Recombination Analysis of S1 Genes
2.3. Antigenic Analysis of Different QX-Type Strains
2.3.1. Preparation of Chicken Antisera against IBVs
2.3.2. Adaption of IBVs in Chicken Embryo Kidney (CEK) Cells
2.3.3. Virus Cross-Neutralization Tests
2.3.4. Antigenic Relatedness Analysis
2.4. Primer Design and Quantitative Real-Time RT-PCR Validation (RT-qPCR)
2.4.1. Primer Design
2.4.2. Standard Plasmid
2.4.3. RT-qPCR Assay
2.5. Pathogenicity Tests
2.5.1. Schedule of Challenge
2.5.2. Clinical Signs and Tissue Lesion
2.5.3. Virus Load Quantification
2.6. Statistical Analysis
2.7. Ethics Statement
3. Results
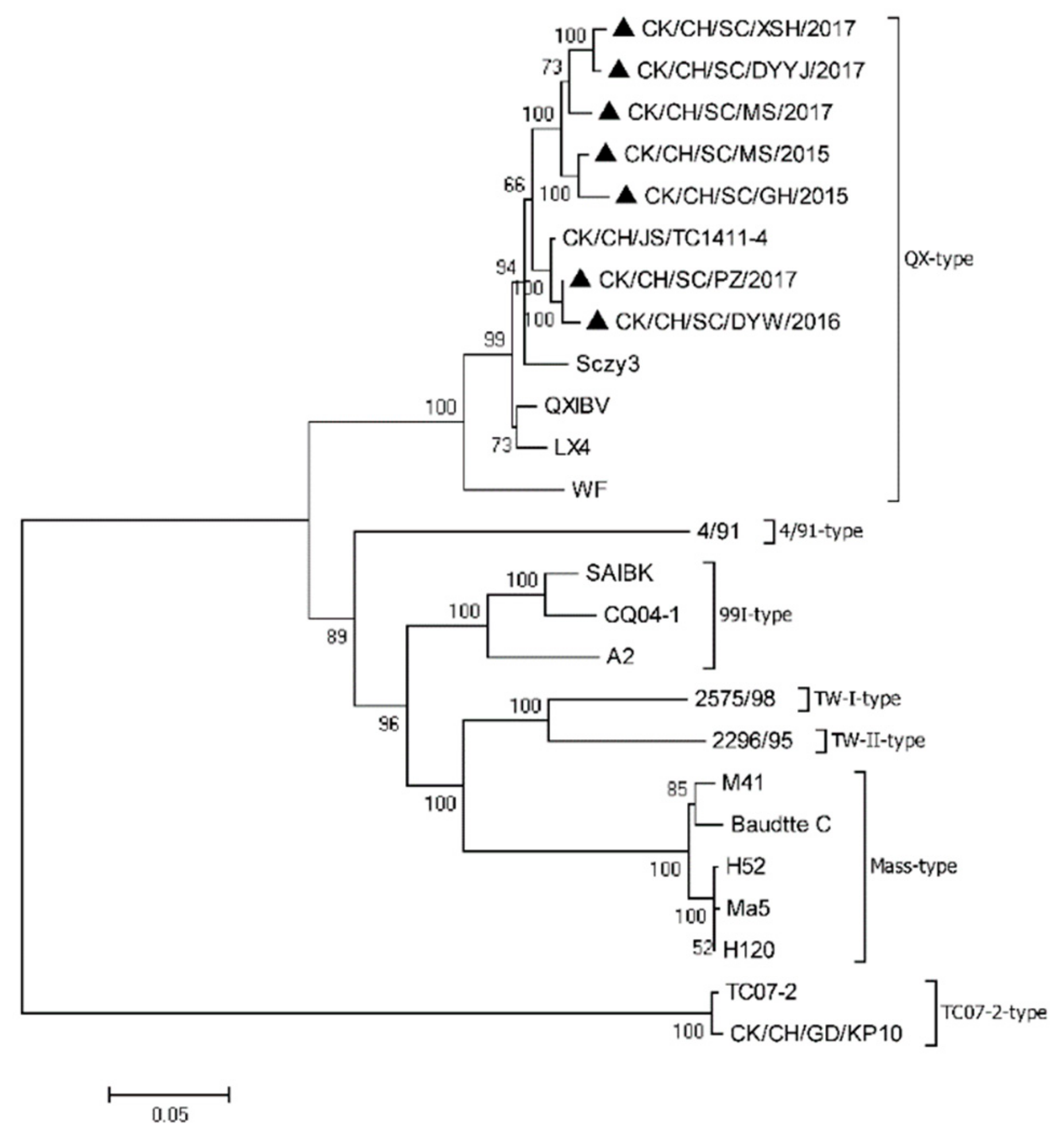
3.1. Phylogenetic and Recombination Analysis of the S1 Gene
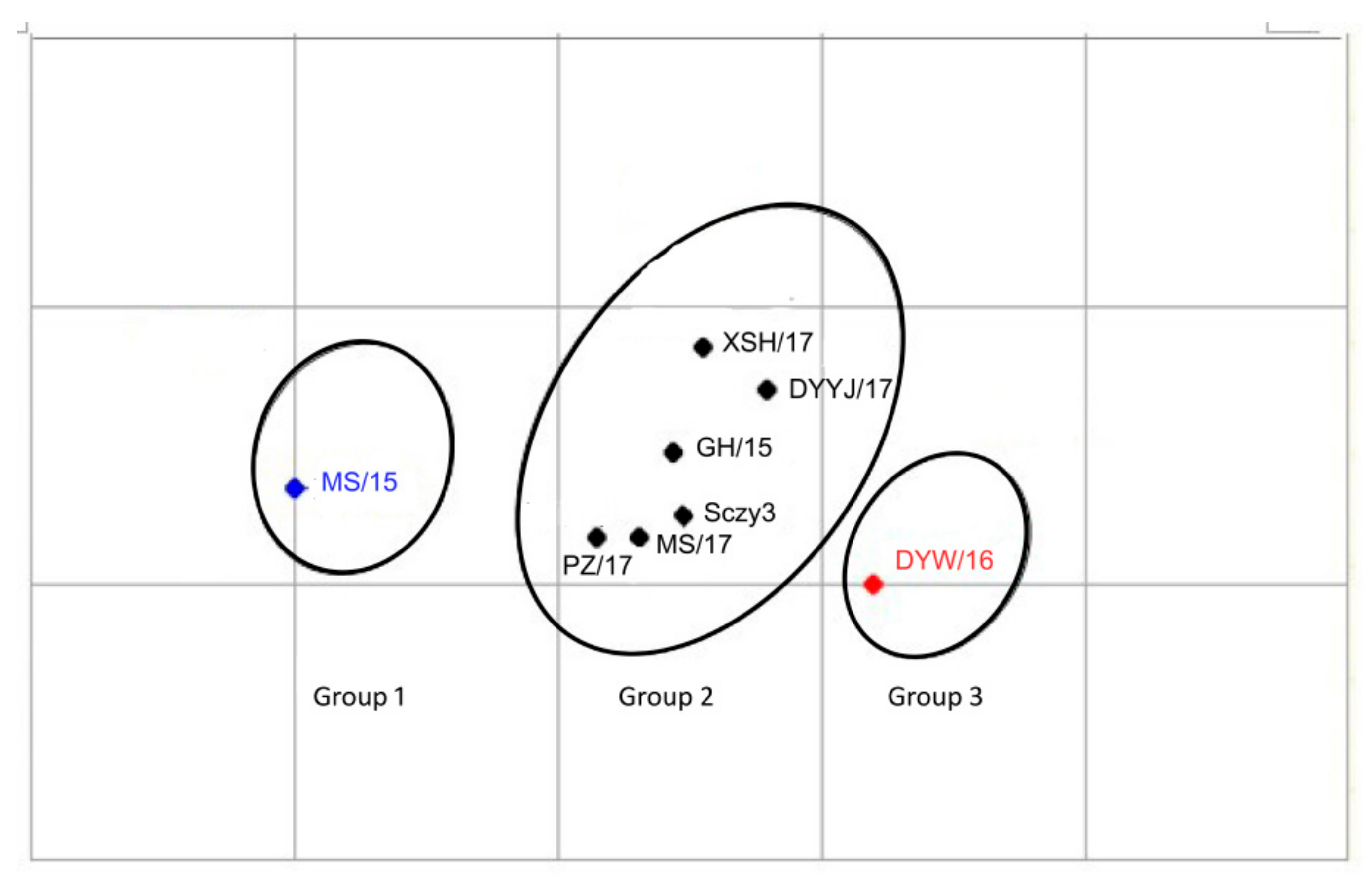
3.2. Antigenic Analysis of the QX-Type Strains
3.3. Establishment of a Standard Curve
3.4. Pathogenicity Tests
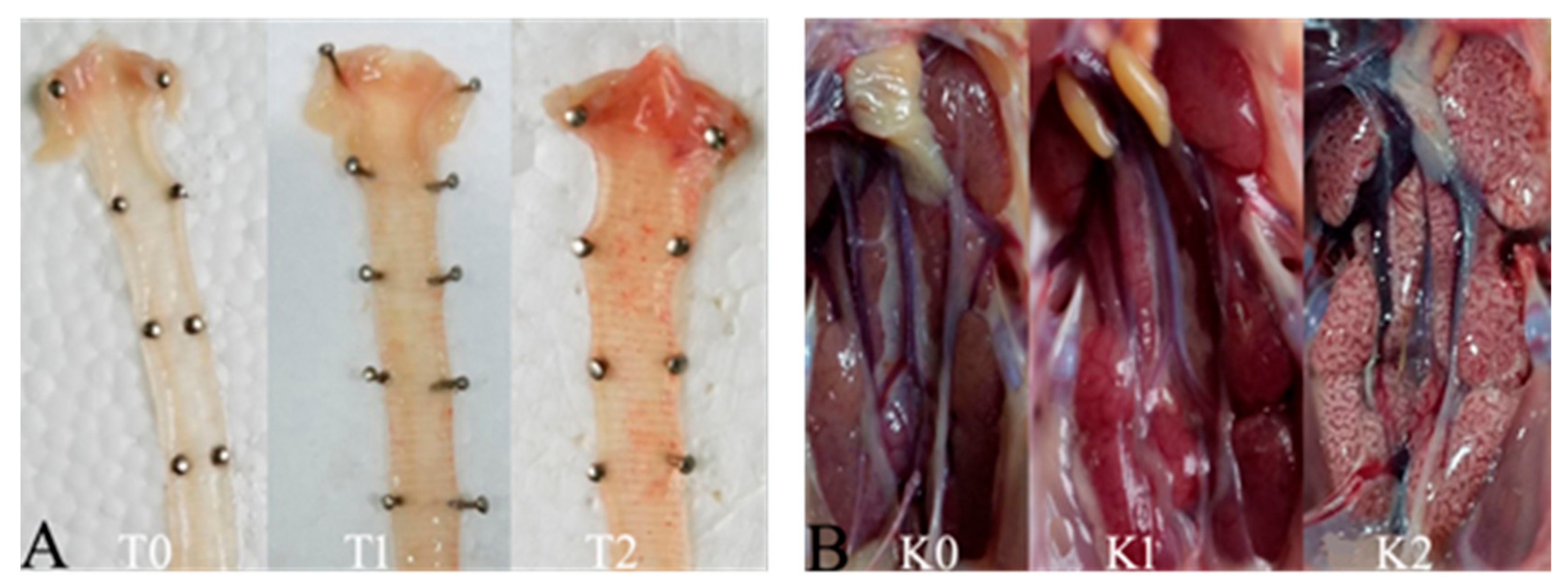
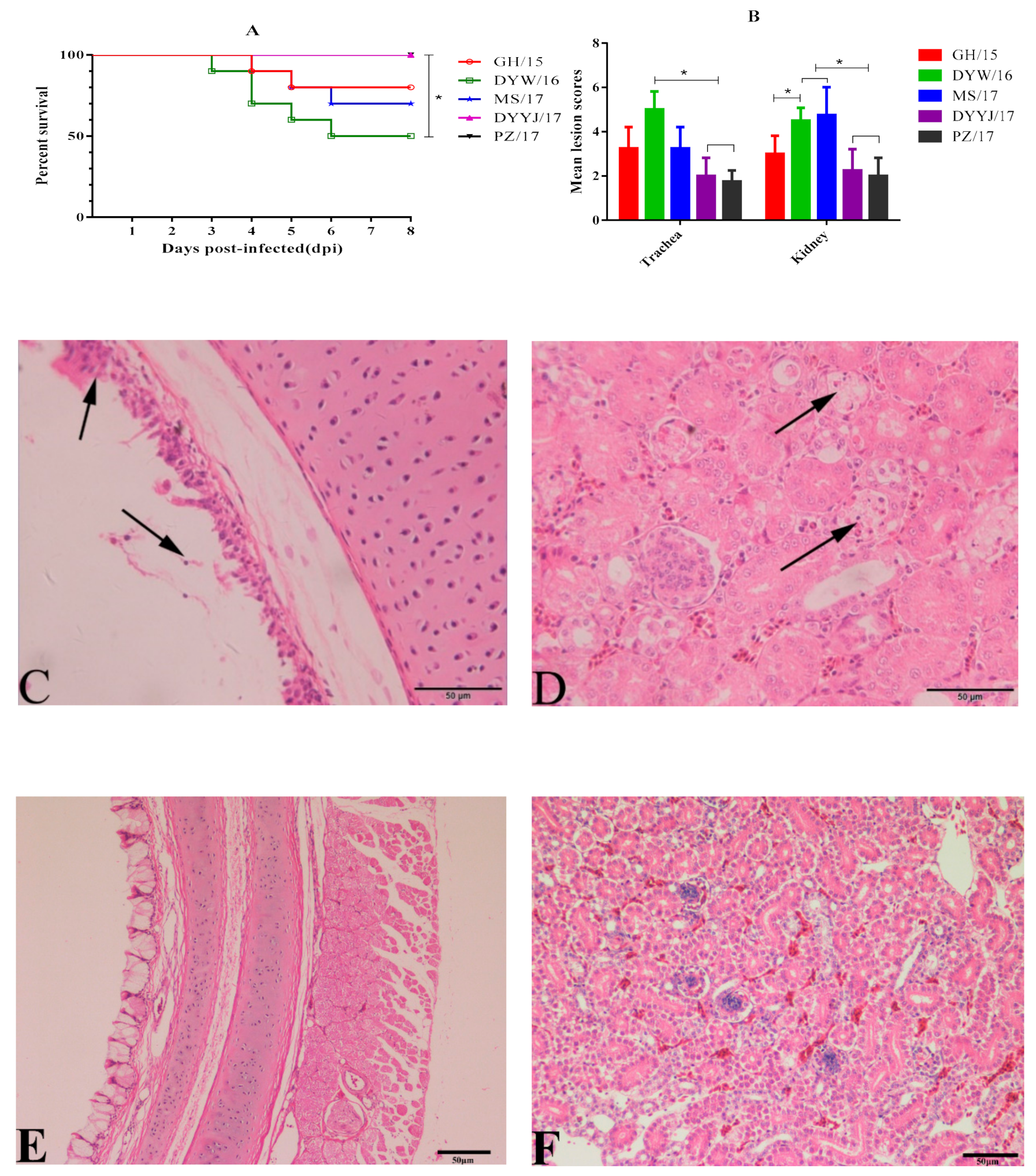
3.4.1. Clinical Signs
3.4.2. Gross and Microscopic Lesions
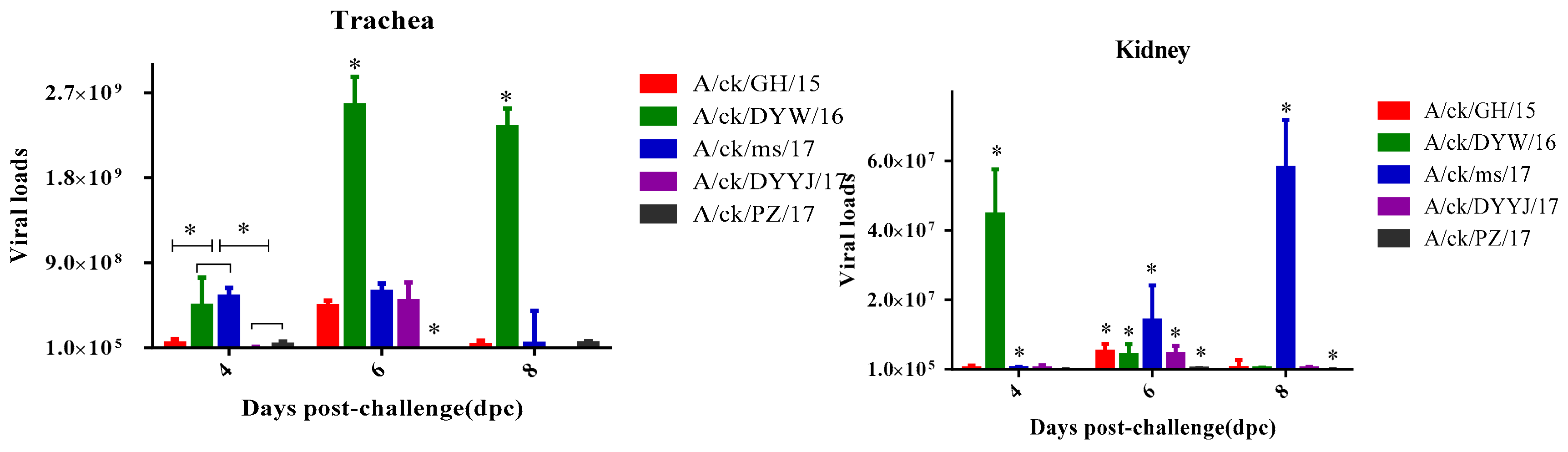
3.4.3. Viral Load in the Trachea and Kidneys
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Abu-Akkada, S.S.; Awad, A.M. Isolation, propagation, identification and comparative pathogenicity of five Egyptian field strains of Eimeriatenella from broiler chickens in five different provinces in Egypt. Res. Vet. Sci. 2012, 92, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Colvero, L.P.; Villarreal, L.Y.; Torres, C.A.; Brañdo, P.E. Assessing the economic burden of avian infectious bronchitis on poultry farms in Brazil. Rev. Sci. Tech. 2015, 34, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Zhang, M.; Xue, T.; Shao, H.; Qian, K.; Ye, J.; Qin, A. Identification of a novel recombinant virulent avian infectious bronchitis virus. Vet. Microbiol. 2017, 199, 120. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cheng, J.L.; Liu, X.Y.; Zhao, J.; Hu, Y.X.; Zhang, G.Z. Safety and efficacy of an attenuated Chinese QX-like infectious bronchitis virus strain as a candidate vaccine. Vet. Microbiol. 2015, 180, 49–58. [Google Scholar] [CrossRef]
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1996, 142, 629–633. [Google Scholar]
- Li, M.; Wang, X.Y.; Wei, P.; Wei, Z.J.; Mo, M.L. Serotype and genotype diversity of infectious bronchitis viruses isolated during 1985-2008 in Guangxi, China. Arch. Virol. 2012, 157, 467–474. [Google Scholar] [CrossRef]
- Cavanagh, D.; Davis, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus IBV: Virus retaining spike glycopolypeptide S2 but not S1 is unable to induce virus-neutralizing or haemagglutination-inhibiting antibody, or induce chicken tracheal protection. J. Gen. Virol. 1986, 67 Pt 7, 1435–1442. [Google Scholar] [CrossRef]
- Parsons, L.M.; Bouwman, K.M.; Azurmendi, H. Glycosylation of the viral attachment protein of avian coronavirus is essential for host cell and receptor binding. J. Biol. Chem. 2019, 294, 7797–7809. [Google Scholar] [CrossRef] [Green Version]
- Promkuntod, N.; Eijndhoven, R.E.W.V.; Vrieze, G.D.; Gröne, A.; Verheije, M.H. Mapping of the receptor-binding domain and amino acids critical for attachment in the spike protein of avian coronavirus infectious bronchitis virus. Virology 2014, 448, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Shan, D.; Fang, S.; Han, Z.; Ai, H.; Zhao, W.; Chen, Y.; Jiang, L.; Liu, S. Effects of Hypervariable Regions in Spike Protein on Pathogenicity, Tropism, and Serotypes of Infectious Bronchitis Virus. Virus Res. 2018, 250, 104–113. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, J.; Yan, S.; Jia, W.; Zhang, K.; Zhang, G. S gene and 5a accessory gene are responsible for the attenuation of virulent infectious bronchitis coronavirus. Virology 2019, 533, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Qin, J.; Chen, F.; Xie, Q.; Bi, Y.; Cao, Y.; Xue, C. Phylogenetic analysis of the S1 glycoprotein gene of infectious bronchitis viruses isolated in China during 2009–2010. Virus Genes 2012, 44, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; He, X.; Yao, K.C.; Du, L.J.; Liu, P.; Yan, Q.G.; Wen, Y.P.; Cao, S.J.; Han, X.F.; Huang, Y. Phylogenetic and antigenic analysis of avian infectious bronchitis virus in southwestern China, 2012–2016. Infect. Genet. Evol. 2016, 45, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Zhao, J.; Zhong, Q.; Zhang, G.Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566. [Google Scholar] [CrossRef] [PubMed]
- Zou, N.L.; Zhao, F.F.; Wang, Y.P.; Liu, P.; Cao, S.J.; Wen, X.T.; Huang, Y. Genetic analysis revealed LX4 genotype strains of avian infectious bronchitis virus became predominant in recent years in Sichuan area, China. Virus Genes 2010, 41, 202–209. [Google Scholar] [CrossRef]
- Abro, S.H.; Ullman, K.; Belák, S.; Baule, C. Characterization and analysis of the full-length genome of a strain of the European QX-like genotype of infectious bronchitis virus. Arch. Virol. 2012, 157, 1211–1215. [Google Scholar] [CrossRef]
- Pohuang, T.; Sasipreeyajan, J. The pathogenesis of a new variant genotype and QX-like infectious bronchitis virus isolated from chickens in Thailand. Thai J. Vet. Med. 2012, 42, 51–57. [Google Scholar]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and Challenges toward the Development of Vaccines against Avian Infectious Bronchitis. J. Immunol. Res. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Liu, X.; Zhao, J.; Xu, G.; Zhao, Y.; Zhang, G. Analysis of antigenicity and pathogenicity reveals major differences among QX-like infectious bronchitis viruses and other serotypes. Vet. Microbiol. 2017, 203, 167–173. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Senapathi, U.D.S.; Abdul-Cader, M.S.; Popowich, S.; Marshall, F.; Cork, S.C.; Meer, F.V.D.; Gomis, S.; Abdul-Careem, M.F. Comparative features of infections of two Massachusetts (Mass) infectious bronchitis virus (IBV) variants isolated from Western Canadian layer flocks. BMC Vet. Res. 2018, 14, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406. [Google Scholar] [PubMed]
- Xia, J.; He, X.; Du, L.J.; Liu, Y.Y.; You, G.J.; Li, S.Y.; Liu, P.; Cao, S.J.; Han, X.F.; Huang, Y. Preparation and protective efficacy of a chicken embryo kidney cell-attenuation GI-19/QX-like avian infectious bronchitis virus vaccine. Vaccine 2018, 36, 4087–4094. [Google Scholar] [CrossRef] [PubMed]
- Gravendyck, M.; Tritt, S.; Spenkochâ-Piper, H.; Kaleta, E.F. Antigenic diversity of psittacine herpesviruses: Cluster analysis of antigenic differences obtained from crossâ-neutralization tests. Avian Pathol. 1996, 25, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.M.; Zhao, J.; Xue, J.; Yang, Y.L.; Zhang, G.Z. Antigenic variation of LaSota and genotype VII Newcastle disease virus (NDV) and their efficacy against challenge with velogenic NDV. Vaccine 2017, 35, 27. [Google Scholar] [CrossRef]
- Lamar, B.J.; Jialiang, Y.; Zhipeng, C.; Tong, Z.; Xiu-Feng, W. AntigenMap 3D: An online antigenic cartography resource. Bioinformatics 2012, 28, 1292–1293. [Google Scholar]
- Wei, Z.J.; Wang, F.Y.; Guo, M.P.; Duan, Z.Z.; Zou, N.L.; Liu, P.; Yan, Q.G.; Wen, X.T.; Cao, S.J.; Huang, Y. Dynamic changes of virus load in supernatant of primary CEK cell culture infected with different generations of avian infectious bronchitis virus strains Sczy3 as revealed by real-time reverse transcription-polymerase chain reaction. Genet. Mol. Res. 2015, 14, 6340–6349. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Z.; Hao, H.; Cheng, S.; Fan, W.; Du, E.; Xiao, S.; Wang, X.; Zhang, S. Development of a SYBR Green real-time RT-PCR assay for the detection of avian encephalomyelitis virus. J. Virol. Methods 2014, 206, 46–50. [Google Scholar] [CrossRef]
- Alexander, D.J.; Collins, M.S. Effect of pH on the growth and cytopathogenicity of avian infectious bronchitis virus in chick kidney cells. Arch. Virol. 1975, 49, 339. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, H. Sequence analysis of nephropathogenic infectious bronchitis virus strains of the Massachusetts genotype in Beijing. Avian Pathol. 2001, 30, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Zou, N.; Xia, J.; Wang, F.; Duan, Z.; Miao, D.; Yan, Q.; Cao, S.; Wen, X.; Liu, P.; Huang, Y. Two novel neutralizing antigenic epitopes of the s1 subunit protein of a QX-like avian infectious bronchitis virus strain Sczy3 as revealed using a phage display peptide library. Vet. Immunol. Immunopathol. 2015, 168, 0165242715001828. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.M.; Saif, L.J.; Marthaler, D.; Wang, Q. Evolution, antigenicity and pathogenicity of global porcine epidemic diarrhea virus strains. Virus Res. 2016, 226, 20–39. [Google Scholar] [CrossRef] [Green Version]
- Leyson, C.; Franca, M.; Jackwood, M.; Jordan, B. Polymorphisms in the S1 spike glycoprotein of Arkansas-type infectious bronchitis virus (IBV) show differential binding to host tissues and altered antigenicity. Virology 2016, 498, 218–225. [Google Scholar] [CrossRef]
- Wickramasinghe, I.N.A.; De Vries, R.P.; Gröne, A.; De Haan, C.A.M.; Verheije, M.H. Binding of avian coronavirus spike proteins to host factors reflects virus tropism and pathogenicity. J. Virol. 2011, 85, 8903–8912. [Google Scholar] [CrossRef] [Green Version]
- Reddy, V.R.A.P.; Trus, I.; Desmarets, L.M.B.; Li, Y.; Theuns, S.; Nauwynck, H.J. Productive replication of nephropathogenic infectious bronchitis virus in peripheral blood monocytic cells, a strategy for viral dissemination and kidney infection in chickens. Vet. Res. 2016, 47, 70. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, R.; Kuchipudi, S.V.; Chantrey, J.; Ganapathy, K. Pathogenicity and tissue tropism of infectious bronchitis virus is associated with elevated apoptosis and innate immune responses. Virology 2016, 488, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Maria, A.; Dave, C.; Paul, B. The replicase gene of avian coronavirus infectious bronchitis virus is a determinant of pathogenicity. PLoS ONE 2009, 4, e7384. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain. | Year of Isolation | Chicken Type | Location | Symptoms Phenotype | Abbreviations |
|---|---|---|---|---|---|
| CK/CH/SC/GH/2015 | 2015 | Broiler | Sichuan (Guanghan) | Respiratory | GH/15 |
| CK/CH/SC/MS/2015 | 2015 | Broiler | Sichuan (Chengdu) | Respiratory | MS/15 |
| CK/CH/SC/DYW/2016 | 2016 | Broiler | Sichuan (Deyang) | Respiratory; Nephritis | DYW/16 |
| CK/CH/SC/XSH/2017 | 2017 | Broiler | Sichuan (Deyang) | Respiratory; Nephritis | XSH/17 |
| CK/CH/SC/PZ/2017 | 2017 | Broiler | Sichuan (Penzhou) | Respiratory | PZ/17 |
| CK/CH/SC/MS/2017 | 2017 | Broiler | Sichuan (Meishan) | Respiratory; Nephritis | MS/17 |
| CK/CH/SC/DYYJ/2017 | 2017 | Broiler | Sichuan (Deyang) | Respiratory | DYYJ/17 |
| Virus | Antiserum | |||||||
|---|---|---|---|---|---|---|---|---|
| Sczy3 | MS/15 | GH/15 | DYW/16 | XSH/17 | MS/17 | DYYJ/17 | PZ/17 | |
| Sczy3 | 1240 | 178 | 178 | 1995 | 45 | 501 | 1412 | 178 |
| MS/15 | 1240 | 1412 | 708 | 501 | 708 | 356 | 708 | 1000 |
| GH/15 | 625 | 178 | 178 | 501 | 708 | 501 | 5623 | 89 |
| DYW/16 | 625 | 89 | 32 | 1000 | 251 | 178 | 1412 | 89 |
| XSH/17 | 1240 | 251 | 178 | 1995 | 501 | 708 | 2818 | 126 |
| MS/17 | 870 | 708 | 126 | 251 | 356 | 356 | 1412 | 63 |
| DYYJ/17 | 1240 | 708 | 45 | 2818 | 251 | 251 | 2818 | 126 |
| PZ/17 | 1240 | 126 | 708 | 708 | 89 | 501 | 1000 | 126 |
| Virus | Antiserum | |||||||
|---|---|---|---|---|---|---|---|---|
| Sczy3 | MS/15 | GH/15 | DYW/16 | XSH/17 | MS/17 | DYYJ/17 | PZ/17 | |
| Sczy3 | 1.00 | 0.38 | 0.71 | 0.60 | 0.28 | 0.99 | 0.71 | 1.18 |
| MS/15 | 1.00 | 0.71 | 0.17 | 0.50 | 0.71 | 0.35 | 0.84 | |
| GH/15 | 1.00 | 0.30 | 1.18 | 0.99 | 0.71 | 1.68 | ||
| DYW/16 | 1.00 | 1.00 | 0.35 | 1.19 | 0.71 | |||
| XSH/17 | 1.00 | 1.19 | 0.70 | 0.42 | ||||
| MS/17 | 1.00 | 0.60 | 0.84 | |||||
| DYYJ/17 | 1.00 | 0.59 | ||||||
| PZ/17 | 1.00 | |||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Du, L.; Xia, J.; Du, J.; You, G.; Wen, Y.; Huang, X.; Zhao, Q.; Han, X.; Yan, Q.; et al. Antigenic and Pathogenic Characteristics of QX-Type Avian Infectious Bronchitis Virus Strains Isolated in Southwestern China. Viruses 2019, 11, 1154. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121154
Li S, Du L, Xia J, Du J, You G, Wen Y, Huang X, Zhao Q, Han X, Yan Q, et al. Antigenic and Pathogenic Characteristics of QX-Type Avian Infectious Bronchitis Virus Strains Isolated in Southwestern China. Viruses. 2019; 11(12):1154. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121154
Chicago/Turabian StyleLi, Shuyun, Lijing Du, Jing Xia, Jiteng Du, Guojin You, Yiping Wen, Xiaobo Huang, Qing Zhao, Xinfeng Han, Qigui Yan, and et al. 2019. "Antigenic and Pathogenic Characteristics of QX-Type Avian Infectious Bronchitis Virus Strains Isolated in Southwestern China" Viruses 11, no. 12: 1154. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121154