Tracing the Lineage of Two Traits Associated with the Coat Protein of the Tombusviridae: Silencing Suppression and HR Elicitation in Nicotiana Species

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculation of Nicotiana Species with TNV Virions
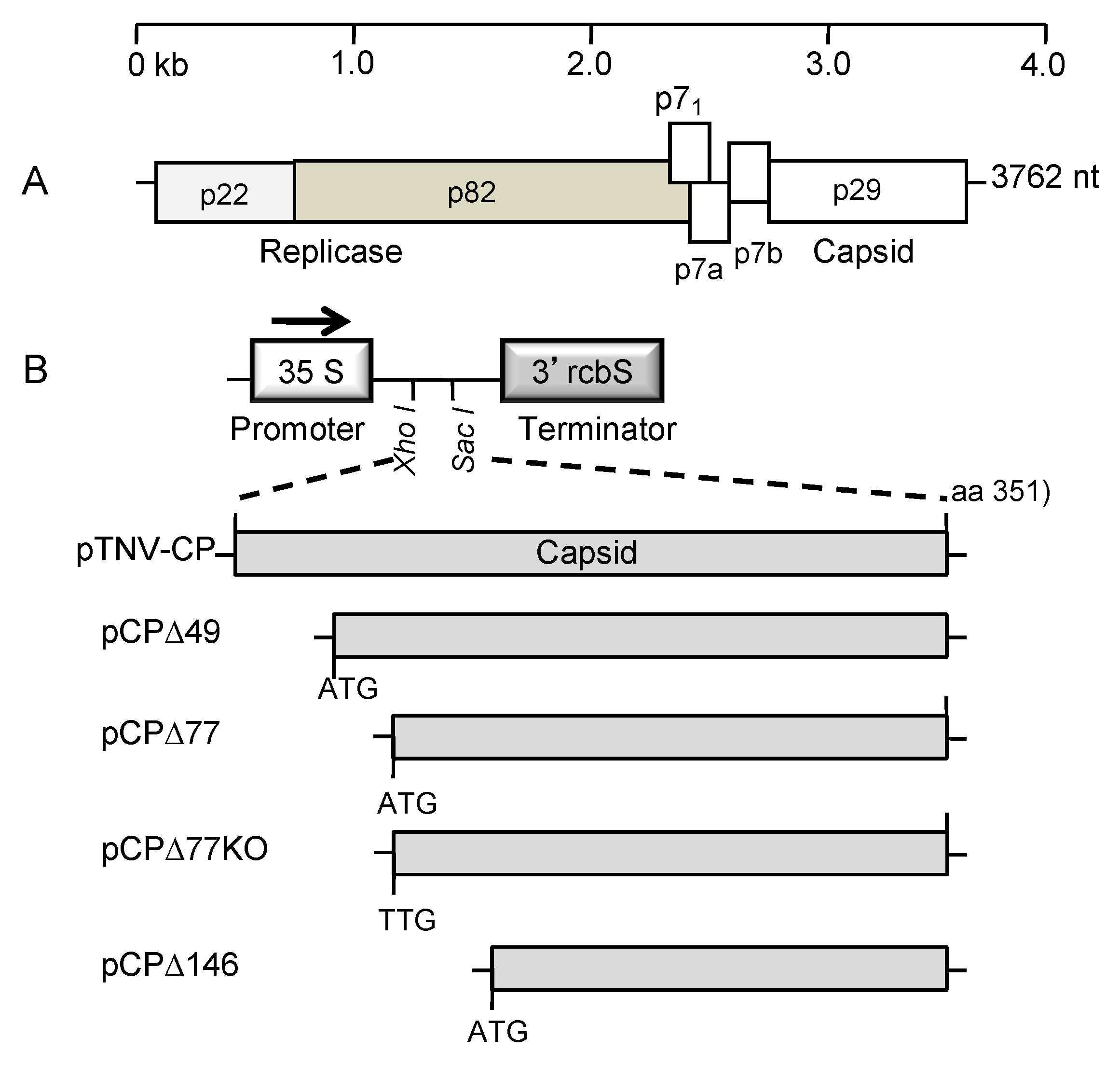
2.2. Coat Protein Constructs
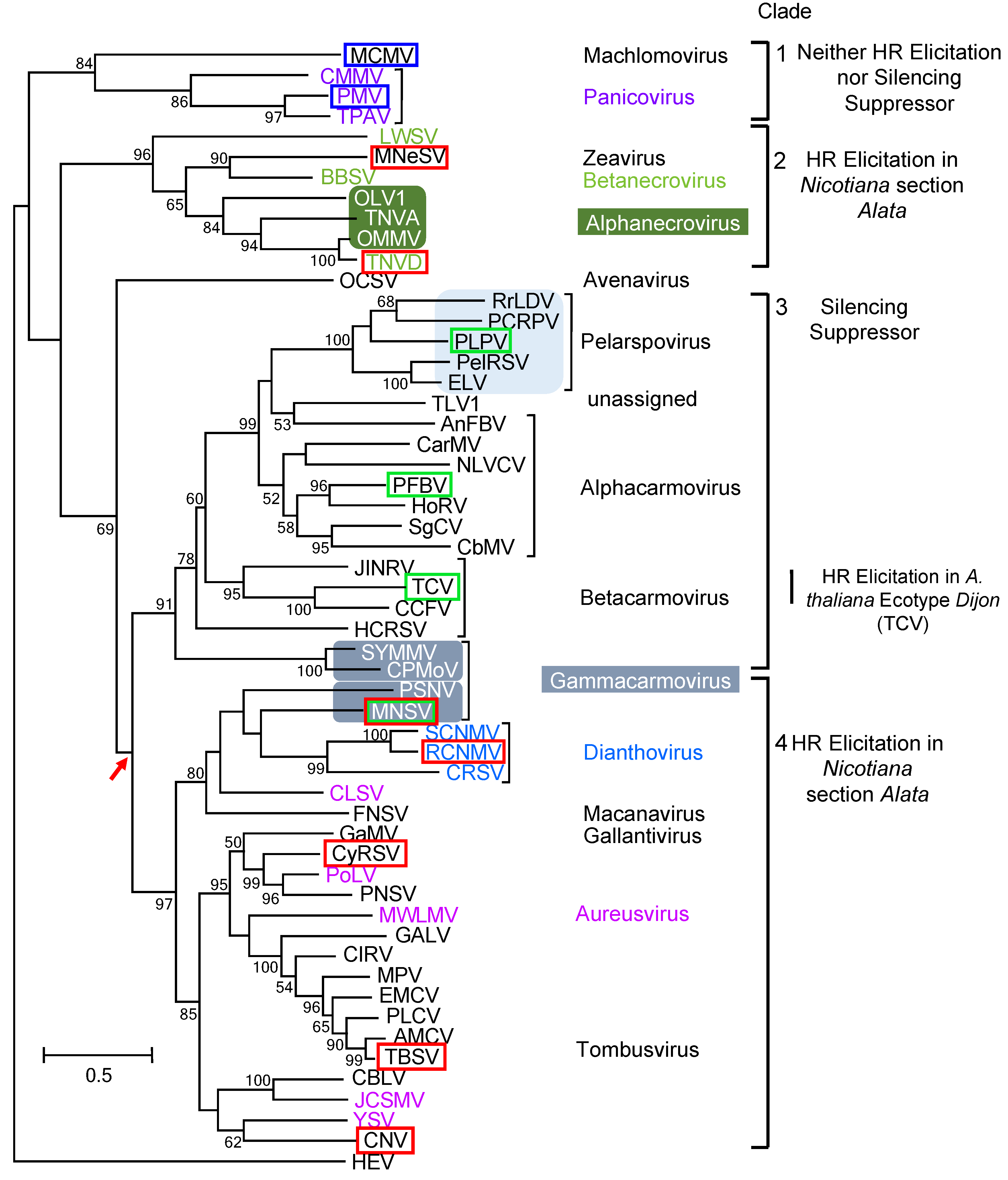
2.3. Phylogenetic Analysis of the Tombusvirid CPs
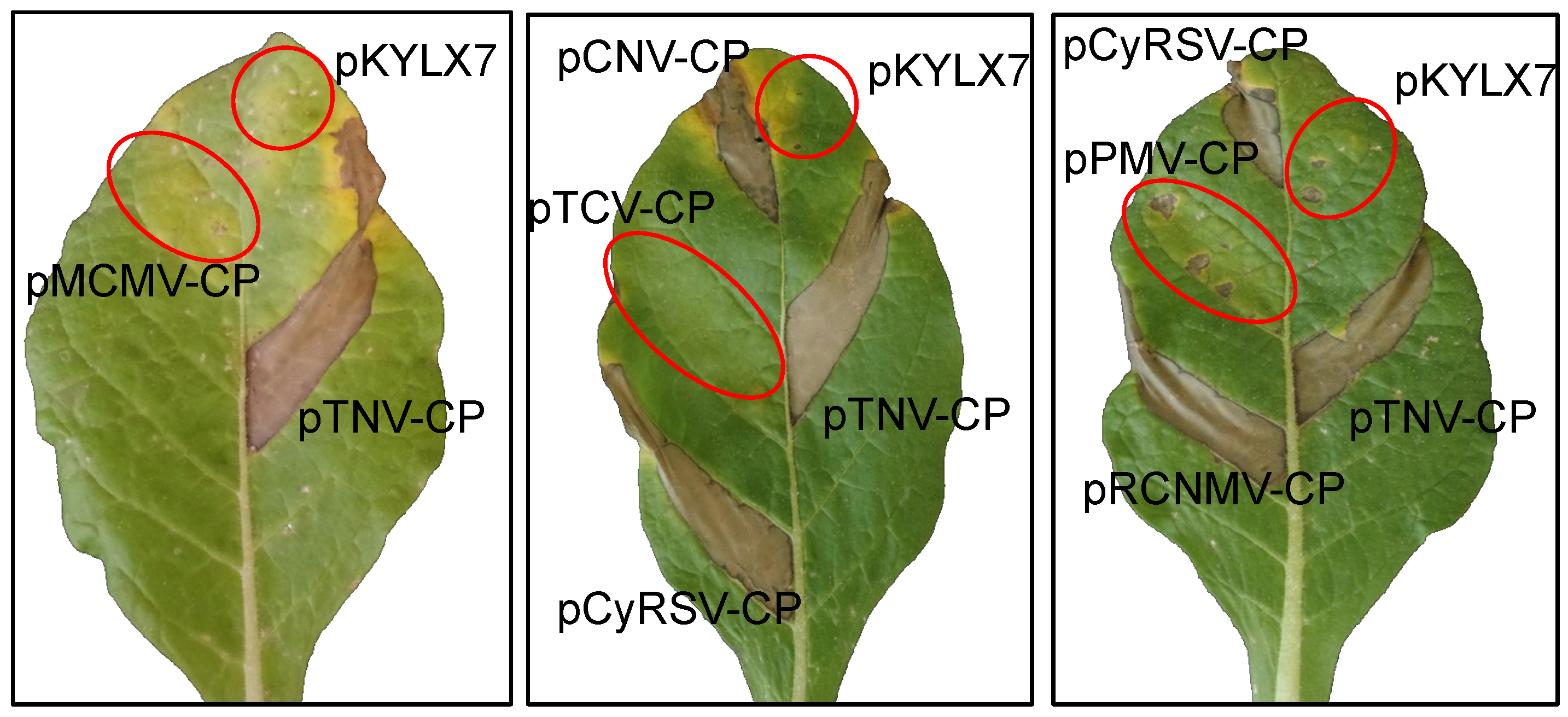
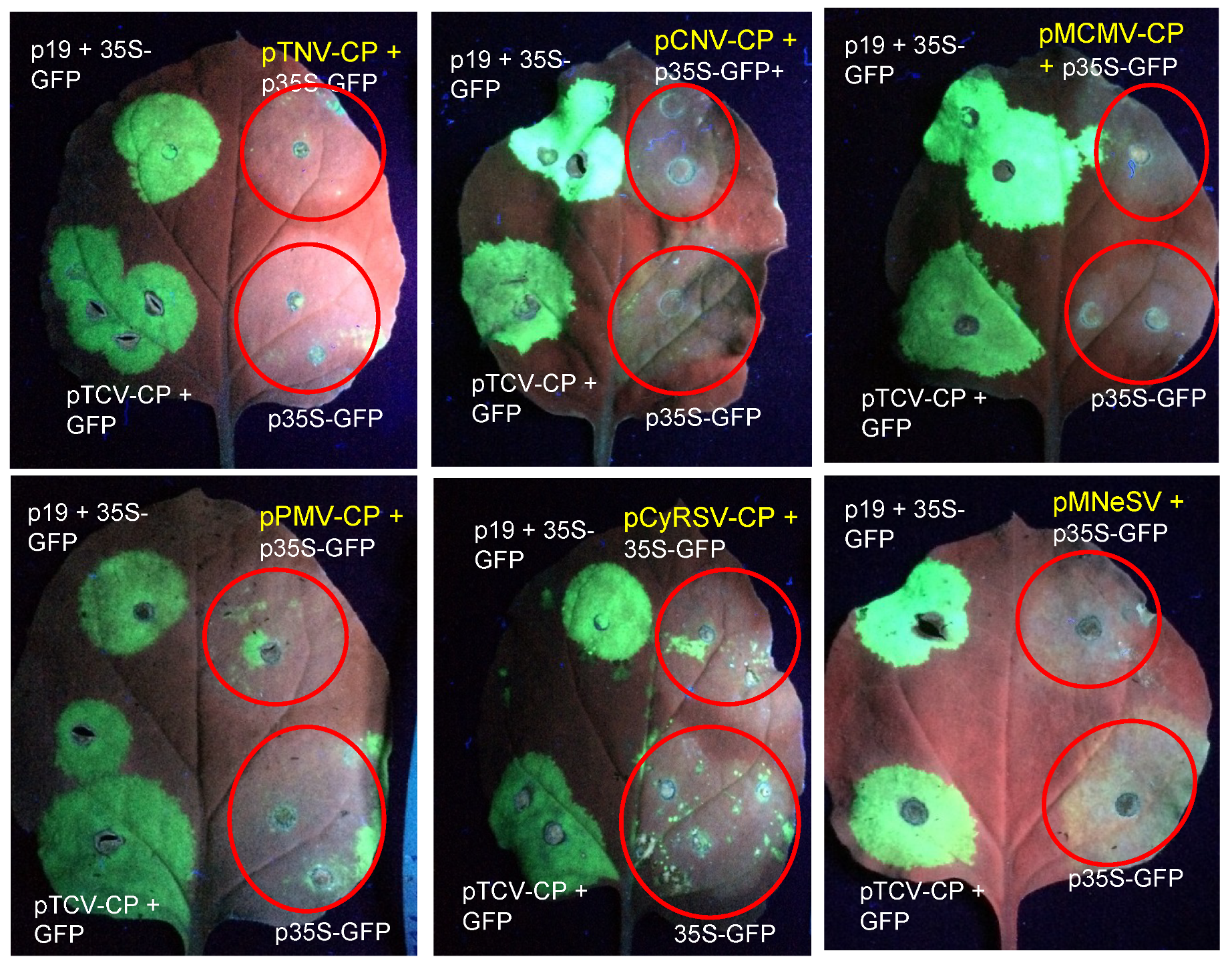
2.4. Agroinfiltration Assay for HR Elicitation and Silencing Suppression
2.5. ELISA Assay for TNV CP expression
3. Results
3.1. Survey of Nicotiana Species for Resistance to TNVDH Virion Inoculations
3.2. The TNVDH Coat Protein Triggers HR in Nicotiana Species Section Alatae.
3.3. Evaluation of the Coat Proteins of the Tombusviridae for Triggering HR in Members of Nicotiana Section Alatae
3.4. Evaluation of the CPs of Selected Tombusvirids for a Functional Silencing Suppressor in N. benthamiana
4. Discussion
4.1. Tracing the Lineage of HR Induction and Silencing Suppression Associated with the CPs of the Tombusviridae
4.2. Tracing the Lineage of Resistance Genes to the Tombusviridae in the Genus Nicotiana
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Qu, F.; Morris, T.J. Suppressors of RNA silencing encoded by plant viruses and their role in viral infections. FEBS Lett. 2005, 579, 5958–5964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholthof, H.B. The tombusvirus-encoded P19: From irrelevance to elegance. Nat. Rev. Microbiol. 2006, 4, 405–411. [Google Scholar] [CrossRef] [PubMed]
- deRonde, D.; Butterbach, P.; Kormelink, R. Dominant resistance against plant viruses. Front. Plant Sci. 2014, 5, 307. [Google Scholar] [CrossRef] [Green Version]
- Mandadi, K.K.; Scholthof, K.-B.G. Plant immune responses against viruses: How does a virus cause disease? Plant Cell 2013, 25, 1489–1505. [Google Scholar] [CrossRef] [PubMed]
- Zvereva, A.S.; Pooggin, M.M. Silencing and innate immunity in plant defense against viral and non-viral pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef] [PubMed]
- Leisner, S.M.; Schoelz, J.E. Joining the crowd: Integrating plant virus proteins into the larger world of pathogen effectors. Annu. Rev. Phytopathol. 2018, 56, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Schoelz, J.E. Viral determinants of resistance and susceptibility. In Natural Resistance Mechanisms of Plant to Viruses; Loebenstein, G., Carr, J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 13–43. [Google Scholar]
- Rochon, D.; Rubino, L.; Russo, M.; Martelli, G.P.; Lommel, S. Tombusviridae. In Virus Taxonomy-Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Academic Press: London, UK, 2012; pp. 1111–1138. [Google Scholar]
- Scheets, K.; White, K.A.; Rubino, L.; Martelli, G.P.; Rochon, D.A. ICTV taxonomic proposal 2015.007a-rP.A.v1.split_Carmovirus. Divide the Genus Carmovirus (family Tombusviridae) into Three New Genera: Alphacarmovirus, Betacarmovirus, and Gammacarmovirus. 2015. Available online: https://talk.ictvonline.org/ICTV/proposals/2015.007a-rP.A.v1.split_Carmovirus.pdf (accessed on 20 May 2019).
- Scheets, K.; Jordan, R.; White, A.; Hernandez, C. ICTV taxonomic proposal 2014.006b-fP.A..v3.Pelarspovirus. In the family Tombusviridae assign 5 previously unassigned species to a new genus, Pelarspovirus. 2014. Available online: https://talk.ictvonline.org/files/ictv_official_taxonomy_updates_since_the_8th_report/m/plant-official/5847 (accessed on 20 May 2019).
- Adams, M.J.; King, A.M.; Carstens, E.B. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2013). Arch. Virol. 2013, 158, 2023–2030. [Google Scholar] [CrossRef] [Green Version]
- Scheets, K.; Redinbaugh, M.G. Infectious cDNA transcripts of maize necrotic streak virus: Infectivity and translational characteristics. Virology 2006, 350, 171–183. [Google Scholar] [CrossRef]
- Oh, J.-W.; Kong, Q.; Song, C.; Carpenter, C.D.; Simon, A.E. Open reading frames of Turnip crinkle virus involved in satellite symptom expression and incompatibility with Arabidopsis thaliana ecotype Dijon. Mol. Plant-Microbe. Interact. 1995, 8, 979–987. [Google Scholar] [CrossRef]
- Zhao, Y.; DelGrosso, L.; Yigit, E.; Dempsey, D.A.; Klessig, D.F.; Wobbe, K.K. The amino terminus of the coat protein of Turnip crinkle virus is the AVR factor recognized by resistant Arabidopsis. Mol. Plant-Microbe Interact. 2000, 13, 1015–1018. [Google Scholar] [CrossRef]
- Qu, F.; Morris, T.J. Efficient infection of Nicotiana benthamiana by Tomato bushy stunt virus is facilitated by the coat protein and maintained by p19 through suppression of gene silencing. Mol. Plant-Microbe Interact. 2002, 15, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ren, T.; Morris, T.J. The coat protein gene of turnip crinkle virus suppresses posttranscriptional gene silencing at an early initiation step. J. Virol. 2003, 77, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.L.; Leh, V.; Lederer, C.; Maule, A.J. Turnip crinkle virus coat protein mediates suppression of RNA silencing in Nicotiana benthamiana. Virology 2003, 306, 33–41. [Google Scholar] [CrossRef]
- Choi, C.W.; Qu, F.; Ren, T.; Ye, X.; Morris, T.J. RNA silencing-suppressor function of Turnip crinkle virus coat protein cannot be attributed to its interaction with the Arabidopsis protein TIP. J. Gen. Virol. 2004, 85, 3415–3420. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Ye, X.; Willie, K.; Lin, J.; Zhang, X.; Redinbaugh, M.G.; Simon, A.E.; Morris, T.J.; Qu, F. The capsid protein of turnip crinkle virus overcomes two separate defense barriers to facilitate systemic movement of the virus in Arabidopsis. J. Virol. 2010, 84, 7793–7802. [Google Scholar] [CrossRef] [PubMed]
- Genovés, A.; Navarro, J.A.; and Pallás, V. Functional analysis of the five melon necrotic spot virus genome-encoded proteins. J. Gen. Virol. 2006, 87, 2371–2380. [Google Scholar] [CrossRef]
- Martinez-Turiño, S.; Hernandez, C. Inhibition of RNA silencing by the coat protein of pelargonium flower break virus: Distinctions from closely related suppressors. J. Gen. Virol. 2009, 90, 519–525. [Google Scholar] [CrossRef]
- Meng, C.; Chen, J.; Peng, J.; Wong, S.M. Host-induced avirulence of Hibiscus chlorotic ringspot virus mutant correlates with reduced gene-silencing suppression activity. J. Gen. Virol. 2006, 87, 451–459. [Google Scholar] [CrossRef]
- Pérez-Cañamás, M.; Hernández, C. Key importance of small RNA binding for the activity of a glycine/tryptophan (GW) motif-containing viral suppressor of RNA silencing. J. Biol. Chem. 2015, 290, 3106–3120. [Google Scholar] [CrossRef]
- Angel, C.A.; Schoelz, J.E. A survey of resistance to tomato bushy stunt virus in the genus Nicotiana reveals that the hypersensitive response is triggered by one of three different viral proteins. Mol. Plant-Microbe Interact. 2013, 26, 240–248. [Google Scholar] [CrossRef]
- Coutts, R.H.A.; Rigden, J.E.; Slabas, A.R.; Lomonosssoff, G.P.; Wise, P.J. The complete nucleotide sequence of tobacco necrosis virus strain D. J. Gen. Virol. 1991, 72, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Havelda, Z.; Dalmay, T.; Szutorisz, H.; and Burgyan, J. Complete nucleotide sequence of tobacco necrosis virus strain DH and genes requried for RNA replication and virus movement. J. Gen. Viol. 1997, 78, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Chkuaseli, T.; Newburn, L.R.; Bakhshinyan, D.; White, K.A. Protein expression strategies in tobacco necrosis virus-D. Virology 2015, 486, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Coutts, R.H. Investigations on the tobacco necrosis virus D p60 replicase protein. PLoS ONE 2013, 8, e80912. [Google Scholar] [CrossRef] [PubMed]
- Price, W.C. Host ranges of six plant viruses. Am. J. Bot. 1940, 27, 530–541. [Google Scholar] [CrossRef]
- Bascó, R.; Künstler, A.; Király, L. Tobacco necrosis virus replication and spread in Arabidopsis thaliana ecotype Columbia: A potential system for studying plant defense reactions to symptomless virus infections. Acta Physiol. Plant. 2016, 38, 139. [Google Scholar] [CrossRef]
- Lewis, R.S.; Nicholson, J.S. Aspects of the evolution of Nicotiana tabacum L. and the status of the Nicotiana Germplasm Collection. Genet. Resour. Crop Evol. 2007, 54, 727–740. [Google Scholar] [CrossRef]
- Grieco, F.; Burgyan, J.; Russo, M. The nucleotide sequence of cymbidium ringspot virus RNA. Nucleic Acids Res. 1989, 17, 6383. [Google Scholar] [CrossRef]
- Rochon, D.; Tremaine, J. Complete nucleotide sequence of the cucumber necrosis virus genome. Virology 1989, 169, 251–259. [Google Scholar] [CrossRef]
- Nutter, R.C.; Scheets, K.; Panganiban, L.C.; Lommel, S.A. The complete nucleotide sequence of the maize chlorotic mottle virus genome. Nucl. Acids Res. 1989, 17, 3163–3177. [Google Scholar] [CrossRef] [Green Version]
- Turina, M.; Maruoka, M.; Monis, J.; Jackson, A.O.; Scholthof, K.B. Nucleotide sequence and infectivity of a full-length cDNA clone of panicum mosaic virus. Virology 1998, 241, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Riviere, C.J.; Rochon, D.M. Nucleotide sequence and genomic organization of melon necrotic spot virus. J. Gen. Virol. 1990, 71, 1887–1896. [Google Scholar] [CrossRef] [PubMed]
- Lazo, G.R.; Stein, P.A.; Ludwig, R.A. A DNA transformation-competent Arabidopsis genomic library in Agrobacterium. BioTechnology 1991, 9, 963–967. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Kim, K.H.; Kendall, T.L.; Lommel, S.A. Synthesis of the putative red clover necrotic mosaic virus RNA polymerase by ribosomal frameshifting in vitro. Virology 1993, 193, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Hajdukiewicz, P.; Svab, Z.; and Maliga, P. The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 1994, 25, 989–994. [Google Scholar] [CrossRef]
- Carrington, J.C.; Heaton, L.A.; Zuidema, D.; Hillman, B.I.; Morris, T.J. The genome structure of turnip crinkle virus. Virology 1989, 170, 219–226. [Google Scholar] [CrossRef]
- Hearne, P.Q.; Knorr, D.A.; Hillman, B.I.; Morris, T.J. The complete genome structure and synthesis of infectious RNA from clones of Tomato bushy stunt virus. Virology 1990, 177, 141–151. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Angel, C.A.; Hsieh, Y.-C.; Schoelz, J.E. Comparative analysis of the capacity of tombusvirus P22 and P119 proteins to function as avirulence determinants in Nicotiana species. Mol. Plant-Microbe Interact. 2011, 24, 91–99. [Google Scholar] [CrossRef]
- Scheets, K. Analysis of gene functions in maize chlorotic mottle virus. Virus Res. 2016, 222, 71–79. [Google Scholar] [CrossRef]
- Palanichelvam, K.; Cole, A.B.; Shababi, M.; Schoelz, J.E. Agroinfiltration of cauliflower mosaic virus gene VI elicits hypersensitive response in Nicotiana species. In Mol. Plant-Microbe-Interact.; 2000; Volume 13, pp. 1275–1279. [Google Scholar]
- Abbink, T.E.M.; Tjernberg, P.A.; Bol, J.F.; Linthorst, H.J.M. Tobacco mosaic virus helicase domain induces necrosis in N-gene-carrying tobacco in the absence of virus replication. Mol. Plant-Microbe. Interact. 1998, 11, 1242–1246. [Google Scholar] [CrossRef]
- Erickson, F.; Holzberg, S.; Calderon-Urrea, A.; Handley, V.; Axtell, M.; Corr, C.; Baker, B. The helicase domain of the TMV replicase proteins induces the N-mediated defense response in tobacco. Plant J. 1999, 18, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liu, Y.; Yu, H.; Yuan, C.; Zeng, J.; Zhao, L.; Tong, Z.; Tao, X. Non-structural protein NSm of tomato spotted wilt virus is an avirulence factor recognized by resistance genes of tobacco and tomato via different elicitor active sites. Viruses 2018, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Daubert, S.D.; Schoelz, J.E.; Li, D.; Shepherd, R.J. Expression of disease symptoms in cauliflower mosaic virus genomic hybrids. J. Mol. Appl. Genet. 1984, 2, 537–547. [Google Scholar] [PubMed]
- Schoelz, J.E.; Shepherd, R.J.; Daubert, S.D. Region VI of cauliflower mosaic virus encodes a host range determinant. Mol. Cell. Biol. 1986, 6, 2632–2637. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, H.B.; Scholthof, K.-B.G.; Jackson, A.O. Identification of tomato bushy stunt virus host-specific symptom determinants by expression of individual genes from a potato virus X vector. Plant Cell 1995, 7, 1157–1172. [Google Scholar]
- Wang, K.; Empleo, R.; Nguyen, T.T.V.; Moffett, P.; Sacco, M.A. Elicitation of hypersensitive responses in Nicotiana glutinosa by the suppressor of RNA silencing protein P0 from poleroviruses. Mol. Plant Pathol. 2015, 16, 435–448. [Google Scholar] [CrossRef]
- Padgett, H.S.; Beachy, R.N. Analyis of a tobacco mosaic virus strain capable of overcoming N gene-mediated resistance. Plant Cell 1993, 5, 577–586. [Google Scholar]
- deRonde, D.; Butterbach, P.; Lohuis, D.; Hedil, M.; van Lent, J.W.M.; Kormelink, R. Tsw gene-based resistance is trigered by a functional RNA silencing suppressor portein of the tomato spotted wilt virus. Mol. Plant Pathol. 2013, 14, 405–415. [Google Scholar] [CrossRef]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–791. [Google Scholar] [CrossRef]
- Vleeshouwers, V.G.A.A.; Rietman, H.; Krenek, P.; Champouret, N.; Young, C.; Oh, S.-K.; Wang, M.; Bouwmeester, K.; Vosman, B.; Visser, R.G.F.; et al. Effector genomics accelerates discovry and funcational profiling of potato disease resistance and Phytophthora infestans avirulence genes. PLoS ONE 2008. [Google Scholar] [CrossRef] [PubMed]
- Morales, F.; Castano, M.; Calvert, L.; Arroyave, J.A. Furcraea necrotic streak virus: An apparent new member of the dianthovirus group. J. Phytopathol. 1992, 134, 247–254. [Google Scholar] [CrossRef]
- Powers, J.G.; Sit, T.L.; Heinsohn, C.; Gerorge, C.G.; Kim, K.-H.; Lommel, S.A. The red clover necrotic mosaic virus RNA-2 encoded movement protein is a second suppressor of RNA silencing. Virology 2008, 381, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Tsukuda, M.; Mizumoto, H.; Okamoto, K.; Kaido, M.; Mise, K.; Okuno, T. A plant RNA virus suppresses RNA silencing through viral RNA replication. EMBO J. 2005, 24, 3147–3157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miras, M.; Sepere, R.N.; Kraft, J.J.; Miller, W.A.; Aradna, M.A.; Truniger, V. Interfamilial recombination between viruses led to acquistion of a novel translation-enhancing RNA element that allows resistance breaking. New Phytol. 2014, 202, 233–246. [Google Scholar] [CrossRef]
- Ren, T.; Qu, F.; Morris, T.J. HRT gene function requires interaction between a NAC protein and viral capsid protein to confer resistance to turnip crinkle virus. Plant Cell 2000, 12, 1917–1925. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.S.; Masuta, C. Interaction between viral RNA silencing suppressors and host factors in plant immunity. Curr. Opin. Plant Biol. 2014, 20, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defense, counter-defense and counter-counter-defense. Nat. Rev. Microbiol. 2013, 11, 745–759. [Google Scholar] [CrossRef]
- Sit, S.L.; Lommel, S.A. Tombusviridae. In eLS; John Wiley and Sons, Ltd: Chichester, UK, 2015. [Google Scholar] [CrossRef]
- Culver, J.N. Tobacco mosaic virus assembly and disassembly: Determinants in pathogenicity and resistance. Annu. Rev. Phytopathol. 2002, 40, 287–308. [Google Scholar] [CrossRef]
- Toedt, J.M.; Braswell, E.H.; Schuster, T.M.; Yphantis, D.A.; Taraporewala, Z.F.; Culver, J.N. Biophysical characterization of a designed TMV coat protein mutant, R46G, that elicits a moderate hypersensitive response in Nicotiana sylvestris. Protein Sci. 1999, 8, 261–270. [Google Scholar] [CrossRef]
- Doroszewska, T.; Depta, A. Resistance of wild Nicotiana species to different PVY isolates. Phytopathologia 2011, 59, 9–24. [Google Scholar]
- Laskowska, D.; Doroszewska, T.; Depta, A.; Kursa, K.; Olszak-Przybyś, H.; Czubacka, A. A survey of Nicotiana germplasm for resistance to Tomato spotted wilt virus (TSWV). Euphytica 2013, 193, 207. [Google Scholar] [CrossRef]
- Yuan, X.; Yan, C.; Wu, Z.; Ren, F.; Zhang, H.; Baker, B.; Chen, J.; Kuang, H. Frequent gain and loss of resistance against tobacco mosaic virus in Nicotiana species. Mol. Plant 2015, 8, 1813–1815. [Google Scholar] [CrossRef] [PubMed]
- Balaji, B.; Cawly, J.; Angel, C.; Zhang, Z.; Palanichelvam, K.; Cole, A.; Schoelz, J. Silencing of the N family of resistance genes in Nicotiana edwardsonii compromises the hypersensitive response to tombusviruses. Mol. Plant-Microbe Interact. 2007, 20, 1262–1270. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nicotiana spp. | Section | TNV Virion Inoc.a | Agroinfiltration of pTNV-CP |
|---|---|---|---|
| N. langsdorffii | Alatae | HR b | N c |
| N. longiflora | Alatae | HR | N |
| N. bonariensis | Alatae | HR | N |
| N. alata | Alatae | HR | N |
| N. forgetiana | Alatae | HR | N |
| N. plumbaginifolia | Alatae | HR | no rxn d |
| N. quadrivalvis | Polydicliae | HR | no rxn |
| N. clevelandii | Polydicliae | HR | no rxn |
| N. edwardsonii d | Undulatae/Polydicliae | HR | no rxn |
| N. glutinosa | Undulatae | HR | no rxn |
| N. arentsii | Undulatae | HR | no rxn |
| N. undulata | Undulatae | HR | no rxn |
| N. tabacum | Nicotiana | HR | no rxn |
| N. sylvestris | Sylvestres | HR | no rxn |
| N. otophora | Tomentosae | CLL e | no rxn |
| N. tomentosiformis | Tomentosae | HR | no rxn |
| N. repanda | Repandae | HR | no rxn |
| N. glauca | Noctiflorae | HR | no rxn |
| N. rustica | Rusticae | HR | no rxn |
| N. benthamiana | Suaveolentes | Susc f | no rxn |
| Virus CP | N. langsdorffii | N. longiflora | N. alata tw7 | N. forgetiana tw50 | N. plumbaginifolia tw106 | Silencing Suppressor Activity a |
|---|---|---|---|---|---|---|
| tombusvirus | ||||||
| pTBSV | N b | N | N | N | no rxn c | No |
| pCyRSV | N | N | N | N | no rxn | No |
| pCNV | N | N | N | N | no rxn | No |
| gammacarmovirus | ||||||
| pMNSV | N | N | N | N | no rxn | Yes |
| dianthovirus | ||||||
| pRCNMV | N | N | N | N | no rxn | No |
| betacarmovirus | ||||||
| pTCV | no rxn | no rxn | no rxn | no rxn | no rxn | Yes |
| pelarspovirus | ||||||
| pPLPV | no rxn | no rxn | no rxn | no rxn | no rxn | Yes |
| alphacarmovirus | ||||||
| pPFBV | no rxn | no rxn | no rxn | no rxn | no rxn | Yes |
| zeavirus | ||||||
| pMNeSV | N | N | N | N | no rxn | No |
| alphanecrovirus | ||||||
| pTNV | N | N | N | N | no rxn | No |
| panicovirus | ||||||
| pPMV | no rxn | no rxn | no rxn | no rxn | no rxn | No |
| machlomovirus | ||||||
| pMCMV | no rxn | no rxn | no rxn | no rxn | no rxn | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhab, M.; Angel, C.; Rodriguez, A.; Fereidouni, M.; Király, L.; Scheets, K.; Schoelz, J.E. Tracing the Lineage of Two Traits Associated with the Coat Protein of the Tombusviridae: Silencing Suppression and HR Elicitation in Nicotiana Species. Viruses 2019, 11, 588. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070588
Adhab M, Angel C, Rodriguez A, Fereidouni M, Király L, Scheets K, Schoelz JE. Tracing the Lineage of Two Traits Associated with the Coat Protein of the Tombusviridae: Silencing Suppression and HR Elicitation in Nicotiana Species. Viruses. 2019; 11(7):588. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070588
Chicago/Turabian StyleAdhab, Mustafa, Carlos Angel, Andres Rodriguez, Mohammad Fereidouni, Lóránt Király, Kay Scheets, and James E. Schoelz. 2019. "Tracing the Lineage of Two Traits Associated with the Coat Protein of the Tombusviridae: Silencing Suppression and HR Elicitation in Nicotiana Species" Viruses 11, no. 7: 588. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070588