A Nymphalid-Infecting Group I Alphabaculovirus Isolated from the Major Passion Fruit Caterpillar Pest Dione juno juno (Lepidoptera: Nymphalidae)

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Purification and Insects
2.2. Transmission Electron Microscopy
2.3. Genome Sequencing, Assembly, and Annotation
2.4. Phylogenetic Analyses
3. Results
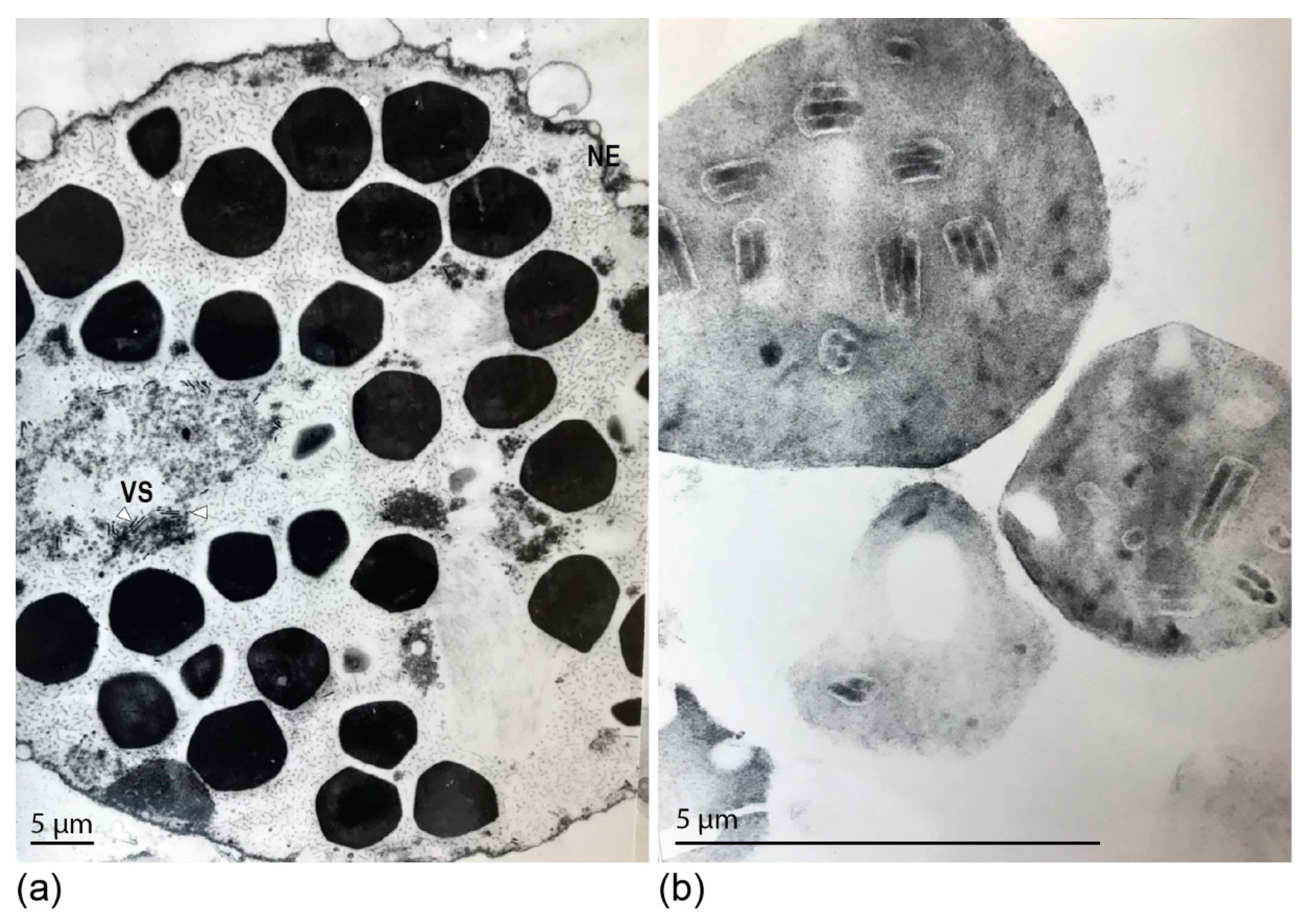
3.1. Ultrastructure of DijuNPV Occlusion Bodies (OBs)
3.2. General Features of the DijuNPV Genome Sequence
3.3. Sequence Variation in the DijuNPV Virus Population
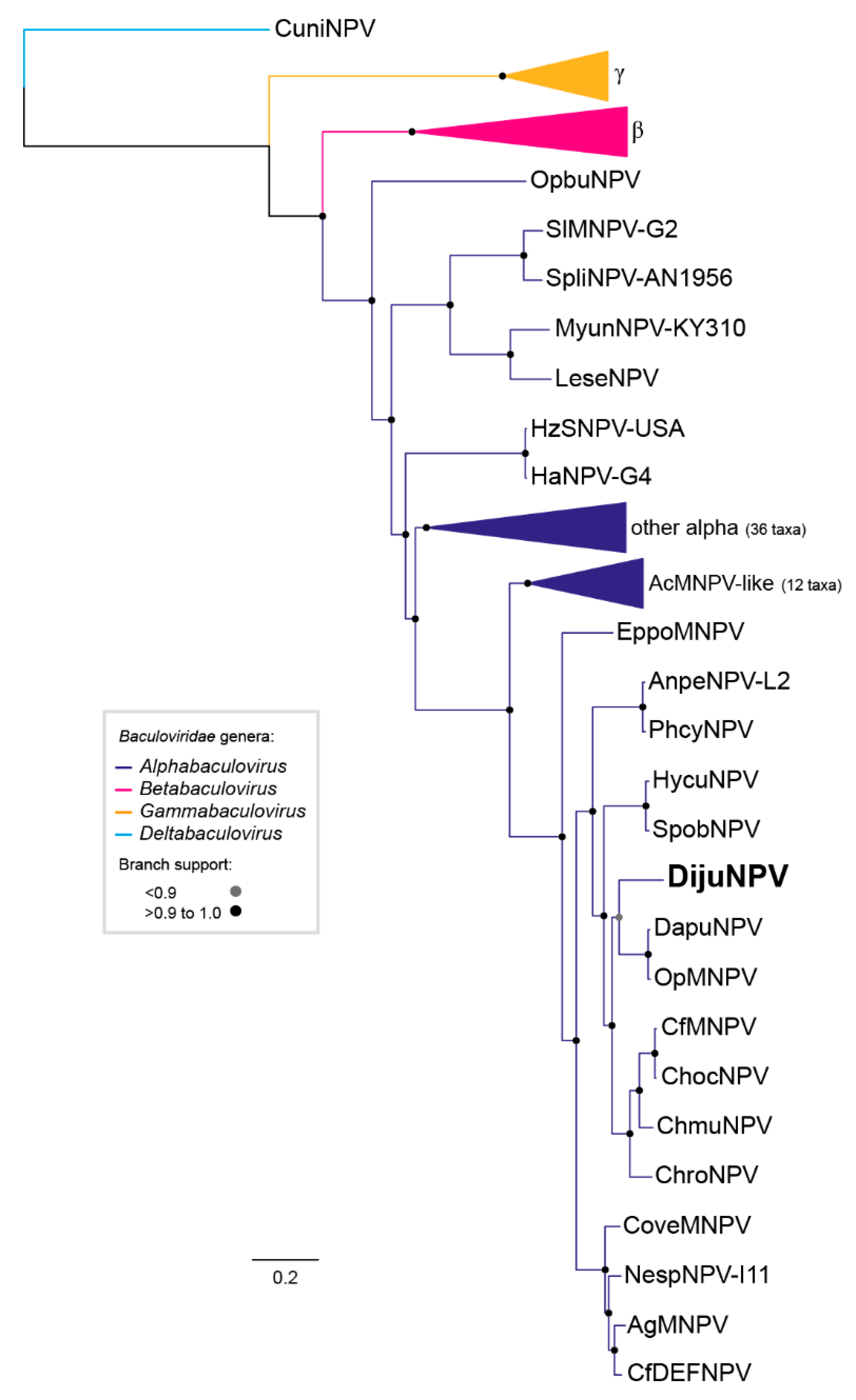
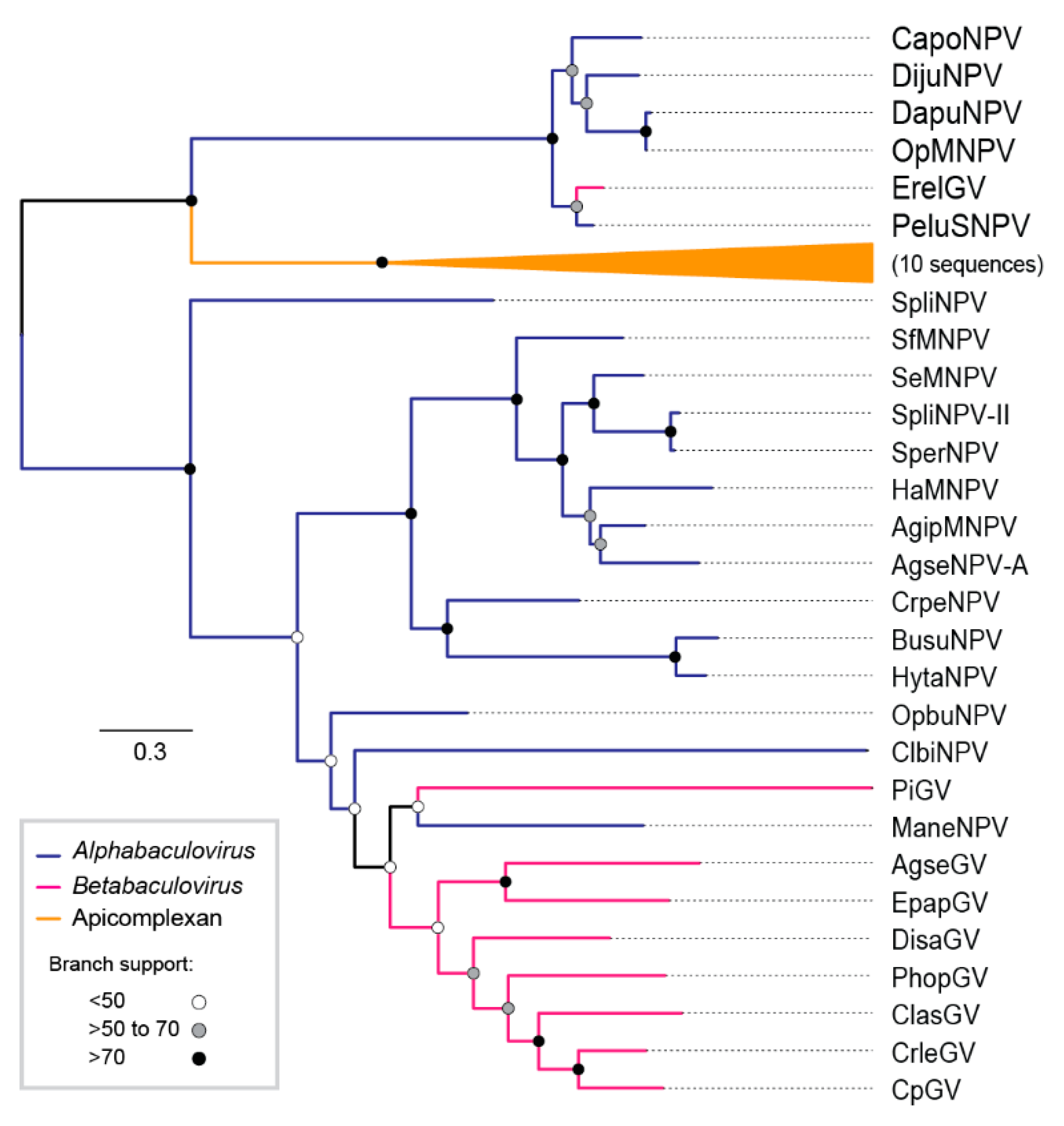
3.4. Phylogeny of DijuNPV
3.5. DijuNPV May be a Representative of a Novel Species Inside Genus Alphabaculovirus
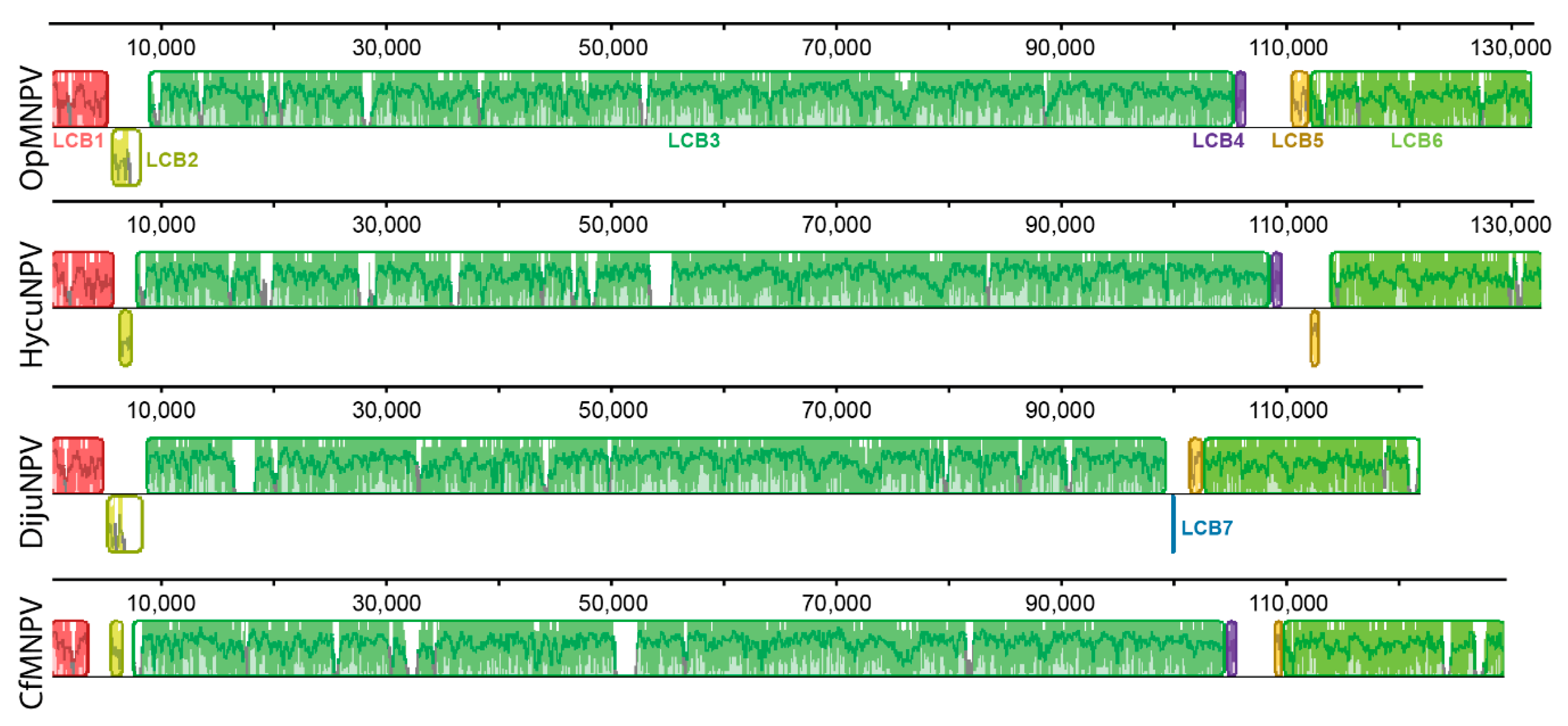
3.6. Genomic Analysis
3.7. DijuNPV ORF Content
3.8. The tmk Homolog Locus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 15, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L.; Peng, H.Y. Recent advances in biological control of pest insects by using viruses in China. Virol. Sin. 2007, 22, 158–162. [Google Scholar] [CrossRef]
- Szewczyk, B.; Hoyos-Carvajal, L.; Paluszek, M.; Skrzecz, I.; De Souza, M.L. Baculoviruses—re-emergingbiopesticides. Biotechnol. Adv. 2006, 24, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Clem, R.J.; Passarelli, A.L. Baculoviruses: Sophisticated pathogens of insects. PLoS Pathog. 2013, 9, 1003729. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK114593/pdf/Bookshelf_NBK114593.pdf (accessed on 12 December 2013).
- Volkman, L.E. Nucleopolyhedrovirus interactions with their insect hosts. Adv. Virus. Res. 1997, 48, 313–348. [Google Scholar]
- Katsuma, S.; Noguchi, Y.; Zhou, C.L.; Kobayashi, M.; Maeda, S. Characterization of the 25K FP gene of the baculovirus Bombyx mori nucleopolyhedrovirus: Implications for post-mortem host degradation. J. Gen. Virol. 1999, 80, 783–791. [Google Scholar] [CrossRef]
- Martignoni, M.E.; Iwai, P.J. A catalogue of viral diseases of insects, mites and ticks. In Microbial Control of Pests and Plant Diseases 1970–1980; Burges, H.D., Ed.; Academic Press: London, UK, 1981; pp. 897–911. [Google Scholar]
- Thiem, S.M. Insect virus diversity. Trends Microbiol. 1999, 7, 463. [Google Scholar] [CrossRef]
- Moscardi, F. Assessment of the application of baculoviruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef]
- Haase, S.; Sciocco-Cap, A.; Romanowski, V. Baculovirus insecticides in Latin America: Historical overview, current status and future perspectives. Viruses 2015, 7, 2230–2267. [Google Scholar] [CrossRef]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; van Oers, M.M.; Donlye, B.C.; Erlandsona, M.A.; Hegedus, D.D.; Theilmann, D.A. Autographa californica multiple nucleopolyhedrovirus AC83 is a per os infectivity factor (PIF) protein required for occlusion-derived virus (ODV) and budded virus nucleocapsid assembly as well as assembly of the PIF complex in ODV envelopes. J. Virol. 2017, 91, e02115-16. [Google Scholar] [CrossRef]
- Ribeiro, B.M.; Zanotto, P.M.A.; McDowell, S.; De Souza, M.L.; Kitajima, E.W. Characterization of a baculovirus infecting the passion fruit caterpillar Dione juno juno. Biocell 1997, 21, 71–82. [Google Scholar]
- Rodríguez, V.A.; Belaich, M.N.; Gómez, D.L.M.; Sciocco-Cap, A.; Ghiringhelli, P.D. Identification of nucleopolyhedrovirus that infect Nymphalid butterflies Agraulis vanillae and Dione juno. J. Invertebr. Pathol. 2011, 106, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Gravena, S. Perspectivas do manejo integrado de pragas. In Cultura do maracujazeiro; Legis Summa: Ribeirão Preto, Brasil, 1987; pp. 134–145. [Google Scholar]
- EMBRAPA. Destinos das exportações brasileiras de maracujá em 2017. Available online: http://www.cnpmf.embrapa.br/Base_de_Dados/index_pdf/dados/brasil/maracuja/b61_maracuja.pdf (accessed on 8 February 2019).
- Gentz, M.C.; Murdoch, G.; King, G.F. Tandem use of selective insecticides and natural enemies for effective, reduced-risk pest management. Biol. Control 2010, 52, 208–215. [Google Scholar] [CrossRef]
- Sarwar, M. Microbial insecticides-an ecofriendly effective line of attack for insect pests management. IJEART 2015, 2, 4–9. [Google Scholar]
- Villani, H.C.; Campos, A.R.; Gravena, S. Effectiveness of Bacillus thuringiensis Berliner and fenitrothion+ fenvalerate for the control of the passion-fruit larva Dione juno juno (Cramer, 1779) (Lepidoptera, Heliconidae). An. Soc. Entomol. Bras. 1980, 9, 255–260. [Google Scholar]
- O’Reilly, D.R.; Miller, L.K.; Luckow, V.A. Baculovirus Expression Vectors: A Laboratory Manual; W.H. Freeman & Company: New York, NY, USA, 1992. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Chen, Y.R.; Zhong, S.; Fei, Z.; Hashimoto, Y.; Xiang, J.Z.; Zhang, S.; Blissard, G.W. The transcriptome of the baculovirus Autographa californica multiple nucleopolyhedrovirus in Trichoplusia ni cells. J. Virol. 2013, 87, 6391–6405. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Jehle, J.A.; Lange, M.; Wang, H.; Hu, Z.; Wang, Y.; Hauschild, R. Molecular identification and phylogenetic analysis of baculoviruses from Lepidoptera. Virology 2006, 346, 180–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garavaglia, M.J.; Miele, S.A.B.; Iserte, J.A.; Belaich, M.N.; Ghiringhelli, P.D. Ac53, ac78, ac101 and ac103 are newly discovered core genes in the family Baculoviridae. J. Virol. 2012, 86, 12069–12079. [Google Scholar] [CrossRef] [PubMed]
- Regier, J.C.; Zwick, A.; Cummings, M.P.; Kawahara, A.Y.; Cho, S.; Weller, S.; Roe, A.; Baixeras, J.; Brown, J.W.; Parr, C.; et al. Toward reconstructing the evolution of advanced moths and butterflies (Lepidoptera: Ditrysia): An initial molecular study. BMC Evol. Biol. 2009, 9, 280. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.R.; Oliveira, L.B.; Peterson, L.; Sosa-Gómez, D.R.; Ribeiro, B.M.; Ardisson-Araújo, D.M.P. The complete genome sequence of the first hesperiid-infecting alphabaculovirus isolated from the leguminous pest Urbanus proteus (Lepidoptera: Hesperiidae). Virus. Res. 2018, 249, 76–84. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, Z.; Zhang, L.; Hou, D.; Wang, M.; Arif, B.; Kou, Z.; Wang, H.; Deng, F.; Hu, Z. Genome sequencing and analysis of Catopsilia pomona nucleopolyhedrovirus: A distinct species in group I alphabaculovirus. PLoS ONE 2016, 11, e0155134. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Cheng, R.L.; Wang, X.F.; Zhang, C.X. The genome of Pieris rapae granulovirus. J. Virol. 2012, 86, 9544. [Google Scholar] [CrossRef]
- Harrison, R.L.; Rowley, D.L.; Mowery, J.D.; Bauchan, G.R.; Burand, J.P. The Operophtera brumata nucleopolyhedrovirus (OpbuNPV) represents an early, divergent lineage within genus Alphabaculovirus. Viruses 2017, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Ardisson-Araújo, D.M.P.; Melo, F.L.; Clem, R.J.; Wolff, J.L.; Ribeiro, B.M. A betabaculovirus-encoded gp64 homolog codes for a functional envelope fusion protein. J. Virol. 2016, 90, 1668–1672. [Google Scholar] [CrossRef] [PubMed]
- Minet, J. Tentative reconstruction of the ditrysian phylogeny (Lepidoptera: Glossata). Insect. Syst. Evol. 1991, 22, 69–95. [Google Scholar] [CrossRef]
- Ikeda, M.; Yanagimoto, K.; Kobayashi, M. Identification and functional analysis of Hyphantria cunea nucleopolyhedrovirus iap genes. Virology 2004, 321, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Chateigner, A.; Bézier, A.; Labrousse, C.; Jiolle, D.; Barbe, V.; Herniou, E.A. Ultra deep sequencing of a baculovirus population reveals widespread genomic variations. Viruses 2015, 7, 3625–3646. [Google Scholar] [CrossRef]
- Ardisson-Araújo, D.M.P.; Lima, R.N.; Melo, F.L.; Clem, R.J.; Huang, N.; Báo, S.N.; Sosa-Gómez, D.R.; Ribeiro, B.M. Genome sequence of Perigonia lusca single nucleopolyhedrovirus: Insights into the evolution of a nucleotide metabolism enzyme in the family Baculoviridae. Sci. Rep. 2016, 24612. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribeiro, B.M.; dos Santos, E.R.; Trentin, L.B.; da Silva, L.A.; de Melo, F.L.; Kitajima, E.W.; Ardisson-Araújo, D.M.P. A Nymphalid-Infecting Group I Alphabaculovirus Isolated from the Major Passion Fruit Caterpillar Pest Dione juno juno (Lepidoptera: Nymphalidae). Viruses 2019, 11, 602. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070602
Ribeiro BM, dos Santos ER, Trentin LB, da Silva LA, de Melo FL, Kitajima EW, Ardisson-Araújo DMP. A Nymphalid-Infecting Group I Alphabaculovirus Isolated from the Major Passion Fruit Caterpillar Pest Dione juno juno (Lepidoptera: Nymphalidae). Viruses. 2019; 11(7):602. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070602
Chicago/Turabian StyleRibeiro, Bergmann Morais, Ethiane Rozo dos Santos, Luana Beló Trentin, Leonardo Assis da Silva, Fernando Lucas de Melo, Elliot Watanabe Kitajima, and Daniel M. P. Ardisson-Araújo. 2019. "A Nymphalid-Infecting Group I Alphabaculovirus Isolated from the Major Passion Fruit Caterpillar Pest Dione juno juno (Lepidoptera: Nymphalidae)" Viruses 11, no. 7: 602. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070602