Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Protein Purification
2.3. Lytic Activity of Ts2631 Endolysin
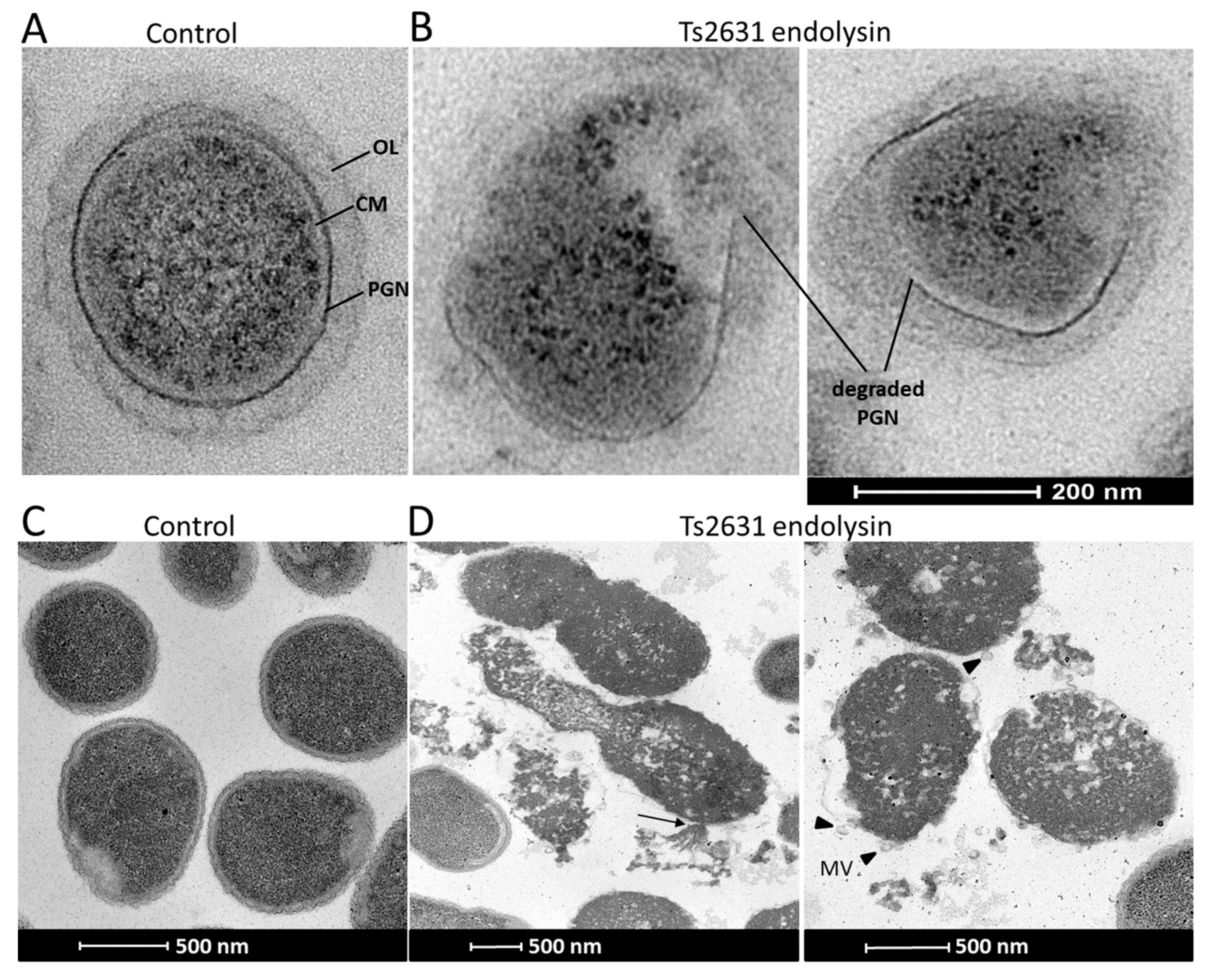
2.4. Transmission Electron Microscopy
2.5. Antibacterial Assays
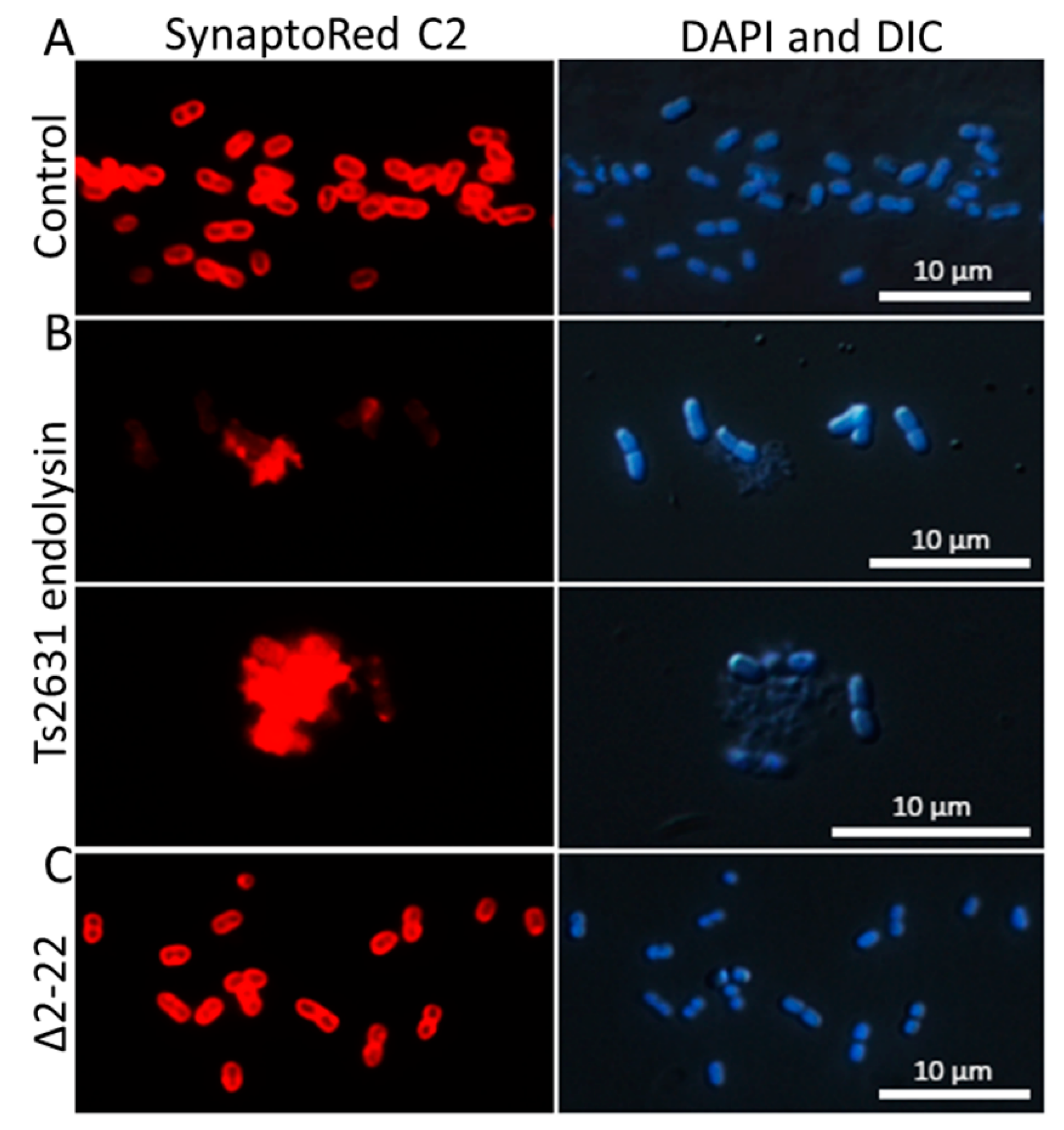
2.6. Fluorescence Microscopy
3. Results
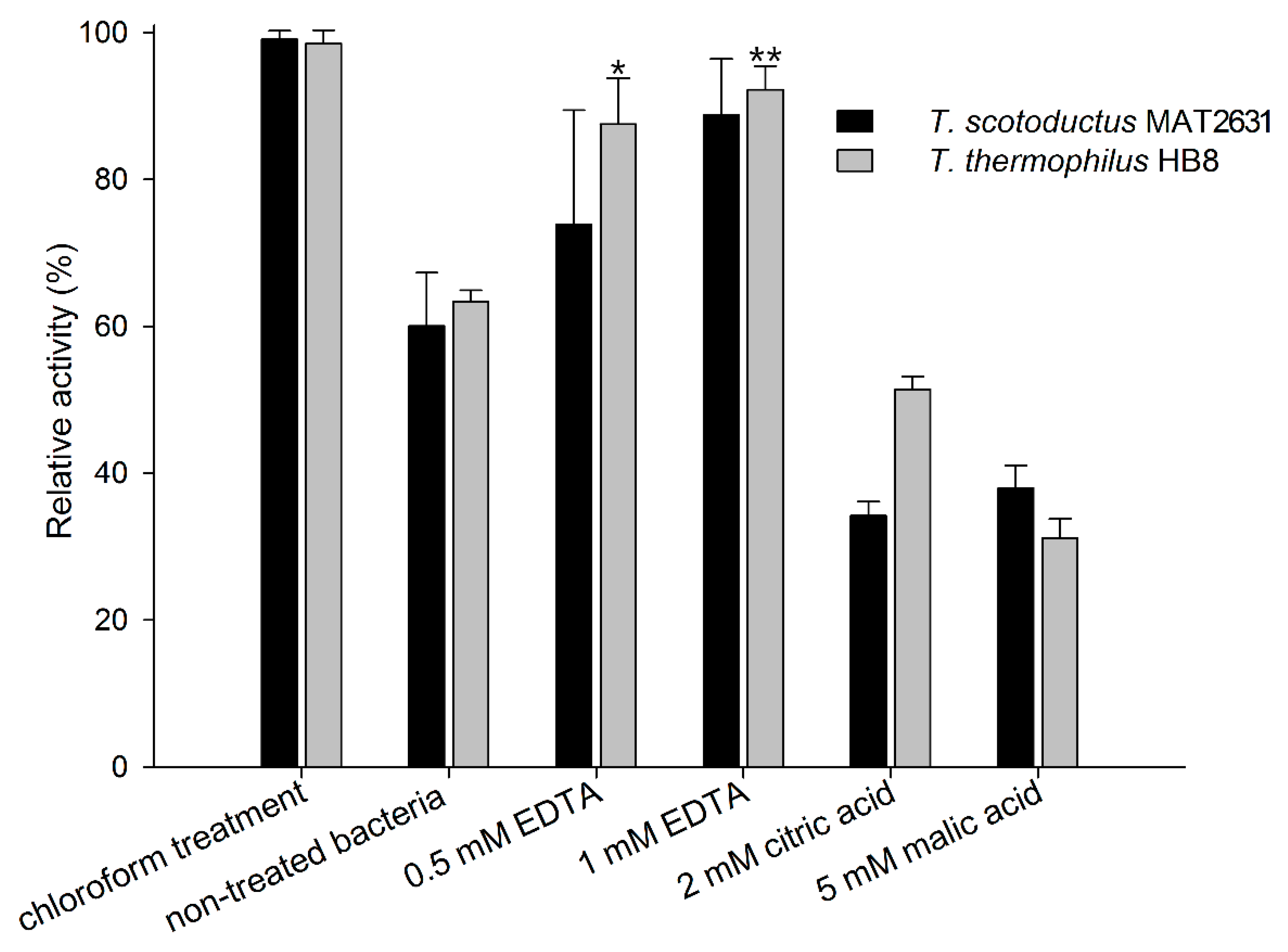
3.1. Activity of Ts2631 Endolysin against Viable Bacteria from the Thermus Genus
3.2. Bactericidal Activity of Ts2631 Endolysin against Gram-Negative MDR Bacterial Pathogens
3.3. Mode of Action of Ts2631 Endolysin on ESKAPE Pathogens
3.4. Intrinsic Membrane Passaging Capability of Ts2631 Endolysin
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed]
- ECDC/EMEA. ECDC/EMEA Joint Technical Report: The Bacterial Challenge: Time to React; European Centre for Disease Prevention and Control: Solna Municipality, Sweden, 2009. [Google Scholar]
- Cassini, A.; Plachouras, D.; Eckmanns, T.; Abu Sin, M.; Blank, H.P.; Ducomble, T.; Haller, S.; Harder, T.; Klingeberg, A.; Sixtensson, M.; et al. Burden of Six Healthcare-Associated Infections on European Population Health: Estimating Incidence-Based Disability-Adjusted Life Years through a Population Prevalence-Based Modelling Study. PLoS Med. 2016, 13, e1002150. [Google Scholar] [CrossRef] [PubMed]
- Turecka, K.; Waleron, K. Inhibitors of bacterial transcription are compounds for potent antimicrobial drugs. Curr. Pharm. Biotechnol. 2013, 14, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.; Harper, D.; et al. Alternatives to antibiotics-a pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef]
- Love, M.J.; Bhandari, D.; Dobson, R.C.J.; Billington, C. Potential for Bacteriophage Endolysins to Supplement or Replace Antibiotics in Food Production and Clinical Care. Antibiotics 2018, 7, 17. [Google Scholar] [CrossRef]
- Nakonieczna, J.; Wozniak, A.; Pieranski, M.; Rapacka-Zdonczyk, A.; Ogonowska, P.; Grinholc, M. Photoinactivation of ESKAPE pathogens: Overview of novel therapeutic strategy. Future Med. Chem. 2019, 11, 443–461. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Michalopoulos, A.; Falagas, M.E. Treatment of Acinetobacter infections. Expert Opin. Pharm. 2010, 11, 779–788. [Google Scholar] [CrossRef]
- WHO. Guidelines for the Prevention and Control of Carbapenem-Resistant Enterobacteriaceae, Acinetobacter baumannii and Pseudomonas aeruginosa in Health Care Facilities; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to manage Pseudomonas aeruginosa infections. Drugs Context 2018, 7, 1–18. [Google Scholar] [CrossRef]
- Hsu, L.Y.; Apisarnthanarak, A.; Khan, E.; Suwantarat, N.; Ghafur, A.; Tambyah, P.A. Carbapenem-Resistant Acinetobacter baumannii and Enterobacteriaceae in South and Southeast Asia. Clin. Microbiol. Rev. 2017, 30, 1–22. [Google Scholar] [CrossRef]
- Da Silva, K.E.; Maciel, W.G.; Croda, J.; Cayô, R.; Ramos, A.C.; de Sales, R.O.; Kurihara, M.N.L.; Vasconcelos, N.G.; Gales, A.C.; Simionatto, S. A high mortality rate associated with multidrug-resistant Acinetobacter baumannii ST79 and ST25 carrying OXA-23 in a Brazilian intensive care unit. PLoS ONE 2018, 13, e0209367. [Google Scholar] [CrossRef] [PubMed]
- Tomczyk, S.; Zanichelli, V.; Grayson, M.L.; Twyman, A.; Abbas, M.; Pires, D.; Allegranzi, B.; Harbarth, S. Control of Carbapenem-resistant Enterobacteriaceae, Acinetobacter baumannii, and Pseudomonas aeruginosa in Healthcare Facilities: A Systematic Review and Reanalysis of Quasi-experimental Studies. Clin. Infect. Dis. 2019, 68, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y. Phage Lytic Enzymes. Viruses 2019, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; São-José, C. Enzymes and Mechanisms Employed by Tailed Bacteriophages to Breach the Bacterial Cell Barriers. Viruses 2018, 10, 396. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, W.; Blanot, D.; de Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borysowski, J.; Weber-Dabrowska, B.; Górski, A. Bacteriophage endolysins as a novel class of antibacterial agents. Exp. Biol. Med. 2006, 231, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotka, M.; Kaczorowska, A.K.; Stefanska, A.; Morzywolek, A.; Fridjonsson, O.H.; Dunin-Horkawicz, S.; Kozlowski, L.; Hreggvidsson, G.O.; Kristjansson, J.K.; Dabrowski, S.; et al. Novel highly thermostable endolysin from Thermus scotoductus MAT2119 bacteriophage Ph2119 with amino acid sequence similarity to eukaryotic peptidoglycan recognition proteins. Appl. Environ. Microbiol. 2014, 80, 886–895. [Google Scholar] [CrossRef]
- Plotka, M.; Kaczorowska, A.K.; Morzywolek, A.; Makowska, J.; Kozlowski, L.P.; Thorisdottir, A.; Skírnisdottir, S.; Hjörleifsdottir, S.; Fridjonsson, O.H.; Hreggvidsson, G.O.; et al. Biochemical Characterization and Validation of a Catalytic Site of a Highly Thermostable Ts2631 Endolysin from the Thermus scotoductus Phage vB_Tsc2631. PLoS ONE 2015, 10, e0137374. [Google Scholar] [CrossRef] [PubMed]
- Plotka, M.; Sancho-Vaello, E.; Dorawa, S.; Kaczorowska, A.K.; Kozlowski, L.P.; Kaczorowski, T.; Zeth, K. Structure and function of the Ts2631 endolysin of Thermus scotoductus phage vB_Tsc2631 with unique N-terminal extension used for peptidoglycan binding. Sci. Rep. 2019, 9, 1261. [Google Scholar] [CrossRef]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Ruiz, I.; Coutinho, F.H.; Rodriguez-Valera, F. Thousands of Novel Endolysins Discovered in Uncultured Phage Genomes. Front. Microbiol. 2018, 9, 1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigoldi, F.; Donini, S.; Giacomina, F.; Sorana, F.; Redaelli, A.; Bandiera, T.; Parisini, E.; Gautieri, A. Thermal stabilization of the deglycating enzyme Amadoriase I by rational design. Sci. Rep. 2018, 8, 3042. [Google Scholar] [CrossRef] [PubMed]
- Vieille, C.; Zeikus, G.J. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability. Microbiol. Mol. Biol. Rev. 2001, 65, 1–43. [Google Scholar] [CrossRef]
- Pask-Hughes, R.; Williams, R.A. Extremely thermophilic gram-negative bacteria from hot tap water. J. Gen. Microbiol. 1975, 88, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowski, T.; Szybalski, W. Co-operativity of hexamer ligation. Gene 1996, 179, 189–193. [Google Scholar] [CrossRef]
- Stefanska, A.; Kaczorowska, A.K.; Plotka, M.; Fridjonsson, O.H.; Hreggvidsson, G.O.; Hjorleifsdottir, S.; Kristjansson, J.K.; Dabrowski, S.; Kaczorowski, T. Discovery and characterization of RecA protein of thermophilic bacterium Thermus thermophilus MAT72 phage Tt72 that increases specificity of a PCR-based DNA amplification. J. Biotechnol. 2014, 182, 1–10. [Google Scholar] [CrossRef]
- Stefanska, A.; Gaffke, L.; Kaczorowska, A.K.; Plotka, M.; Dabrowski, S.; Kaczorowski, T. Highly thermostable RadA protein from the archaeon Pyrococcus woesei enhances specificity of simplex and multiplex PCR assays. J. Appl. Genet. 2016, 57, 239–249. [Google Scholar] [CrossRef]
- Plotka, M.; Wozniak, M.; Kaczorowski, T. Quantification of Plasmid Copy Number with Single Colour Droplet Digital PCR. PLoS ONE 2017, 12, e0169846. [Google Scholar] [CrossRef]
- Filatova, L.Y.; Becker, S.C.; Donovan, D.M.; Gladilin, A.K.; Klyachko, N.L. LysK, the enzyme lysing Staphylococcus aureus cells: Specific kinetic features and approaches towards stabilization. Biochimie 2010, 92, 507–513. [Google Scholar] [CrossRef]
- Haddad Kashani, H.; Schmelcher, M.; Sabzalipoor, H.; Seyed Hosseini, E.; Moniri, R. Recombinant Endolysins as Potential Therapeutics against Antibiotic-Resistant Staphylococcus aureus: Current Status of Research and Novel Delivery Strategies. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Maciąg-Dorszyńska, M.; Ignatowska, M.; Jannière, L.; Węgrzyn, G.; Szalewska-Pałasz, A. Mutations in central carbon metabolism genes suppress defects in nucleoid position and cell division of replication mutants in Escherichia coli. Gene 2012, 503, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Thiagarajan, V.; Walmagh, M.; Sillankorva, S.; Lavigne, R.; Neves-Petersen, M.T.; Kluskens, L.D.; Azeredo, J. A thermostable Salmonella phage endolysin, Lys68, with broad bactericidal properties against gram-negative pathogens in presence of weak acids. PLoS ONE 2014, 9, e108376. [Google Scholar] [CrossRef] [PubMed]
- Quintela, J.C.; Pittenauer, E.; Allmaier, G.; Arán, V.; de Pedro, M.A. Structure of peptidoglycan from Thermus thermophilus HB8. J. Bacteriol. 1995, 177, 4947–4962. [Google Scholar] [CrossRef] [PubMed]
- Orito, Y.; Morita, M.; Hori, K.; Unno, H.; Tanji, Y. Bacillus amyloliquefaciens phage endolysin can enhance permeability of Pseudomonas aeruginosa outer membrane and induce cell lysis. Appl. Microbiol. Biotechnol. 2004, 65, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.A.; Shin, H.; Heu, S.; Ryu, S. Exogenous lytic activity of SPN9CC endolysin against gram-negative bacteria. J. Microbiol. Biotechnol. 2014, 24, 803–811. [Google Scholar] [CrossRef]
- Torrent, M.; Di Tommaso, P.; Pulido, D.; Nogués, M.V.; Notredame, C.; Boix, E.; Andreu, D. AMPA: An automated web server for prediction of protein antimicrobial regions. Bioinformatics 2012, 28, 130–131. [Google Scholar] [CrossRef]
- Nelson, D.; Loomis, L.; Fischetti, V.A. Prevention and elimination of upper respiratory colonization of mice by group A streptococci by using a bacteriophage lytic enzyme. Proc. Natl. Acad. Sci. USA 2001, 98, 4107–4112. [Google Scholar] [CrossRef] [Green Version]
- Fischetti, V.A. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The Preclinical and Clinical Progress of Bacteriophages and Their Lytic Enzymes: The Parts are Easier than the Whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, B.; Olszak, T.; Drulis-Kawa, Z. Applications of bacteriophages versus phage enzymes to combat and cure bacterial infections: An ambitious and also a realistic application? Appl. Microbiol. Biotechnol. 2018, 102, 2563–2581. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhang, X. Characterization of a lysin from deep-sea thermophilic bacteriophage GVE2. Appl. Microbiol. Biotechnol. 2008, 78, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Swift, S.M.; Seal, B.S.; Garrish, J.K.; Oakley, B.B.; Hiett, K.; Yeh, H.Y.; Woolsey, R.; Schegg, K.M.; Line, J.E.; Donovan, D.M. A Thermophilic Phage Endolysin Fusion to a Clostridium perfringens-Specific Cell Wall Binding Domain Creates an Anti-Clostridium Antimicrobial with Improved Thermostability. Viruses 2015, 7, 3019–3034. [Google Scholar] [CrossRef] [PubMed]
- Briers, Y.; Walmagh, M.; Van Puyenbroeck, V.; Cornelissen, A.; Cenens, W.; Aertsen, A.; Oliveira, H.; Azeredo, J.; Verween, G.; Pirnay, J.P.; et al. Engineered endolysin-based “Artilysins” to combat multidrug-resistant gram-negative pathogens. MBio 2014, 5, e01379–e01414. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Guo, Z.; Gao, C.; Zhu, R.; Wang, S.; Yu, L.; Qin, W.; Xia, X.; Gu, J.; Yan, G.; et al. Enhancement of the direct antimicrobial activity of Lysep3 against Escherichia coli by inserting cationic peptides into its C terminus. Antonie Van Leeuwenhoek 2017, 110, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Walmagh, M.; Briers, Y.; dos Santos, S.B.; Azeredo, J.; Lavigne, R. Characterization of modular bacteriophage endolysins from Myoviridae phages OBP, 201φ2-1 and PVP-SE1. PLoS ONE 2012, 7, e36991. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Tanji, Y.; Orito, Y.; Mizoguchi, K.; Soejima, A.; Unno, H. Functional analysis of antibacterial activity of Bacillus amyloliquefaciens phage endolysin against Gram-negative bacteria. FEBS. Lett. 2001, 500, 56–59. [Google Scholar] [CrossRef]
- Lai, M.J.; Lin, N.T.; Hu, A.; Soo, P.C.; Chen, L.K.; Chen, L.H.; Chang, K.C. Antibacterial activity of Acinetobacter baumannii phage ϕAB2 endolysin (LysAB2) against both gram-positive and gram-negative bacteria. Appl. Microbiol. Biotechnol. 2011, 90, 529–539. [Google Scholar] [CrossRef]
- Lai, M.J.; Soo, P.C.; Lin, N.T.; Hu, A.; Chen, Y.J.; Chen, L.K.; Chang, K.C. Identification and characterisation of the putative phage-related endolysins through full genome sequence analysis in Acinetobacter baumannii ATCC 17978. Int. J. Antimicrob. Agents 2013, 42, 141–148. [Google Scholar] [CrossRef]
- Oliveira, H.; Vilas Boas, D.; Mesnage, S.; Kluskens, L.D.; Lavigne, R.; Sillankorva, S.; Secundo, F.; Azeredo, J. Structural and Enzymatic Characterization of ABgp46, a Novel Phage Endolysin with Broad Anti-Gram-Negative Bacterial Activity. Front. Microbiol. 2016, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.Y.; You, R.I.; Lai, M.J.; Lin, N.T.; Chen, L.K.; Chang, K.C. Highly potent antimicrobial modified peptides derived from the Acinetobacter baumannii phage endolysin LysAB2. Sci. Rep. 2017, 7, 11477. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Cárcamo-Oyarce, G.; Yamamoto, T.; Eisenstein, F.; Hsiao, C.C.; Kurosawa, M.; Gademann, K.; Pilhofer, M.; Nomura, N.; Eberl, L. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat. Commun. 2017, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Schuch, R.; Nelson, D.; Fischetti, V.A. A bacteriolytic agent that detects and kills Bacillus anthracis. Nature 2002, 418, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Silver, L.L. Challenges of antibacterial discovery. Clin. Microbiol. Rev. 2011, 24, 71–109. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Ts2631 | Origin/Characteristics # |
|---|---|---|
| A. baumannii CRAB KPD 205 | 3.71 ± 0.06 2.175 ± 0.51 * 1.66 ± 0.09 ** | Carbapenem-resistant clinical strain (PIP, TZP, CAZ, FEP, IMP, MEM, CIP, LVX, and SXT) |
| A. baumannii MDR KPD 581 | 2.31 ± 0.04 1.07 ± 0.07 * | Multidrug-resistant clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, ETP, MEM, CIP, SXT, and TOB) |
| P. aeruginosa KPD 430 | 2.22 ± 0.16 | Clinical strain (PIP, TZP, CAZ, FEP, CIP, LVX, TCC, and TOB) |
| P. aeruginosa CRPA KPD 431 | 1.36 ± 0.20 | Clinical strain (GM, PIP, TZP, CAZ, FEP, TCC, and MEM) |
| E. coli KPD 217 | 0.35 ± 0.08 | Clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, CSL, and SXT) |
| C. freundii KPD 219 | 0.00 | Clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, CSL, ETP, and SXT) |
| C. braakii KPD 218 | 0.82 ± 0.06 0.63 ± 0.06 * | Clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, CSL, ETP, and SXT) |
| K. pneumoniae KPD 298 | 0.11 ± 0.03 | Clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, ETP, IMP, MEM, AKN, CIP, SXT, and TOB) |
| E. cloacae KPD 297 | 0.00 | Clinical strain (AMP, AMC, TZP, CEP, CXM, FOX, CTX, CAZ, FEP, CSL, ETP, CIP, and SXT) |
| Bacterial Species | K/PO4/EDTA | Ts2631/EDTA | K/PO4/Malic | Ts2631/Malic | K/PO4/Citric | Ts2631/Citric |
|---|---|---|---|---|---|---|
| A. baumannii CRAB* KPD 205 | 2.20 ± 0.05 | 4.24 ± 0.40 | 0.00 | 2.93 ± 0.12 | 0.00 | 3.30 ± 0.09 |
| A. baumannii MDR* KPD 581 | 2.77 ± 0.05 | 3.93 ± 0.05 | 0.00 | 1.83 ± 0.11 | 0.00 | 2.16 ± 0.26 |
| P. aeruginosa KPD 430 | >6.00 | 0.00 | 0.81 ± 0.20 | 0.00 | 2.56 ± 0.06 | |
| P. aeruginosa CRPA KPD 431 | >6.00 | 0.00 | 0.58 ± 0.14 | 0.00 | 1.39 ± 0.05 | |
| E. coli KPD 217 | 0.68 ± 0.07 | 3.08 ± 0.11 | 0.00 | 0.65 ± 0.16 | 0.00 | 0.98 ± 0.08 |
| C. freundii KPD 219 | 0.78 ± 0.06 | 3.45 ± 0.37 | 0.00 | 0.00 | 0.00 | 0.05 ± 0.02 |
| C. braakii KPD 218 | 0.94 ± 0.07 | >6.00 | 0.25 ± 0.12 | 0.94 ± 0.07 | 0.33 ± 0.14 | 1.31 ± 0.08 |
| C. braakii KPD 218 * | 0.84 ± 0.08 | 5.47 ± 0.08 | 0.00 | 0.77 ± 0.18 | 0.00 | 1.00 ± 0.05 |
| K. pneumoniae KPD 298 | 0.24 ± 0.09 | 1.18 ± 0.02 | 0.00 | 0.00 | 0.00 | 0.00 |
| E. cloacae KPD 297 | 0.40 ± 0.13 | 3.85 ± 0.04 | 0.00 | 0.00 | 0.00 | 0.14 ± 0.02 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plotka, M.; Kapusta, M.; Dorawa, S.; Kaczorowska, A.-K.; Kaczorowski, T. Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses 2019, 11, 657. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070657
Plotka M, Kapusta M, Dorawa S, Kaczorowska A-K, Kaczorowski T. Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria. Viruses. 2019; 11(7):657. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070657
Chicago/Turabian StylePlotka, Magdalena, Malgorzata Kapusta, Sebastian Dorawa, Anna-Karina Kaczorowska, and Tadeusz Kaczorowski. 2019. "Ts2631 Endolysin from the Extremophilic Thermus scotoductus Bacteriophage vB_Tsc2631 as an Antimicrobial Agent against Gram-Negative Multidrug-Resistant Bacteria" Viruses 11, no. 7: 657. https://0-doi-org.brum.beds.ac.uk/10.3390/v11070657