Pulmonary Involvement during the Ebola Virus Disease

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
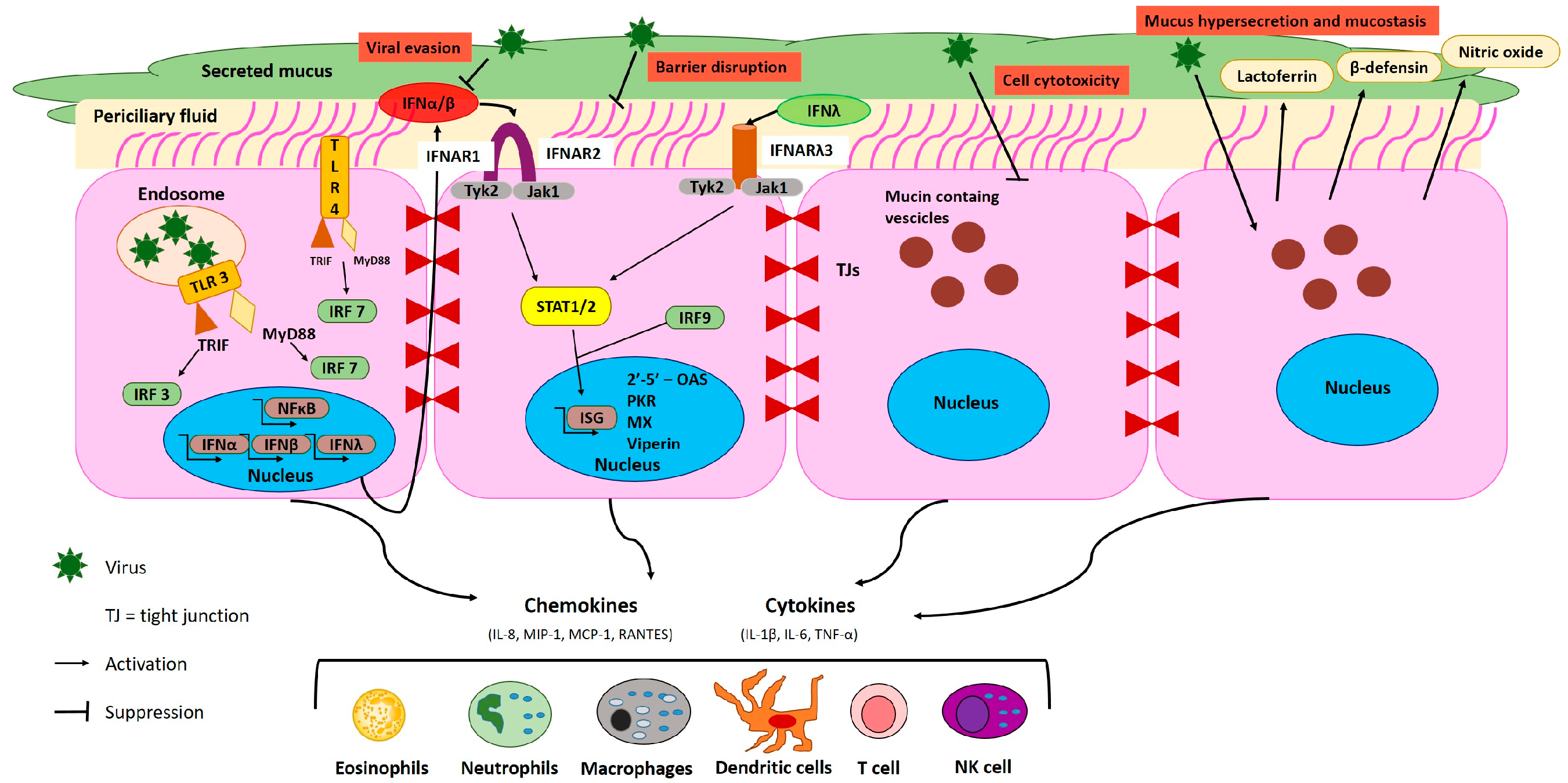
2. Host Defense Mechanisms in the Respiratory Tract during EBOV Infection
3. Ebola Virus Disease
3.1. Animal Studies
3.2. Clinical and Pathology Findings in Humans during EVD
3.3. Impact of Epidemiological and Virological Data on Infection and Control Measures
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldstein, T.; Anthony, S.J.; Gbakima, A.; Bird, B.H.; Bangura, J.; Tremeau-Bravard, A.; Belaganahalli, M.N.; Wells, H.L.; Dhanota, J.K.; Liang, E.; et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat. Microbiol. 2018, 3, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, J.H.; Adachi, T.; Adhikari, N.K.J.; Arribas, J.R.; Bah, I.E.; Bausch, D.G.; Bhadelia, N.; Borchert, M.; Brantsæter, A.B.; Brett-Major, D.M.; et al. New filovirus disease classification and nomenclature. Nat. Rev. Microbiol. 2019, 17, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Baize, S.; Pannetier, D.; Oestereich, L.; Rieger, T.; Koivogui, L.; Magassouba, N.; Soropogui, B.; Sow, M.S.; Keïta, S.; De Clerck, H.; et al. Emergence of Zaire Ebola virus disease in Guinea. N. Engl. J. Med. 2014, 371, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, H.; Sanchez, A.T.W.G. Filoviridae: Marburg and Ebola Viruses. In Fields Virology, 6th ed.; Wolters Kluwer Health Adis/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; Volume 1, pp. 923–956. [Google Scholar]
- CDC. Ebola History. Available online: https://www.cdc.gov/vhf/ebola/history/2014-2016-outbreak/index.html (accessed on 23 August 2019).
- World Health Organization (WHO). Ebola Situation Reports. Available online: https://www.who.int/ebola/situation-reports/drc-2018/en/ (accessed on 23 August 2019).
- Atherstone, C.; Smith, E.; Ochungo, P.; Roesel, K.; Grace, D. Assessing the Potential Role of Pigs in the Epidemiology of Ebola Virus in Uganda. Transbound. Emerg. Dis. 2017, 64, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Bausch, D.G.; Towner, J.S.; Dowell, S.F.; Kaducu, F.; Lukwiya, M.; Sanchez, A.; Nichol, S.T.; Ksiazek, T.G.; Rollin, P.E. Assessment of the risk of Ebola virus transmission from bodily fluids and fomites. J. Infect. Dis. 2007, 196 (Suppl. S2), S142–S147. [Google Scholar] [CrossRef] [PubMed]
- Martines, R.B.; Ng, D.L.; Greer, P.W.; Rollin, P.E.; Zaki, S.R. Tissue and cellular tropism, pathology and pathogenesis of Ebola and Marburg viruses. J. Pathol. 2015, 235, 153–174. [Google Scholar] [CrossRef]
- Osterholm, M.T.; Moore, K.A.; Kelley, N.S.; Brosseau, L.M.; Wong, G.; Murphy, F.A.; Peters, C.J.; LeDuc, J.W.; Russell, P.K.; Van Herp, M.; et al. Transmission of Ebola Viruses: What We Know and What We Do Not Know. MBio 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Vareille, M.; Kieninger, E.; Edwards, M.R.; Regamey, N. The Airway Epithelium: Soldier in the Fight against Respiratory Viruses. Clin. Microbiol. Rev. 2011, 24, 210–229. [Google Scholar] [CrossRef] [Green Version]
- Hiemstra, P.S. Epithelial antimicrobial peptides and proteins: Their role in host defence and inflammation. Paediatr. Respir. Rev. 2001, 2, 306–310. [Google Scholar] [CrossRef]
- Zaki, S.; Blau, D.M.; Hughes, J.M.; Nolte, K.B.; Lynfield, R.; Carr, W.; Popovic, T. Centers for Disease Control and Prevention (CDC) CDC Grand Rounds: Discovering new diseases via enhanced partnership between public health and pathology experts. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 121–126. [Google Scholar]
- Biava, M.; Caglioti, C.; Bordi, L.; Castilletti, C.; Colavita, F.; Quartu, S.; Nicastri, E.; Lauria, F.N.; Petrosillo, N.; Lanini, S.; et al. Detection of Viral RNA in Tissues following Plasma Clearance from an Ebola Virus Infected Patient. PLoS Pathog. 2017, 13, e1006065. [Google Scholar] [CrossRef]
- Hensley, L.E.; Young, H.A.; Jahrling, P.B.; Geisbert, T.W. Proinflammatory response during Ebola virus infection of primate models: Possible involvement of the tumor necrosis factor receptor superfamily. Immunol. Lett. 2002, 80, 169–179. [Google Scholar] [CrossRef]
- Ströher, U.; West, E.; Bugany, H.; Klenk, H.D.; Schnittler, H.J.; Feldmann, H. Infection and activation of monocytes by Marburg and Ebola viruses. J. Virol. 2001, 75, 11025–11033. [Google Scholar] [CrossRef]
- Kerber, R.; Krumkamp, R.; Korva, M.; Rieger, T.; Wurr, S.; Duraffour, S.; Oestereich, L.; Gabriel, M.; Sissoko, D.; Anglaret, X.; et al. Kinetics of Soluble Mediators of the Host Response in Ebola Virus Disease. J. Infect. Dis. 2018, 218, S496–S503. [Google Scholar] [CrossRef] [Green Version]
- McElroy, A.K.; Akondy, R.S.; Davis, C.W.; Ellebedy, A.H.; Mehta, A.K.; Kraft, C.S.; Lyon, G.M.; Ribner, B.S.; Varkey, J.; Sidney, J.; et al. Human Ebola virus infection results in substantial immune activation. Proc. Natl. Acad. Sci. USA 2015, 112, 4719–4724. [Google Scholar] [CrossRef] [Green Version]
- Colavita, F.; Biava, M.; Castilletti, C.; Lanini, S.; Miccio, R.; Portella, G.; Vairo, F.; Ippolito, G.; Capobianchi, M.R.; Di Caro, A.; et al. Inflammatory and Humoral Immune Response during Ebola Virus Infection in Survivor and Fatal Cases Occurred in Sierra Leone during the 2014–2016 Outbreak in West Africa. Viruses 2019, 11, 373. [Google Scholar] [CrossRef]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Wauquier, N.; Becquart, P.; Padilla, C.; Baize, S.; Leroy, E.M. Human fatal zaire ebola virus infection is associated with an aberrant innate immunity and with massive lymphocyte apoptosis. PLoS Negl. Trop. Dis. 2010, 4, e837. [Google Scholar] [CrossRef]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Lubaki, N.M.; Ilinykh, P.; Pietzsch, C.; Tigabu, B.; Freiberg, A.N.; Koup, R.A.; Bukreyev, A. The lack of maturation of Ebola virus-infected dendritic cells results from the cooperative effect of at least two viral domains. J. Virol. 2013, 87, 7471–7485. [Google Scholar] [CrossRef]
- Bosio, C.M.; Aman, M.J.; Grogan, C.; Hogan, R.; Ruthel, G.; Negley, D.; Mohamadzadeh, M.; Bavari, S.; Schmaljohn, A. Ebola and Marburg viruses replicate in monocyte-derived dendritic cells without inducing the production of cytokines and full maturation. J. Infect. Dis. 2003, 188, 1630–1638. [Google Scholar] [CrossRef]
- McElroy, A.K.; Mühlberger, E.; Muñoz-Fontela, C. Immune barriers of Ebola virus infection. Curr. Opin. Virol. 2018, 28, 152–160. [Google Scholar] [CrossRef]
- Cimini, E.; Viola, D.; Cabeza-Cabrerizo, M.; Romanelli, A.; Tumino, N.; Sacchi, A.; Bordoni, V.; Casetti, R.; Turchi, F.; Martini, F.; et al. Different features of Vδ2 T and NK cells in fatal and non-fatal human Ebola infections. PLoS Negl. Trop. Dis. 2017, 11, 11. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Bryniarski, K.; Nazimek, K. Ebola haemorrhagic fever virus: Pathogenesis, immune responses, potential prevention. Folia Med. Cracov. 2014, 54, 39–48. [Google Scholar]
- Malvy, D.; McElroy, A.K.; de Clerck, H.; Günther, S.; van Griensven, J. Ebola virus disease. Lancet 2019, 393, 936–948. [Google Scholar] [CrossRef] [Green Version]
- Klompas, M.; Yokoe, D.S. The Ebola transmission paradox. Am. J. Infect. Control 2015, 43, 786–787. [Google Scholar] [CrossRef]
- Piercy, T.J.; Smither, S.J.; Steward, J.A.; Eastaugh, L.; Lever, M.S. The survival of filoviruses in liquids, on solid substrates and in a dynamic aerosol. J. Appl. Microbiol. 2010, 109, 1531–1539. [Google Scholar] [CrossRef]
- Jahrling, P.B.; Geisbert, T.W.; Dalgard, D.W.; Johnson, E.D.; Ksiazek, T.G.; Hall, W.C.; Peters, C.J. Preliminary report: Isolation of Ebola virus from monkeys imported to USA. Lancet (Lond. Engl.) 1990, 335, 502–505. [Google Scholar] [CrossRef]
- Barrette, R.W.; Metwally, S.A.; Rowland, J.M.; Xu, L.; Zaki, S.R.; Nichol, S.T.; Rollin, P.E.; Towner, J.S.; Shieh, W.-J.; Batten, B.; et al. Discovery of swine as a host for the Reston ebolavirus. Science 2009, 325, 204–206. [Google Scholar] [CrossRef]
- Marsh, G.A.; Haining, J.; Robinson, R.; Foord, A.; Yamada, M.; Barr, J.A.; Payne, J.; White, J.; Yu, M.; Bingham, J.; et al. Ebola Reston virus infection of pigs: Clinical significance and transmission potential. J. Infect. Dis. 2011, 204, S804–S809. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, W.; Cui, L.; Hua, X.; Wang, M.; Zeng, Q. Reston virus in domestic pigs in China. Arch. Virol. 2014, 159, 1129–1132. [Google Scholar] [CrossRef]
- Johnson, E.; Jaax, N.; White, J.; Jahrling, P. Lethal experimental infections of rhesus monkeys by aerosolized Ebola virus. Int. J. Exp. Pathol. 1995, 76, 227–236. [Google Scholar]
- Kobinger, G.P.; Leung, A.; Neufeld, J.; Richardson, J.S.; Falzarano, D.; Smith, G.; Tierney, K.; Patel, A.; Weingartl, H.M. Replication, pathogenicity, shedding, and transmission of Zaire ebolavirus in pigs. J. Infect. Dis. 2011, 204, 200–208. [Google Scholar] [CrossRef]
- Weingartl, H.M.; Embury-Hyatt, C.; Nfon, C.; Leung, A.; Smith, G.; Kobinger, G. Transmission of Ebola virus from pigs to non-human primates. Sci. Rep. 2012, 2, 4. [Google Scholar] [CrossRef]
- Nfon, C.K.; Leung, A.; Smith, G.; Embury-Hyatt, C.; Kobinger, G.; Weingartl, H.M. Immunopathogenesis of Severe Acute Respiratory Disease in Zaire ebolavirus-Infected Pigs. PLoS ONE 2013, 8, e61904. [Google Scholar] [CrossRef]
- Twenhafel, N.A.; Mattix, M.E.; Johnson, J.C.; Robinson, C.G.; Pratt, W.D.; Cashman, K.A.; Wahl-Jensen, V.; Terry, C.; Olinger, G.G.; Hensley, L.E.; et al. Pathology of Experimental Aerosol Zaire Ebolavirus Infection in Rhesus Macaques. Vet. Pathol. 2013, 50, 514–529. [Google Scholar] [CrossRef]
- Bulletin of the World Health Organization Vols. 1 to 97. 1948 to 2019. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/journals/522/ (accessed on 23 August 2019).
- Heymann, D.L.; Weisfeld, J.S.; Webb, P.A.; Johnson, K.M.; Cairns, T.; Berquist, H. Ebola hemorrhagic fever: Tandala, Zaire, 1977–1978. J. Infect. Dis. 1980, 142, 372–376. [Google Scholar] [CrossRef]
- Baron, R.C.; McCormick, J.B.; Zubeir, O.A. Ebola virus disease in southern Sudan: Hospital dissemination and intrafamilial spread. Bull. World Health Organ. 1983, 61, 997–1003. [Google Scholar]
- Miranda, M.E.; White, M.E.; Dayrit, M.M.; Hayes, C.G.; Ksiazek, T.G.; Burans, J.P. Seroepidemiological study of filovirus related to Ebola in the Philippines. Lancet (Lond. Engl.) 1991, 337, 425–426. [Google Scholar] [CrossRef] [Green Version]
- Epidemiologic Notes and Reports Update: Ebola-Related Filovirus Infection in Nonhuman Primates and Interim Guidelines for Handling Nonhuman Primates during Transit and Quarantine. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/00001538.htm (accessed on 23 August 2019).
- Le Guenno, B.; Formenty, P.; Formentry, P.; Wyers, M.; Gounon, P.; Walker, F.; Boesch, C. Isolation and partial characterisation of a new strain of Ebola virus. Lancet (Lond. Engl.) 1995, 345, 1271–1274. [Google Scholar] [CrossRef]
- Georges, A.J.; Leroy, E.M.; Renaut, A.A.; Benissan, C.T.; Nabias, R.J.; Ngoc, M.T.; Obiang, P.I.; Lepage, J.P.; Bertherat, E.J.; Bénoni, D.D.; et al. Ebola hemorrhagic fever outbreaks in Gabon, 1994–1997: Epidemiologic and health control issues. J. Infect. Dis. 1999, 179, S65–S75. [Google Scholar] [CrossRef]
- Khan, A.S.; Tshioko, F.K.; Heymann, D.L.; Le Guenno, B.; Nabeth, P.; Kerstiëns, B.; Fleerackers, Y.; Kilmarx, P.H.; Rodier, G.R.; Nkuku, O.; et al. The reemergence of Ebola hemorrhagic fever, Democratic Republic of the Congo, 1995. Commission de Lutte contre les Epidémies à Kikwit. J. Infect. Dis. 1999, 179, S76–S86. [Google Scholar] [CrossRef]
- Okware, S.I.; Omaswa, F.G.; Zaramba, S.; Opio, A.; Lutwama, J.J.; Kamugisha, J.; Rwaguma, E.B.; Kagwa, P.; Lamunu, M. An outbreak of Ebola in Uganda. Trop. Med. Int. Health 2002, 7, 1068–1075. [Google Scholar] [CrossRef]
- The Weekly Epidemiological Record (WER). Available online: https://www.who.int/wer/en/ (accessed on 23 August 2019).
- Formenty, P.; Libama, F.; Epelboin, A.; Allarangar, Y.; Leroy, E.; Moudzeo, H.; Tarangonia, P.; Molamou, A.; Lenzi, M.; Ait-Ikhlef, K.; et al. Outbreak of Ebola hemorrhagic fever in the Republic of the Congo, 2003: A new strategy? Med. Trop. (Mars) 2003, 63, 291–295. [Google Scholar]
- MacNeil, A.; Rollin, P.E. Ebola and Marburg Hemorrhagic Fevers: Neglected Tropical Diseases? PLoS Negl. Trop. Dis. 2012, 6, e1546. [Google Scholar] [CrossRef]
- Global Alert and Response. Available online: https://www.who.int/ihr/global_alert/en/ (accessed on 23 August 2019).
- Shoemaker, T.; MacNeil, A.; Balinandi, S.; Campbell, S.; Wamala, J.F.; McMullan, L.K.; Downing, R.; Lutwama, J.; Mbidde, E.; Ströher, U.; et al. Reemerging Sudan Ebola Virus Disease in Uganda, 2011. Emerg. Infect. Dis. 2012, 18, 1480. [Google Scholar] [CrossRef]
- Albariño, C.G.; Uebelhoer, L.S.; Vincent, J.P.; Khristova, M.L.; Chakrabarti, A.K.; McElroy, A.; Nichol, S.T.; Towner, J.S. Development of a reverse genetics system to generate recombinant Marburg virus derived from a bat isolate. Virology 2013, 446, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Maganga, G.D.; Kapetshi, J.; Berthet, N.; Kebela Ilunga, B.; Kabange, F.; Mbala Kingebeni, P.; Mondonge, V.; Muyembe, J.-J.T.; Bertherat, E.; Briand, S.; et al. Ebola Virus Disease in the Democratic Republic of Congo. N. Engl. J. Med. 2014, 371, 2083–2091. [Google Scholar] [CrossRef]
- Nsio, J.; Kapetshi, J.; Makiala, S.; Raymond, F.; Tshapenda, G.; Boucher, N.; Corbeil, J.; Okitandjate, A.; Mbuyi, G.; Kiyele, M.; et al. 2017 Outbreak of Ebola Virus Disease in Northern Democratic Republic of Congo. J. Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Bai, C.Q.; Mu, J.S.; Kargbo, D.; Bin Song, Y.; Niu, W.K.; Nie, W.M.; Kanu, A.; Liu, W.W.; Wang, Y.P.; Dafae, F.; et al. Clinical and Virological Characteristics of Ebola Virus Disease Patients Treated with Favipiravir (T-705)—Sierra Leone, 2014. Clin. Infect. Dis. 2016, 63, 1288–1294. [Google Scholar] [CrossRef]
- Cantoni, D.; Hamlet, A.; Michaelis, M.; Wass, M.N.; Rossman, J.S. Risks Posed by Reston, the Forgotten Ebolavirus. mSphere 2016, 1. [Google Scholar] [CrossRef]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The Pathogenesis of Ebola Virus Disease. Annu. Rev. Pathol. 2017, 12, 387–418. [Google Scholar] [CrossRef]
- St Claire, M.C.; Ragland, D.R.; Bollinger, L.; Jahrling, P.B. Animal Models of Ebolavirus Infection. Comp. Med. 2017, 67, 253–262. [Google Scholar]
- Zaki, S.R.; Goldsmith, C.S. Pathologic features of filovirus infections in humans. Curr. Top. Microbiol. Immunol. 1999, 235, 97–116. [Google Scholar]
- Caswell, J.L.; Williams, K.J. Jubb, Kennedy and Palmer’s Pathology of Domestic Animals; Saunders Ltd.: Philadelphia, PA, USA, 2007. [Google Scholar]
- Irving, W.L. Ebola virus transmission. Int. J. Exp. Pathol. 1995, 76, 225–226. [Google Scholar]
- WHO/International Study Team. Ebola haemorrhagic fever in Sudan, 1976. Bull. World Health Organ. 1978, 56, 247–270. [Google Scholar]
- Jones, R.M.; Brosseau, B.L. Ebola virus transmission via contact and aerosol—A new paradigm. Cent. Infect. Dis. Res. Policy 2014. Available online: http://www.cidrap.umn.edu/news-perspective/2014/11/commentary-ebola-virus-transmission-contact-and-aerosol-new-paradigm (accessed on 23 August 2019).
- Schieffelin, J.S.; Shaffer, J.G.; Goba, A.; Gbakie, M.; Gire, S.K.; Colubri, A.; Sealfon, R.S.G.; Kanneh, L.; Moigboi, A.; Momoh, M.; et al. Clinical illness and outcomes in patients with Ebola in Sierra Leone. N. Engl. J. Med. 2014, 371, 2092–2100. [Google Scholar] [CrossRef]
- Nanclares, C.; Kapetshi, J.; Lionetto, F.; de la Rosa, O.; Tamfun, J.-J.M.; Alia, M.; Kobinger, G.; Bernasconi, A. Ebola Virus Disease, Democratic Republic of the Congo, 2014. Emerg. Infect. Dis. 2016, 22, 1579–1586. [Google Scholar] [CrossRef]
- Dunning, J.; Sahr, F.; Rojek, A.; Gannon, F.; Carson, G.; Idriss, B.; Massaquoi, T.; Gandi, R.; Joseph, S.; Osman, H.K.; et al. Experimental Treatment of Ebola Virus Disease with TKM-130803: A Single-Arm Phase 2 Clinical Trial. PLoS Med. 2016, 13, 19. [Google Scholar] [CrossRef]
- Ji, Y.J.; Duan, X.Z.; Gao, X.D.; Li, L.; Li, C.; Ji, D.; Li, W.G.; Wang, L.F.; Meng, Y.H.; Yang, X.; et al. Clinical presentations and outcomes of patients with Ebola virus disease in Freetown, Sierra Leone. Infect. Dis. Poverty 2016, 5, 10. [Google Scholar] [CrossRef]
- Qureshi, A.I.; Abd-Allah, F.; Aleu, A.; Connors, J.J.; Hanel, R.A.; Hassan, A.E.; Hussein, H.M.; Janjua, N.A.; Khatri, R.; Kirmani, J.F.; et al. Endovascular treatment for acute ischemic stroke patients: Implications and interpretation of IMS III, MR RESCUE, and SYNTHESIS EXPANSION trials: A report from the Working Group of International Congress of Interventional Neurology. J. Vasc. Interv. Neurol. 2014, 7, 56–75. [Google Scholar]
- Ingelbeen, B.; Bah, E.I.; Decroo, T.; Balde, I.; Nordenstedt, H.; van Griensven, J.; De Weggheleire, A. Mortality among PCR negative admitted Ebola suspects during the 2014/15 outbreak in Conakry, Guinea: A retrospective cohort study. PLoS ONE 2017, 12, e0180070. [Google Scholar] [CrossRef]
- Kangbai, J.B.; Heumann, C.; Hoelscher, M.; Sahr, F.; Froeschl, G. Epidemiological characteristics, clinical manifestations, and treatment outcome of 139 paediatric Ebola patients treated at a Sierra Leone Ebola treatment center. BMC Infect. Dis. 2019, 19, 81. [Google Scholar] [CrossRef]
- Rojek, A.; Horby, P.; Dunning, J. Insights from clinical research completed during the west Africa Ebola virus disease epidemic. Lancet. Infect. Dis. 2017, 17, e280–e292. [Google Scholar] [CrossRef] [Green Version]
- Van Griensven, J.; Bah, E.I.; Haba, N.; Delamou, A.; Camara, B.S.; Olivier, K.J.-J.; De Clerck, H.; Nordenstedt, H.; Semple, M.G.; Van Herp, M.; et al. Electrolyte and Metabolic Disturbances in Ebola Patients during a Clinical Trial, Guinea, 2015. Emerg. Infect. Dis. 2016, 22, 2120. [Google Scholar] [CrossRef]
- Uyeki, T.M.; Mehta, A.K.; Davey, R.T.; Liddell, A.M.; Wolf, T.; Vetter, P.; Schmiedel, S.; Grünewald, T.; Jacobs, M.; Arribas, J.R.; et al. Clinical Management of Ebola Virus Disease in the United States and Europe. N. Engl. J. Med. 2016, 374, 636–646. [Google Scholar] [CrossRef] [Green Version]
- Petrosillo, N.; Nicastri, E.; Lanini, S.; Capobianchi, M.R.; Di Caro, A.; Antonini, M.; Puro, V.; Lauria, F.N.; Shindo, N.; Magrini, N.; et al. Ebola virus disease complicated with viral interstitial pneumonia: A case report. BMC Infect. Dis. 2015, 15, 432. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Josse, R.; Johnson, E.D.; Merlin, M.; Georges, A.J.; Abandja, J.; Danyod, M.; Delaporte, E.; Dupont, A.; Ghogomu, A. Antibody prevalence against haemorrhagic fever viruses in randomized representative Central African populations. Res. Virol. 1989, 140, 319–331. [Google Scholar] [CrossRef]
- Roels, T.H.; Bloom, A.S.; Buffington, J.; Muhungu, G.L.; Mac Kenzie, W.R.; Khan, A.S.; Ndambi, R.; Noah, D.L.; Rolka, H.R.; Peters, C.J.; et al. Ebola hemorrhagic fever, Kikwit, Democratic Republic of the Congo, 1995: Risk factors for patients without a reported exposure. J. Infect. Dis. 1999, 179, S92–S97. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention Guidance for Donning and Doffing Personal Protective Equipment (PPE) During Management of Patients with Ebola Virus Disease in U.S. Hospitals. Available online: www.cdc.gov/vhf/ebola/hcp/ppe-training/index.html (accessed on 23 August 2019).
- Interim Infection Prevention and Control Guidance for Care of Patients with Suspected or Confirmed Filovirus Haemorrhagic Fever in Health-Care Settings, with Focus on Ebola; World Health Organization: Geneva, Switzerland, 2014; Available online: http://www.euro.who.int/__data/assets/pdf_file/0005/268772/Interim-Infection-Prevention-and-Control-Guidance-for-Care-of-Patients-with-Suspected-or-Confirmed-Filovirus-Haemorrhagic-Fever-in-Health-Care-Settings,-with-Focus-on-Ebola-Eng.pdf (accessed on 23 August 2019).
- World Health Organization. World Health Report 2004 Statistical Anne; WHO: Geneva, Switzerland, 2004. [Google Scholar]
- Mahony, J.B. Nucleic acid amplification-based diagnosis of respiratory virus infections. Expert Rev. Anti. Infect. Ther. 2010, 8, 1273–1292. [Google Scholar] [CrossRef]
- Judson, S.; Prescott, J.; Munster, V. Understanding Ebola virus transmission. Viruses 2015, 7, 511–521. [Google Scholar] [CrossRef]
- Gonzalez, J.P.; Nakoune, E.; Slenczka, W.; Vidal, P.; Morvan, J.M. Ebola and Marburg virus antibody prevalence in selected populations of the Central African Republic. Microbes Infect. 2000, 2, 39–44. [Google Scholar] [CrossRef]
- Khanal, S.; Ghimire, P.; Dhamoon, A. The Repertoire of Adenovirus in Human Disease: The Innocuous to the Deadly. Biomedicines 2018, 6, 30. [Google Scholar] [CrossRef]
- Falsey, A.R. Respiratory syncytial virus infection in adults. Semin. Respir. Crit. Care Med. 2007, 28, 171–181. [Google Scholar] [CrossRef]
- Pierangeli, A.; Scagnolari, C.; Antonelli, G. Respiratory syncytial virus. Minerva Pediatr. 2018, 70, 553–565. [Google Scholar] [CrossRef]
- Yanagi, Y.; Takeda, M.; Ohno, S. Measles virus: Cellular receptors, tropism and pathogenesis. J. Gen. Virol. 2006, 87, 2767–2779. [Google Scholar] [CrossRef]
- Capelozzi, V.L.; Parra, E.R.; Ximenes, M.; Bammann, R.H.; Barbas, C.S.V.; Duarte, M.I.S. Pathological and ultrastructural analysis of surgical lung biopsies in patients with swine-origin influenza type A/H1N1 and acute respiratory failure. Clinics (Sao Paulo) 2010, 65, 1229–1237. [Google Scholar] [CrossRef] [Green Version]
- Lau, Y.L.; Peiris, J.S.M. Pathogenesis of severe acute respiratory syndrome. Curr. Opin. Immunol. 2005, 17, 404–410. [Google Scholar] [CrossRef]
- Paessler, S.; Walker, D.H. Pathogenesis of the viral hemorrhagic fevers. Annu. Rev. Pathol. 2013, 8, 411–440. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Su, C.-Y.; Chow, N.-H.; Lai, W.-W.; Lei, H.-Y.; Chang, C.-L.; Chang, T.-Y.; Chen, S.-H.; Lin, Y.-S.; Yeh, T.-M.; et al. Dengue viruses can infect human primary lung epithelia as well as lung carcinoma cells, and can also induce the secretion of IL-6 and RANTES. Virus Res. 2007, 126, 216–225. [Google Scholar] [CrossRef]
- Yun, N.E.; Walker, D.H. Pathogenesis of Lassa Fever. Viruses 2012, 4, 2031–2048. [Google Scholar] [CrossRef]
- Safronetz, D.; Prescott, J.; Feldmann, F.; Haddock, E.; Rosenke, R.; Okumura, A.; Brining, D.; Dahlstrom, E.; Porcella, S.F.; Ebihara, H.; et al. Pathophysiology of hantavirus pulmonary syndrome in rhesus macaques. Proc. Natl. Acad. Sci. USA 2014, 111, 7114–7119. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Year | Animal | Virus (Co-Infection; Provenance) | Analyzed Tissue | Route of Inoculation | Pathological Findings | Clinical Signs | Reference |
|---|---|---|---|---|---|---|---|
| 1989 | Cynomolgus monkeys (Macaca fascicularis) | RESTV (co-infection with SHFV; Philippines) | Plasma, Sera, tissues | Natural infection | Enlarged spleens and kidneys | Anorexia, cough, nasal exudates, swollen eyelids | Jahrling PB, et al., Lancet 1990 [31] |
| 2008 | Domestic swine | RESTV (co-infection with PRRSV and PCV-2; Philippines) | Plasma, sera, tissues (lymph nodes) | Natural infection | RESTV isolation from lung and lymph nodes | Severe respiratory disease syndrome, interstitial pneumonia | Barrette RW, et al., Science 2009 [32] |
| 2011 | Domestic pigs | RESTV (Philippines swine isolate) | Blood, swabs, tissues | Challenge by subcutaneous and oral routes | RESTV isolation from superficial (submandibular, axillary, inguinal) and internal (bronchial, mesenteric) lymph nodes, nasal turbinates, muscle, and lung | Mild acute rhinitis Acute bronchopneumonia | Marsh GA, et al., JID 2011 [33] |
| 2014 | Domestic pigs | RESTV (co-infection with PRRSV, China) | Spleen | Natural infection | Positive for RESTV RT-PCR | Typical clinical signs of porcine reproductive and respiratory syndrome | Pan Y, et al., Arch Virol 2014 [34] |
| 1995 | Monkeys (Macaca mulatta) | EBOV (EBOV isolate) | Plasma, tissues | Aerosol exposure | Bronchial and bronchiolar epithelium, alveolar pneumocytes, and alveolar macrophages showed positive EBOV antigen staining | Typical signs of EVD. Serosanguineous nasal discharge, subcutaneous haemorrhage | Johnson E et al., Int. J. Exp. Path. 1995 [35] |
| 2010 | Domesticated Landrace pigs | EBOV (EBOV strain Kikwit 95) | Blood, tissues | Intranasal, intraocular and oral routes | Macroscopic pathological changes in lungs. Abundant viral antigen detection in lungs | Most prominent and progressive clinical signs were respiratory | Kobinger G, et al., J Infect Dis 2011 [36] |
| 2012 | Pigs (breed Landrace) and cynomolgus macaques (Macaca fascicularis) | EBOV (EBOV strain Kikwit 95) | Blood, tissues | Oro-nasal inoculation of the pigs. Macaques in close contact with the pigs to evaluate aerosol transmission | Pigs: viral antigens in bronchioles Macaques: EBOV antigen in alveolar, septal macrophages, pneumocytes and endothelial cells | Pigs: broncho-interstitial pneumonia with a lobular pattern Macaques: typical signs of EBOV infection, with damages mainly to the lung and liver | Weingartl HM et al., Sci Rep. 2012 [37] |
| 2013 | Domesticated Landrace pigs | EBOV (EBOV strain kikwit 95) | Blood, tissues | Intranasal, intraocular and oral routes | Pneumonia, distributed primarily in the dorso-caudal lobes, characterized by consolidation and haemorrhage affecting more than 70% of the lung tissue | Typical signs of EBOV infection. An increase in respiratory rate as well as difficult, abdominal breathing, inappetence, weakness and reluctance to move | Nfon CK, et al., Plos One 2013 [38] |
| – | Rhesus macaques (Macaca mulatta) | EBOV (EBOV strain from Zaire 95) | Blood, set of tissues from all major organs | Aerosol exposure | Histologic changes within the lungs included alveolar histiocytosis, alveolar fibrin, and multifocal fibrinoid vasculitis | Typical signs of EBOV infection | Twenhalfel NA, et al., Vet Path 2013 [39] |
| Date | Country | Virus | No of Cases | No of Deaths | CFR | Clinical Evidence | Diagnostic Evidence | Reference |
|---|---|---|---|---|---|---|---|---|
| Jun–Nov 1976 | Sudan | SUDV | 284 | 151 | 53% | Chest pain 153 (83%), Cough 90 (49%) of 183 patients | 2 autoptic findings with proliferative thickening of alveolar septa | WHO. Bull WHO 1978 [40] |
| Aug 1976 | Zaire | EBOV | 318 | 280 | 88% | Cough 36% in 208 deceased patients, 18% in 34 serogically confirmed patients | Clinical evidence | WHO. Bull WHO 1976 [40] |
| Jun 1977 | Zaire | EBOV | 1 | 1 | 100% | No respiratory sign | Clinical evidence | Heymann DL. J Infect Dis 1980 [41] |
| Aug–Sep 1979 | Sudan | SUDV | 34 | 22 | 65% | No respiratory sign | Clinical evidence | Baron RC. Bull WHO 1983 [42] |
| 1989 | Philippine | RESTV | 3 | 0 | 0% | No respiratory sign | Clinical evidence | Miranda ME. Lancet 1991 [43] |
| 1990 | USA | RESTV | 4 | 0 | 0% | No respiratory sign | Clinical evidence | CDC. MMWR 1990 [44] |
| 1994 | Cote d’Ivoire | TAIFV | 4 | 0 | 0% | No respiratory sign | Clinical evidence | Le Guenno B. Lancet 1995 [45] |
| Dec 1994–Feb 1995 | Gabon | EBOV | 52 | 31 | 60% | No respiratory sign | Clinical evidence | Georges AJ. J Infect Dis 1999 [46] |
| May–Jul 1995 | Zaire | EBOV | 315 | 250 | 79% | Dyspnea 55 (25%) of 209 | Clinical evidence | Khan AS. J Infect Dis 1999 [47] |
| Jan–Apr 1996 | Gabon | EBOV | 60 | 45 | 75% | No respiratory sign | Clinical evidence | Georges AJ. J Infect Dis 1999 [46] |
| Jul 1996–Mar 1997 | Gabon | EBOV | 37 | 21 | 57% | No respiratory sign | Clinical evidence | Georges AJ. J Infect Dis 1999 [46] |
| Oct 2000–Jan 2001 | Uganda | SUDV | 425 | 224 | 53% | No data | Clinical evidence | Okware SI. Trop Med Int Health 2002 [48] |
| Oct 2001–Jul 2002 | Gabon, DRC | EBOV | 124 | 96 | 77% | Article not available | No data | WHO. Week Epi Rec 2003 [49] |
| Dec 2002–Apr 2003 | DRC | EBOV | 143 | 128 | 90% | No data | Clinical evidence | Formenty P. Med Trop 2003 [50] |
| Nov–Dec 2003 | DRC | EBOV | 35 | 29 | 83% | No data | Clinical evidence | WHO. Week Epi Rec 2004 [49] |
| Apr–Jun 2004 | Sudan | SUDV | 17 | 7 | 41% | Cough in 11 of 13 cases, 85% | Clinical evidence | WHO. Week Epi Rec 2005 [49] |
| April 2005 | DRC | EBOV | 12 | 10 | 83% | No data | No data | Article not avalaible |
| Aug–Nov 2007 | DRC | EBOV | 264 | 187 | 71% | No data | No data | WHO. Week Epi Rec 2007 [49] |
| Dec 2007–Jan 2008 | Uganda | BDBV | 149 | 37 | 25% | No data | No data | MacNeil AJ. Infect dis 2011 [51] |
| Dec 2008–Feb 2009 | DRC | EBOV | 32 | 15 | 47% | No data | No data | WHO. Glob Aler Resp 2009 [52] |
| May 2011 | Uganda | SUDV | 1 | 1 | 100% | No respiratory symptoms –Respiratory failure | Clinical evidence | Shoemaker T. EID 2012 [53] |
| Jun–Aug 2012 | Uganda | SUDV | 17 | 7 | 41% | No data | No data | Albarino CG. Virol 2013 [54] |
| Jun–Nov 2012 | DRC | BDBV | 35 | 13 | 36% | No data | No data | Albarino CG. Virol 2013 [54] |
| Dec 2013–Jan 2016 | Western Africa | EBOV | 28,616 | 11,310 | 39% | Cough, dyspnoea, pulmonary oedema, pneumonia | Viral replication markers in sputum samples | Baize S. N Engl J Med 2014 [3] |
| Aug–Nov 2014 | DRC | EBOV | 66 | 49 | 74% | Difficult breathing 21.4% Cough 18% | Clinical evidence | Maganga GD. N Eng J Med 2014 [55] |
| May 2017 | DRC | EBOV | 8 | 4 | 50% | Cough 25% | Clinical evidence | Nsio J. J Infect Dis 2019 [56] |
| May–Jul 2018 | DRC | EBOV | 54 | 33 | 61% | Difficult breathing 34.4% | Clinical evidence | The Ebola Outbreak Epidemiology Team Lancet 2019 [38] |
| August 2018–ongoing | DRC | EBOV | 2620 | 1762 | 67% | No data | No data | WHO, 2019 [57] |
| Family | Genus | Virus | Pathological Findings | Most Common Symptoms | Ref |
|---|---|---|---|---|---|
| Adenoviridae | Mastadenovirus | Adenovirus | Interstitial and peribronchial infiltration, Acute bronchiolitis, Necrosis, Haloed basophilic inclusions | Common cold, Laryngitis, tracheobronchitis | Khanal S. et al., Biomedicines 2018 [85] |
| Herpesviridae | Cytomegalovirus | Cytomegalovirus | Interstitial pneumonitis, Intra-alveolar damage, DAD §, Cytomegaly, Eosinophilic intranuclear Cowdry type-B inclusions | Bronchiolitis, Pneumonia * | Falsey AR et al., Semin Respir Crit Care Med. 2007 [86], Pierangeli A Minerva Pediatr. 2018 [87] |
| Paramyxoviridae | Pneumovirus | Respiratory Syncytial Virus | Atelectasis, Mucosal ulcerations, DAD Giant cells pneumonia | Common cold, Bronchiolitis °,*, Pneumonia °,* | Pierangeli A Minerva Pediatr. 2018 [87] |
| Morbillivirus | Measles | Squamous metaplasia of bronchial epithelium, DAD, Multinucleated giant cells | Fever, Sore throat, Tracheobronchitis, Laryngitis | Yanagi Y J Gen Virol. 2006 [88] | |
| Orthomyxoviridae | Influenza | Influenza virus | Tracheobronchitis, Bronchiolitis, DAD, Hemorrhage oedema, Squamous metaplasia of bronchial epithelium | Fever, Laryngitis, Tracheobronchitis | Capelozzi VL Clinics (Sao Paulo). 2010 [89] |
| Coronoviridae | Betacoronavirus | Severe Acute Respiratory Syndrome (SARS) | DAD, Bronchiolar injury, Multinucleated cells Harvey-Comb lung, Acute bronchopneumonia | Bronchitis, Pneumonia | Lau YL Curr Opin Immunol. 2005 [90] |
| Flaviviridae | Flavivirus | Yellow fever virus Dengue virus | Alveolar oedema, Interstitila pneumonitis, DAD, DAH # | Pneumonia | Paessler S Annu Rev Pathol. 2013 [91], Lee YR Virus Res. 2007 [92] |
| Arenaviridae | Arenavirus | Lassa virus Machupo Virus Guanarito Virus | Alveolar oedema, Interstitial pneumonitis, DAD, Bronchopneumonia | Pneumonia | Paessler S Annu Rev Pathol. 2013 [91], Yun NE Viruses. 2012 [93] |
| Buanyaviridae | Bunyavirus | Hantavirus | Alveolar oedema, DAH, Bronchopneumonia | Pneumonia | Paessler S Annu Rev Pathol. 2013 [91], Safronetz D Proc Natl Acad Sci USA. 2014 [94] |
| Nairoviridae | Crimean-Congo Hemorrhagic Fever (CCHF) | Alveolar oedema | Pneumonia | Paessler S Annu Rev Pathol. 2013 [91] | |
| Filoviridae | Ebolavirus | Ebola virus | Pneumonia, Pulmonary oedema, Pulomnary effusion | Cough, Bronchitis, Pneumonia | Paessler S Annu Rev Pathol. 2013 [91], Marcinkiewicz J Folia Med Cracov. 2014 [27], Martines RB J Pathol. 2015 [9] |
| Marburgvirus | Marburgvirus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalle, E.; Biava, M.; Nicastri, E.; Colavita, F.; Di Caro, A.; Vairo, F.; Lanini, S.; Castilletti, C.; Langer, M.; Zumla, A.; et al. Pulmonary Involvement during the Ebola Virus Disease. Viruses 2019, 11, 780. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090780
Lalle E, Biava M, Nicastri E, Colavita F, Di Caro A, Vairo F, Lanini S, Castilletti C, Langer M, Zumla A, et al. Pulmonary Involvement during the Ebola Virus Disease. Viruses. 2019; 11(9):780. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090780
Chicago/Turabian StyleLalle, Eleonora, Mirella Biava, Emanuele Nicastri, Francesca Colavita, Antonino Di Caro, Francesco Vairo, Simone Lanini, Concetta Castilletti, Martin Langer, Alimuddin Zumla, and et al. 2019. "Pulmonary Involvement during the Ebola Virus Disease" Viruses 11, no. 9: 780. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090780