Identification of Two Porcine Reproductive and Respiratory Syndrome Virus Variants Sharing High Genomic Homology but with Distinct Virulence

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Differential Detection
2.2. Virus Isolation and Genome Sequencing
2.3. Multiple Alignments, Phylogenetic and Recombination Analyses
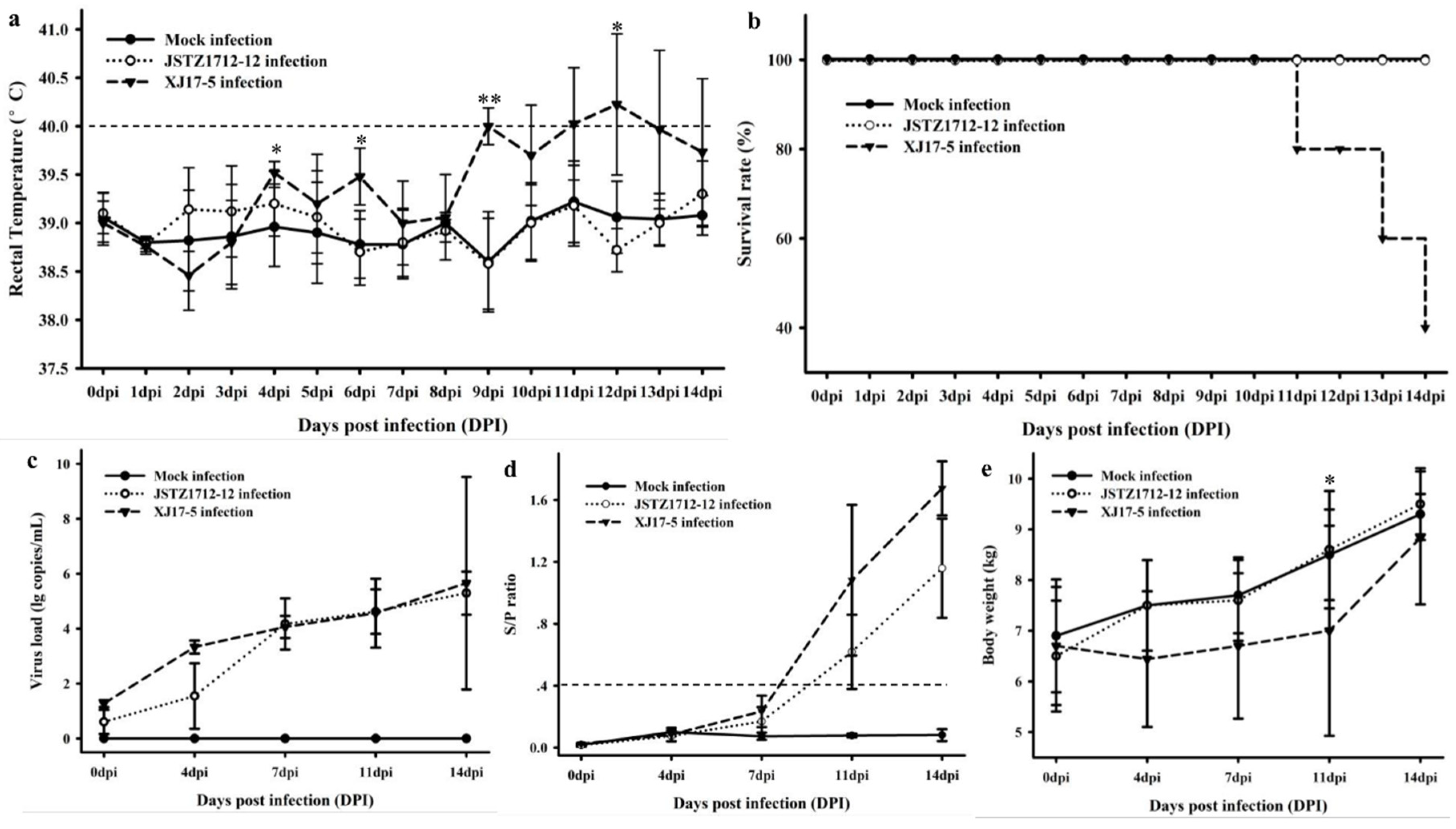
2.4. Animal Challenge Study
2.5. Statistical Analysis
3. Results
3.1. Clinical Sample Detection
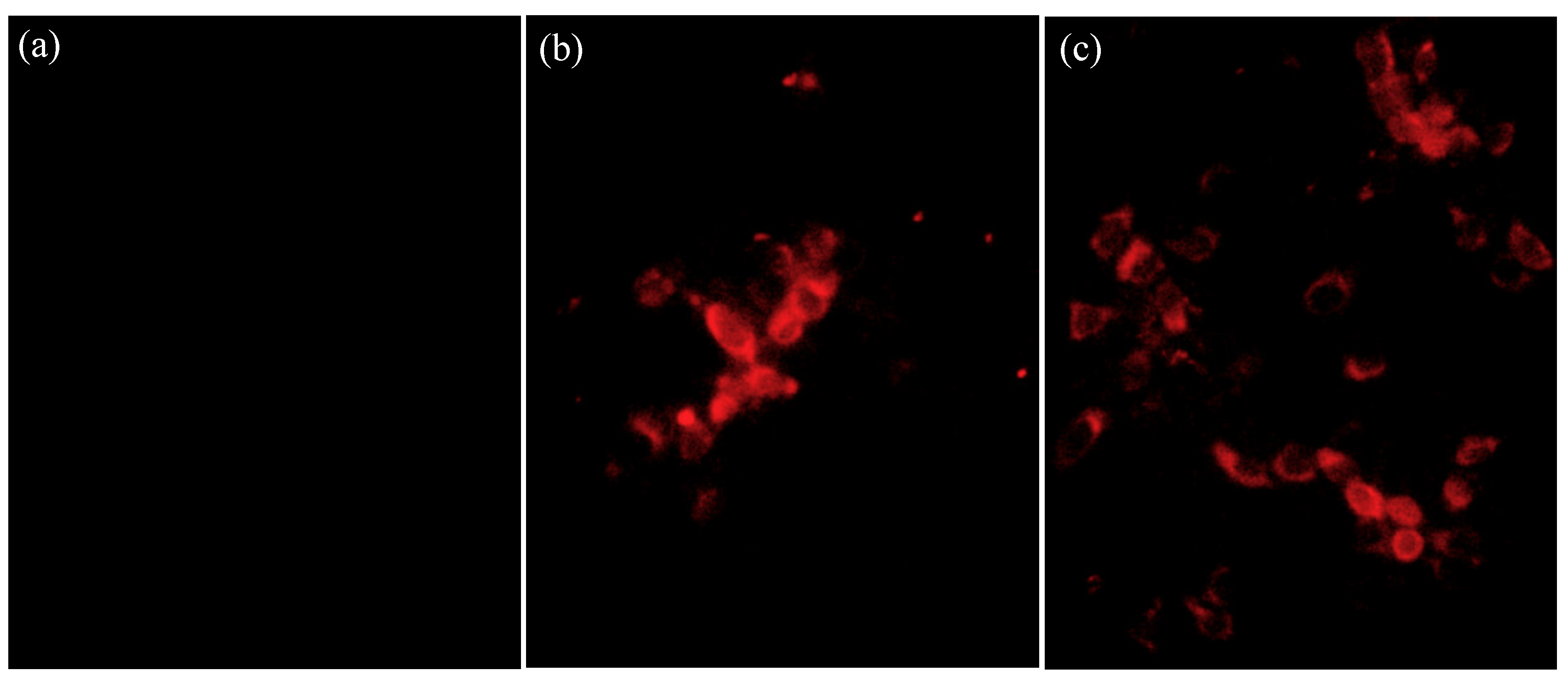
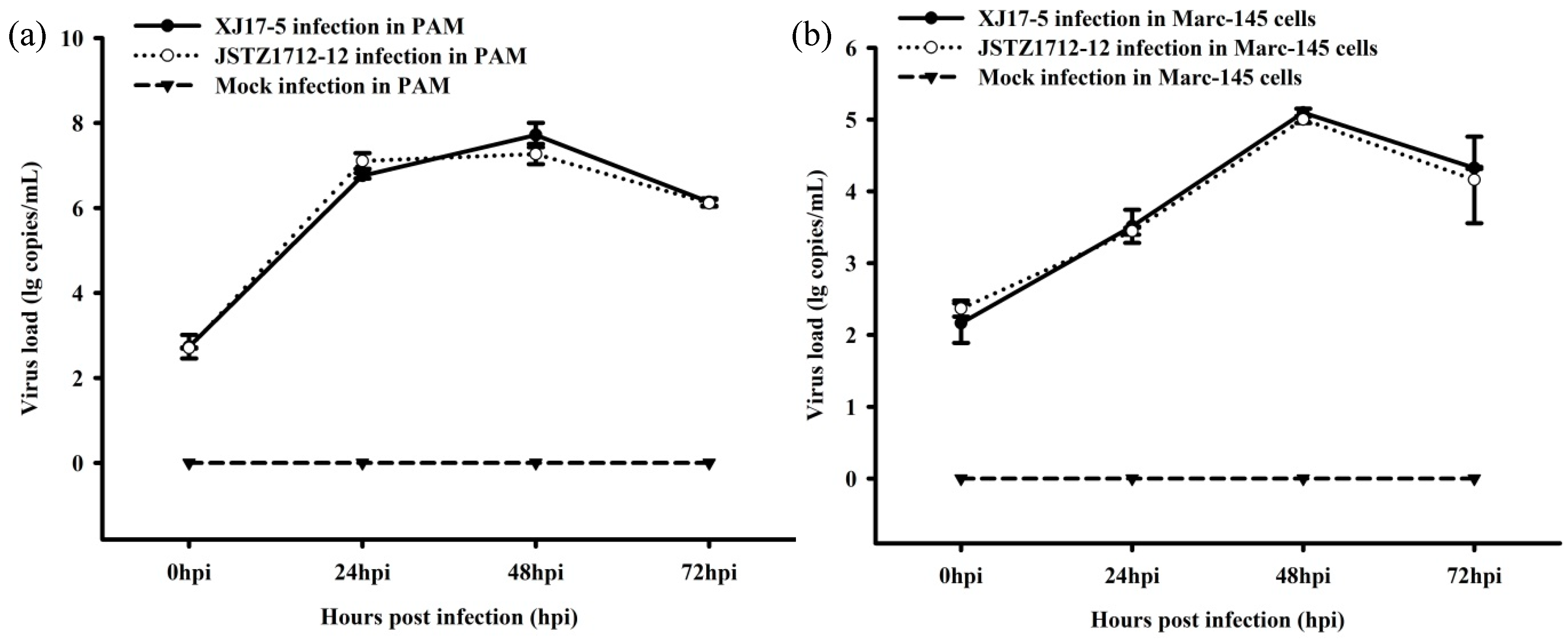
3.2. Virus Isolation and Growth Curve
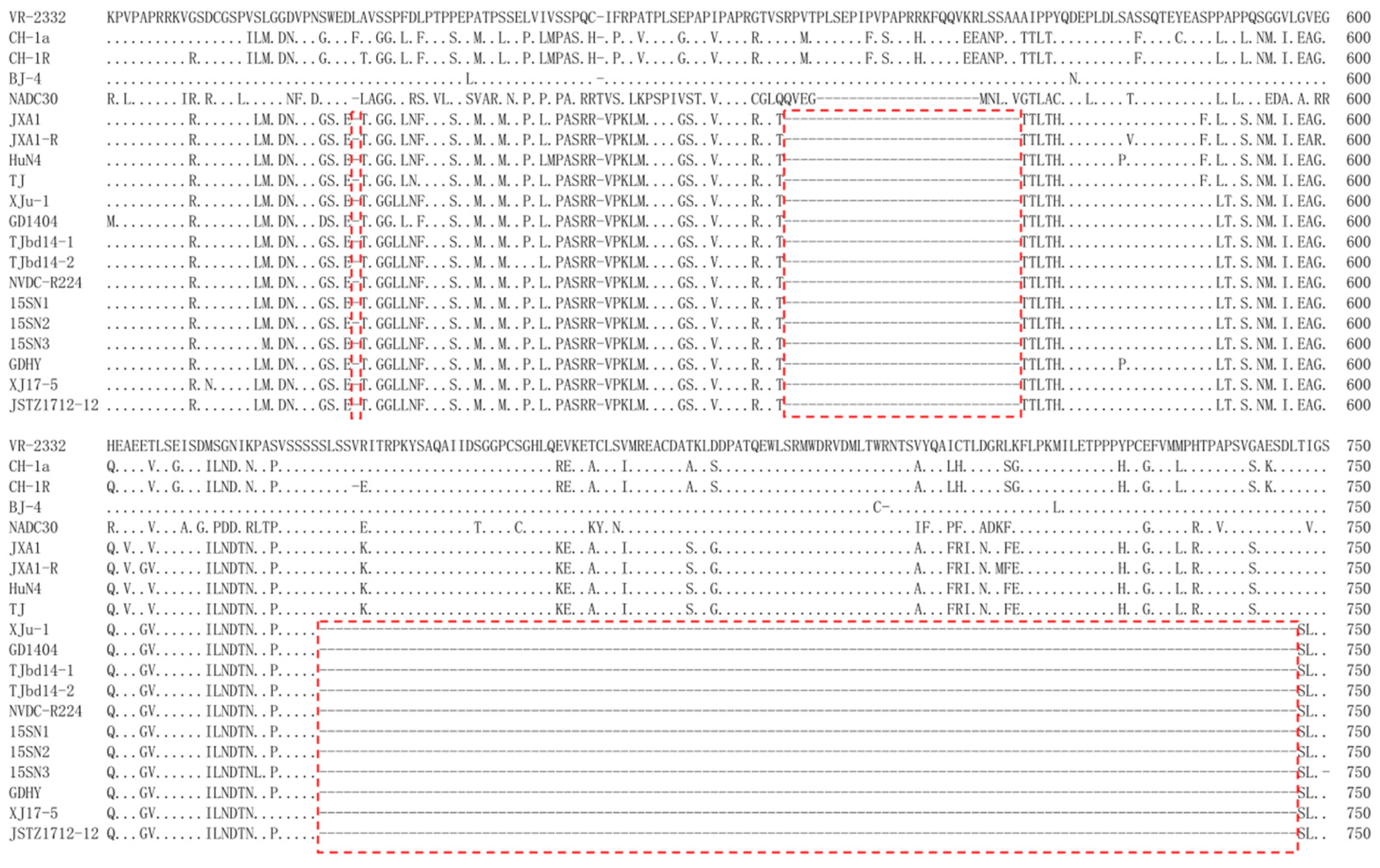
3.3. Genomic Comparison
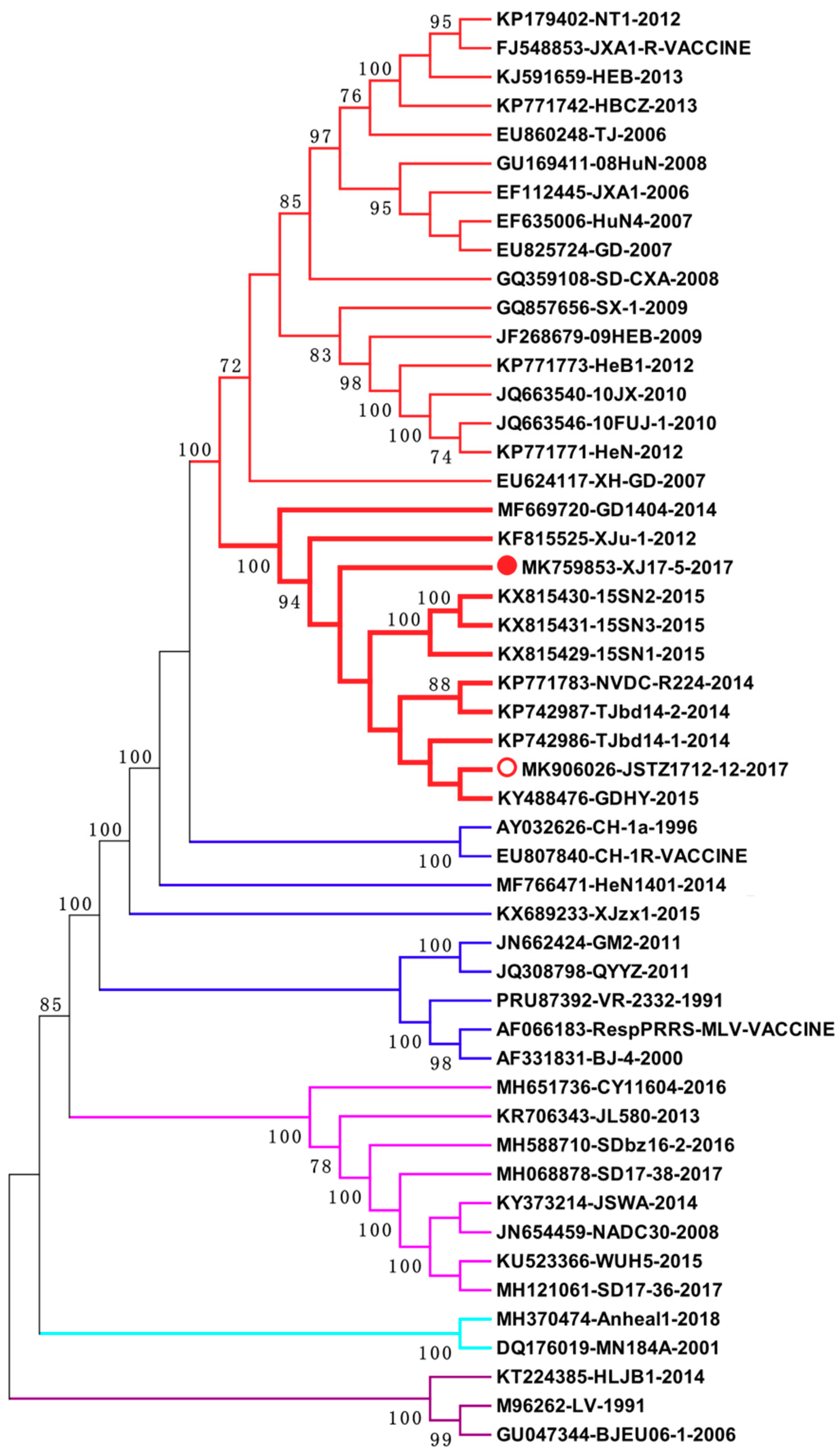
3.4. Phylogenetic Analysis
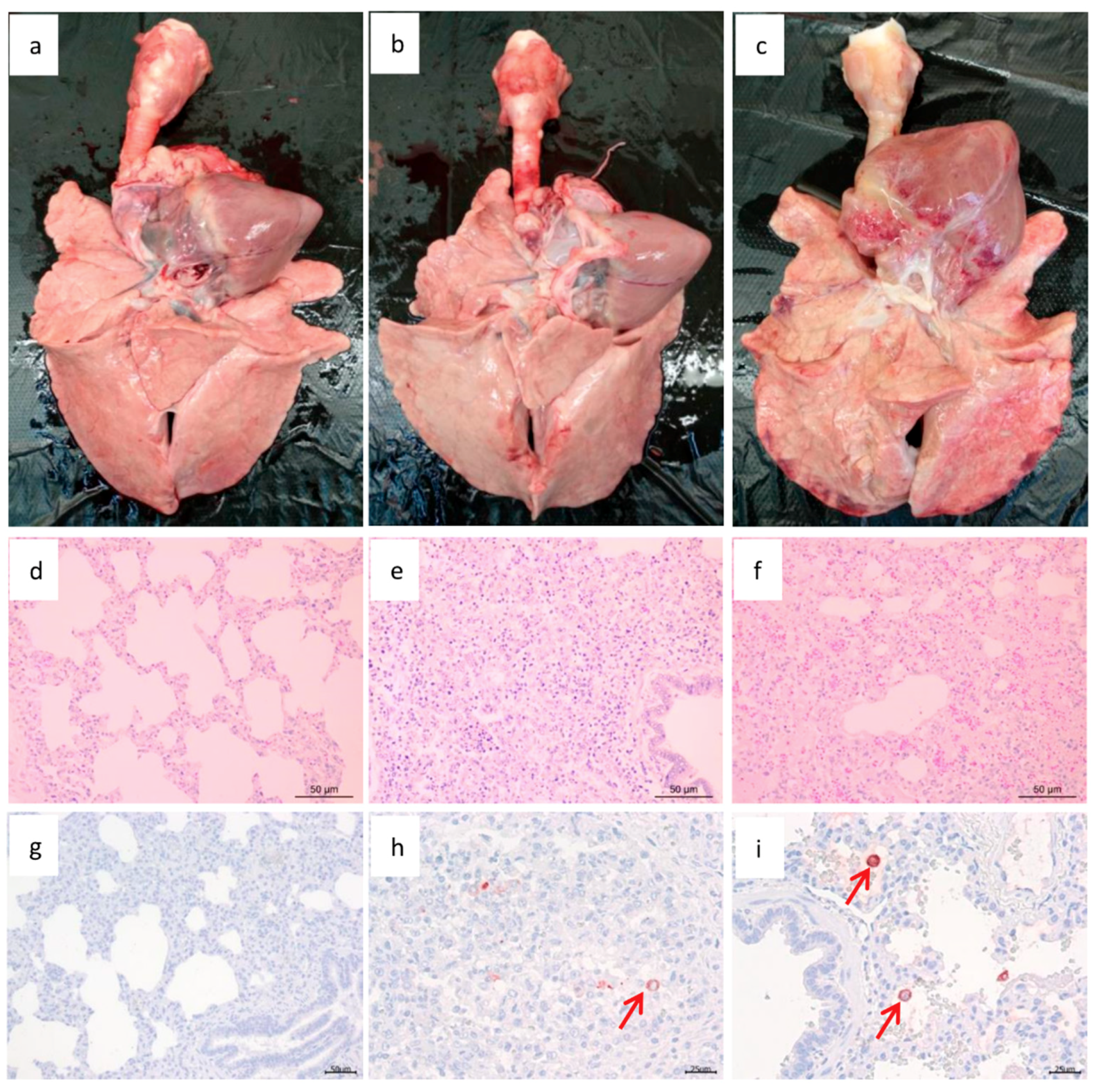
3.5. Pathogenic Analysis
3.6. Potential Virulence-Associated Amino Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Keffaber, K.K. Reprodutive failure of unknown etiology. Am. Assoc. Swine Vet. 1989, 1, 1–10. [Google Scholar]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Treffers, E.E.; Li, Y.; Tas, A.; Sun, Z.; van der Meer, Y.; de Ru, A.H.; van Veelen, P.A.; Atkins, J.F.; Snijder, E.J.; et al. Efficient -2 frameshifting by mammalian ribosomes to synthesize an additional arterivirus protein. Proc. Natl. Acad. Sci. USA 2012, 109, E2920–E2928. [Google Scholar] [CrossRef] [PubMed]
- Normile, D. Virology. China, Vietnam grapple with ‘rapidly evolving’ pig virus. Science 2007, 317, 1017. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, N.; Wang, L.; Wu, J.; Zhou, Z.; Ni, J.; Li, X.; Zhai, X.; Shi, J.; Tian, K. New genomic characteristics of highly pathogenic porcine reproductive and respiratory syndrome viruses do not lead to significant changes in pathogenicity. Vet. Microbiol. 2012, 158, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef]
- Chen, N.; Cao, Z.; Yu, X.; Deng, X.; Zhao, T.; Wang, L.; Liu, Q.; Li, X.; Tian, K. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J. Gen. Virol. 2011, 92, 880–892. [Google Scholar] [CrossRef]
- Chen, N.; Huang, Y.; Ye, M.; Li, S.; Xiao, Y.; Cui, B.; Zhu, J. Co-infection status of classical swine fever virus (CSFV), porcine reproductive and respiratory syndrome virus (PRRSV) and porcine circoviruses (PCV2 and PCV3) in eight regions of China from 2016 to 2018. Infect. Genet. Evol. 2019, 68, 127–135. [Google Scholar] [CrossRef]
- Guo, B.; Chen, Z.; Liu, W.; Cui, Y. Isolation and identification of porcine reproductory and respiratory syndrome (PRRS) virus from aborted fetuses suspected of PRRS. Chin. J. Prev. Vet. Med. 1996, 2, 1–5. [Google Scholar]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yang, H. Infection of porcine reproductive and respiratory syndrome virus suppresses the antibody response to classical swine fever virus vaccination. Vet. Microbiol. 2003, 95, 295–301. [Google Scholar] [CrossRef]
- Tong, G.Z.; Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; An, T.Q.; Qiu, H.J. Highly pathogenic porcine reproductive and respiratory syndrome, China. Emerg. Infect. Dis. 2007, 13, 1434–1436. [Google Scholar] [CrossRef] [PubMed]
- Leng, X.; Li, Z.; Xia, M.; Li, X.; Wang, F.; Wang, W.; Zhang, X.; Wu, H. Mutations in the genome of the highly pathogenic porcine reproductive and respiratory syndrome virus potentially related to attenuation. Vet. Microbiol. 2012, 157, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, J.; Zeng, J.; Yin, S.; Li, Y.; Zheng, L.; Guo, X.; Ge, X.; Yang, H. The 30-amino-acid deletion in the Nsp2 of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China is not related to its virulence. J. Virol. 2009, 83, 5156–5167. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, J.; Bai, X.; Ji, G.; Yan, H.; Li, Y.; Wang, Y.; Tan, F.; Xiao, Y.; Li, X.; et al. Pathogenicity comparison between highly pathogenic and NADC30-like porcine reproductive and respiratory syndrome virus. Arch. Virol. 2016, 161, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. 2015, 21, 2256–2257. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Loving, C.L.; Vorwald, A.C.; Kehrli, M.E., Jr.; Baker, R.B.; Nicholson, T.L.; Lager, K.M.; Miller, L.C.; Faaberg, K.S. Genomic sequence and virulence comparison of four Type 2 porcine reproductive and respiratory syndrome virus strains. Virus Res. 2012, 169, 212–221. [Google Scholar] [CrossRef]
- Chen, N.; Li, S.; Zhou, R.; Zhu, M.; He, S.; Ye, M.; Huang, Y.; Li, S.; Zhu, C.; Xia, P.; et al. Two novel porcine epidemic diarrhea virus (PEDV) recombinants from a natural recombinant and distinct subtypes of PEDV variants. Virus Res. 2017, 242, 90–95. [Google Scholar] [CrossRef]
- Chen, N.; Ye, M.; Xiao, Y.; Li, S.; Huang, Y.; Li, X.; Tian, K.; Zhu, J. Development of universal and quadruplex real-time RT-PCR assays for simultaneous detection and differentiation of porcine reproductive and respiratory syndrome viruses. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.H.; Chen, X.Z.; Hu, D.M.; Yu, X.L.; Wang, L.L.; Han, W.; Wu, J.J.; Cao, Z.; Wang, C.B.; Zhang, Q.; et al. Rapid differential detection of classical and highly pathogenic North American Porcine Reproductive and Respiratory Syndrome virus in China by a duplex real-time RT-PCR. J. Virol. Methods 2009, 161, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Ye, M.; Li, S.; Huang, Y.; Zhou, R.; Yu, X.; Tian, K.; Zhu, J. Emergence of a novel highly pathogenic recombinant virus from three lineages of porcine reproductive and respiratory syndrome virus 2 in China 2017. Transbound. Emerg. Dis. 2018, 65, 1775–1785. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.X.; Wang, X.; Yu, H.; Jiang, Y.F.; Gao, F.; Tong, W.; Li, L.W.; Li, H.C.; Yang, S.; Chen, P.F.; et al. The emergence of a highly pathogenic porcine reproductive and respiratory syndrome virus with additional 120aa deletion in Nsp2 region in Jiangxi, China. Transbound. Emerg. Dis. 2018, 65, 1740–1748. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Liu, Q.; Qiao, M.; Deng, X.; Chen, X.; Sun, M. Whole genome characterization of a novel porcine reproductive and respiratory syndrome virus 1 isolate: Genetic evidence for recombination between Amervac vaccine and circulating strains in mainland China. Infect. Genet. Evol. 2017, 54, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [Green Version]
- Kroll, J.; Piontkowski, M.; Kraft, C.; Coll, T.; Gomez-Duran, O. Initial vaccination and revaccination with Type I PRRS 94881 MLV reduces viral load and infection with porcine reproductive and respiratory syndrome virus. Porc. Health Manag. 2018, 4, 23. [Google Scholar] [CrossRef]
- Ooms, K.; van Gorp, H.; Botti, S.; van Gaever, T.; Delputte, P.L.; Nauwynck, H.J. Evaluation of viral peptide targeting to porcine sialoadhesin using a porcine reproductive and respiratory syndrome virus vaccination-challenge model. Virus Res. 2013, 177, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Holmes, E.C.; Brar, M.S.; Leung, F.C. Recombination is associated with an outbreak of novel highly pathogenic porcine reproductive and respiratory syndrome viruses in China. J. Virol. 2013, 87, 10904–10907. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Xu, X.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; Tian, K. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine 2016, 34, 5540–5545. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy evaluation of three modified-live virus vaccines against a strain of porcine reproductive and respiratory syndrome virus NADC30-like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, Y.; Ransburgh, R.; Snijder, E.J.; Fang, Y. Nonstructural protein 2 of porcine reproductive and respiratory syndrome virus inhibits the antiviral function of interferon-stimulated gene 15. J. Virol. 2012, 86, 3839–3850. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kaiser, T.J.; Horlen, K.; Keith, M.L.; Taylor, L.P.; Jolie, R.; Calvert, J.G.; Rowland, R.R. Insertion and deletion in a non-essential region of the nonstructural protein 2 (nsp2) of porcine reproductive and respiratory syndrome (PRRS) virus: Effects on virulence and immunogenicity. Virus Genes 2009, 38, 118–128. [Google Scholar] [CrossRef]
- Yoshii, M.; Okinaga, T.; Miyazaki, A.; Kato, K.; Ikeda, H.; Tsunemitsu, H. Genetic polymorphism of the nsp2 gene in North American type--porcine reproductive and respiratory syndrome virus. Arch. Virol. 2008, 153, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Chen, S.; Huang, J.; Ma, W.; Du, W.; Yang, X.; Liu, Y.; Yan, G. Complete genomic characterization of a porcine reproductive and respiratory syndrome virus isolate in Xinjiang province of China. Virus Genes 2015, 50, 39–45. [Google Scholar] [CrossRef]
- Yu, L.; Zhao, P.; Dong, J.; Liu, Y.; Zhang, L.; Liang, P.; Wang, L.; Song, C. Genetic characterization of 11 porcine reproductive and respiratory syndrome virus isolates in South China from 2014 to 2015. Virol. J. 2017, 14, 139. [Google Scholar] [CrossRef]
- Zhao, H.; Han, Q.; Zhang, L.; Zhang, Z.; Wu, Y.; Shen, H.; Jiang, P. Emergence of mosaic recombinant strains potentially associated with vaccine JXA1-R and predominant circulating strains of porcine reproductive and respiratory syndrome virus in different provinces of China. Virol. J. 2017, 14, 67. [Google Scholar] [CrossRef]
- Li, B.; Fang, L.; Xu, Z.; Liu, S.; Gao, J.; Jiang, Y.; Chen, H.; Xiao, S. Recombination in vaccine and circulating strains of porcine reproductive and respiratory syndrome viruses. Emerg. Infect. Dis. 2009, 15, 2032–2035. [Google Scholar] [CrossRef] [PubMed]
- Ming, S.; Yongying, M.; Bohua, L.; Huiying, L.; Xiaoyu, D.; Qiaorong, L.; Mingming, Q.; Xi, C.; Xinyan, Y.; Xizhao, C. Pathogenic Characterization of European Genotype Porcine Reproductive and Respiratory Syndrome Virus Recently Isolated in Mainland China. Open Virol. J. 2017, 11, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Xia, M.; Wang, W.; Ju, D.; Cao, L.; Wu, B.; Wang, X.; Wu, Y.; Song, N.; Hu, J.; et al. An Attenuated Highly Pathogenic Chinese PRRS Viral Vaccine Confers Cross Protection to Pigs against Challenge with the Emerging PRRSV NADC30-Like Strain. Virol. Sin. 2018, 33, 153–161. [Google Scholar] [CrossRef]
- Han, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Pathogenesis and control of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Allende, R.; Kutish, G.F.; Laegreid, W.; Lu, Z.; Lewis, T.L.; Rock, D.L.; Friesen, J.; Galeota, J.A.; Doster, A.R.; Osorio, F.A. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch. Virol. 2000, 145, 1149–1161. [Google Scholar] [CrossRef]
- Kim, W.I.; Kim, J.J.; Cha, S.H.; Yoon, K.J. Different biological characteristics of wild-type porcine reproductive and respiratory syndrome viruses and vaccine viruses and identification of the corresponding genetic determinants. J. Clin. Microbiol. 2008, 46, 1758–1768. [Google Scholar] [CrossRef]
- Han, W.; Wu, J.J.; Deng, X.Y.; Cao, Z.; Yu, X.L.; Wang, C.B.; Zhao, T.Z.; Chen, N.H.; Hu, H.H.; Bin, W.; et al. Molecular mutations associated with the in vitro passage of virulent porcine reproductive and respiratory syndrome virus. Virus Genes 2009, 38, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Tang, Y.D.; Liu, C.X.; Xiang, L.R.; Zhang, W.L.; Leng, C.L.; Wang, Q.; An, T.Q.; Peng, J.M.; Tian, Z.J.; et al. Adaptions of field PRRSVs in Marc-145 cells were determined by variations in the minor envelope proteins GP2a-GP3. Vet. Microbiol. 2018, 222, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Trus, I.; Oh, D.; Kvisgaard, L.K.; Rappe, J.C.F.; Ruggli, N.; Vanderheijden, N.; Larsen, L.E.; Lefevre, F.; Nauwynck, H.J. A Triple Amino Acid Substitution at Position 88/94/95 in Glycoprotein GP2a of Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV1) Is Responsible for Adaptation to MARC-145 Cells. Viruses 2019, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B.; Ansari, I.H.; Pattnaik, A.K.; Osorio, F.A. Identification of virulence determinants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones. Virology 2008, 380, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, L.; Zhang, J.; Ge, X.; Zhou, R.; Zheng, H.; Geng, G.; Guo, X.; Yang, H. Nsp9 and nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in china. PLoS Pathog. 2014, 10, e1004216. [Google Scholar] [CrossRef] [PubMed]
- Osorio, F.A.; Galeota, J.A.; Nelson, E.; Brodersen, B.; Doster, A.; Wills, R.; Zuckermann, F.; Laegreid, W.W. Passive transfer of virus-specific antibodies confers protection against reproductive failure induced by a virulent strain of porcine reproductive and respiratory syndrome virus and establishes sterilizing immunity. Virology 2002, 302, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Lager, K.M.; Mengeling, W.L.; Wesley, R.D. Strain predominance following exposure of vaccinated and naive pregnant gilts to multiple strains of porcine reproductive and respiratory syndrome virus. Can. J. Vet. Res. 2003, 67, 121–127. [Google Scholar] [PubMed]
- Mengeling, W.L.; Lager, K.M.; Vorwald, A.C.; Koehler, K.J. Strain specificity of the immune response of pigs following vaccination with various strains of porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2003, 93, 13–24. [Google Scholar] [CrossRef]
- Xu, L.; Zhou, L.; Sun, W.; Zhang, P.; Ge, X.; Guo, X.; Han, J.; Yang, H. Nonstructural protein 9 residues 586 and 592 are critical sites in determining the replication efficiency and fatal virulence of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Virology 2018, 517, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Gao, J.C.; Xiong, J.Y.; Guo, J.C.; Yang, Y.B.; Jiang, C.G.; Tang, Y.D.; Tian, Z.J.; Cai, X.H.; Tong, G.Z.; et al. Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| XJ17-5/JSTZ1712-12 * | XJ17-5/JSTZ1712-12 # | VR-2332 | NADC30 | TJ | LV |
|---|---|---|---|---|---|
| Nucleotide Identity (%) to XJ17-5/JSTZ1712-12 | |||||
| Complete (14960) | 99.45 | 86.27/86.45 | 82.13/81.93 | 96.46/96.84 | 58.31/58.31 |
| 5’UTR (1–189) | 99.47 | 91.01/91.53 | 90.58/91.10 | 96.30/96.83 | 51.80/53.15 |
| ORF1a (190–7251) | 99.35 | 82.00/82.31 | 72.55/72.65 | 93.82/94.25 | 53.30/53.31 |
| Nsp1α (190–729) | 99.63 | 92.04/92.04 | 89.81/90.00 | 99.26/99.63 | 62.11/61.92 |
| Nsp1β (730–1338) | 98.36 | 86.21/86.54 | 80.62/80.79 | 98.03/99.01 | 47.76/47.92 |
| Nsp2 (1339–4479) | 99.33 | 73.88/74.21 | 67.26/67.30 | 88.17/88.60 | 49.44/49.39 |
| Nsp2N (1339–3438) | 99.14 | 66.31/66.75 | 58.31/58.54 | 83.41/83.98 | 41.27/41.16 |
| Nsp2TF (1339–3438, 3441–3945) | 99.31 | 70.92/71.82 | 63.97/64.22 | 86.11/86.59 | 47.37/47.17 |
| Nsp3 (4477–5166) | 98.99 | 88.55/89.28 | 82.03/82.46 | 97.83/98.55 | 56.23/56.52 |
| Nsp4 (5167–5778) | 99.67 | 89.87/89.87 | 85.29/84.97 | 99.67/99.67 | 60.62/60.95 |
| Nsp5 (5779–6288) | 99.61 | 89.02/89.41 | 90.20/90.59 | 99.02/99.41 | 63.14/63.33 |
| Nsp6 (6289–6336) | 100 | 93.75/93.75 | 91.67/91.67 | 97.92/97.92 | 70.83/70.83 |
| Nsp7α (6337–6783) | 99.55 | 90.38/90.83 | 84.56/84.56 | 98.88/99.33 | 55.36/55.58 |
| Nsp7β (6784–7113) | 100 | 87.88/87.88 | 79.39/79.39 | 99.39/99.39 | 48.66/48.66 |
| Nsp8 (7114–7248) | 100 | 96.30/96.30 | 89.63/89.63 | 100/100 | 64.44/64.44 |
| ORF1b (7248–11621) | 99.47 | 91.08/91.06 | 87.93/87.86 | 98.88/99.22 | 62.11/62.14 |
| Nsp9 (7248–9167) | 99.32 | 92.45/92.29 | 87.60/87.34 | 98.54/98.91 | 66.35/66.35 |
| Nsp10 (9168–10487) | 99.39 | 89.85/90.00 | 85.76/85.91 | 98.79/99.24 | 60.30/60.30 |
| Nsp11 (10488–11159) | 99.70 | 90.18/90.18 | 91.22/91.22 | 99.26/99.55 | 65.33/65.63 |
| Nsp12 (11160–11618) | 100 | 89.39/89.39 | 89.83/89.83 | 99.13/99.13 | 46.15/46.27 |
| ORF2a (11623–12393) | 99.74 | 93.13/93.39 | 86.64/86.38 | 99.35/99.61 | 62.13/63.55 |
| ORF2b (11628–11849) | 100 | 93.24/93.24 | 87.67/87.67 | 100/100 | 68.72/68.72 |
| ORF3 (12246–13010) | 99.74 | 89.15/89.41 | 83.14/83.40 | 98.95/99.22 | 61.92/62.17 |
| ORF4 (12791–13327) | 99.63 | 89.76/90.13 | 86.78/87.15 | 99.07/99.44 | 64.49/64.67 |
| ORF5 (13338–13940) | 99.83 | 89.05/88.89 | 86.24/86.07 | 99.17/99.34 | 61.33/61.17 |
| ORF5a (13328–13468) | 100 | 88.65/88.65 | 85.82/85.82 | 98.58/98.58 | 55.32/55.32 |
| ORF6 (13925–14449) | 99.62 | 95.43/95.05 | 89.33/88.95 | 99.43/99.81 | 69.14/69.14 |
| ORF7 (14439–14810) | 98.92 | 93.28/93.55 | 90.32/90.59 | 98.92/99.46 | 62.37/60.86 |
| 3’UTR (14811–14960) | 98.67 | 92.05/93.38 | 88.08/89.40 | 98.67/100 | 55.63/54.30 |
| No. | Protein | Position * | XJ17-5 | JSTZ1712-12 | XJu-1 | GD1404 | NVDC-R224 | TJbd14-1 | TJbd14-2 | GDHY | 15SN1 | 15SN2 | 15SN3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Nsp1α | 114 | A# | T | T | T | T | T | T | T | T | T | T |
| 2 | Nsp1β | 8 | C | R | R | R | R | R | R | R | R | R | R |
| 3 | 83 | N | D | D | D | D | D | D | D | D | D | D | |
| 4 | 107 | I | F | I | I | I | I | I | I | I | I | I | |
| 5 | 122 | A | P | P | A | S | S | S | P | P | P | P | |
| 6 | 137 | A | T | T | A | T | A | A | T | A | A | A | |
| 7 | Nsp2 | 26 | V | I | I | T | I | I | I | I | I | I | I |
| 8 | 147 | E | G | E | E | E | E | E | E | E | E | E | |
| 9 | 335 | F | S | S | S | S | S | S | S | F | F | F | |
| 10 | 355 | V | A | A | V | V | V | V | A | V | V | V | |
| 11 | 363 | I | V | V | V | V | V | V | V | V | V | V | |
| 12 | 393 | K | E | E | E | E | E | E | E | E | E | G | |
| 13 | 463 | N | D | D | D | D | D | D | D | D | D | D | |
| 14 | 590 | S | P | P | P | P | P | P | P | P | P | P | |
| 15 | 604 | G | D | D | D | D | D | D | D | D | D | - | |
| 16 | 640 | S | P | P | P | P | P | P | P | P | P | P | |
| 17 | 999 | E | D | E | E | E | D | E | E | E | E | E | |
| 18 | Nsp3 | 32 | T | A | A | A | A | A | A | A | A | A | A |
| 19 | 102 | T | A | A | A | A | A | A | A | A | A | A | |
| 20 | Nsp4 | 184 | K | N | K | N | N | N | N | N | N | N | N |
| 21 | Nsp5 | 53 | A | V | V | V | V | V | V | V | V | V | V |
| 22 | Nsp7α | 8 | I | M | M | M | M | M | M | M | M | M | M |
| 23 | Nsp9 | 478 | G | A | A | A | A | A | A | A | A | A | A |
| 24 | 502 | I | T | T | I | I | I | I | I | I | I | I | |
| 25 | Nsp10 | 109 | K | R | R | R | R | R | R | R | R | R | R |
| 26 | 191 | I | V | V | V | V | V | V | V | V | V | V | |
| 27 | 297 | S | A | A | S | S | A | S | A | S | S | S | |
| 28 | 316 | A | V | V | V | V | V | V | V | V | V | V | |
| 29 | GP3 | 69 | S | P | P | P | P | P | P | P | P | P | P |
| 30 | 228 | P | S | S | S | S | S | S | S | S | S | S | |
| 31 | GP4 | 46 | A | V | V | V | V | V | V | V | V | V | V |
| 32 | GP5 | 200 | P | L | L | L | P | L | P | L | L | L | L |
| 33 | N | 51 | E | G | G | E | G | G | G | G | G | G | E |
| 34 | 123 | V | A | A | A | A | A | A | A | A | A | A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, N.; Ye, M.; Huang, Y.; Li, S.; Xiao, Y.; Li, X.; Li, S.; Li, X.; Yu, X.; Tian, K.; et al. Identification of Two Porcine Reproductive and Respiratory Syndrome Virus Variants Sharing High Genomic Homology but with Distinct Virulence. Viruses 2019, 11, 875. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090875
Chen N, Ye M, Huang Y, Li S, Xiao Y, Li X, Li S, Li X, Yu X, Tian K, et al. Identification of Two Porcine Reproductive and Respiratory Syndrome Virus Variants Sharing High Genomic Homology but with Distinct Virulence. Viruses. 2019; 11(9):875. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090875
Chicago/Turabian StyleChen, Nanhua, Mengxue Ye, Yucheng Huang, Shuai Li, Yanzhao Xiao, Xinshuai Li, Shubin Li, Xiangdong Li, Xiuling Yu, Kegong Tian, and et al. 2019. "Identification of Two Porcine Reproductive and Respiratory Syndrome Virus Variants Sharing High Genomic Homology but with Distinct Virulence" Viruses 11, no. 9: 875. https://0-doi-org.brum.beds.ac.uk/10.3390/v11090875