Functional Transcomplementation between Wheat Dwarf Virus Strains in Wheat and Barley

 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
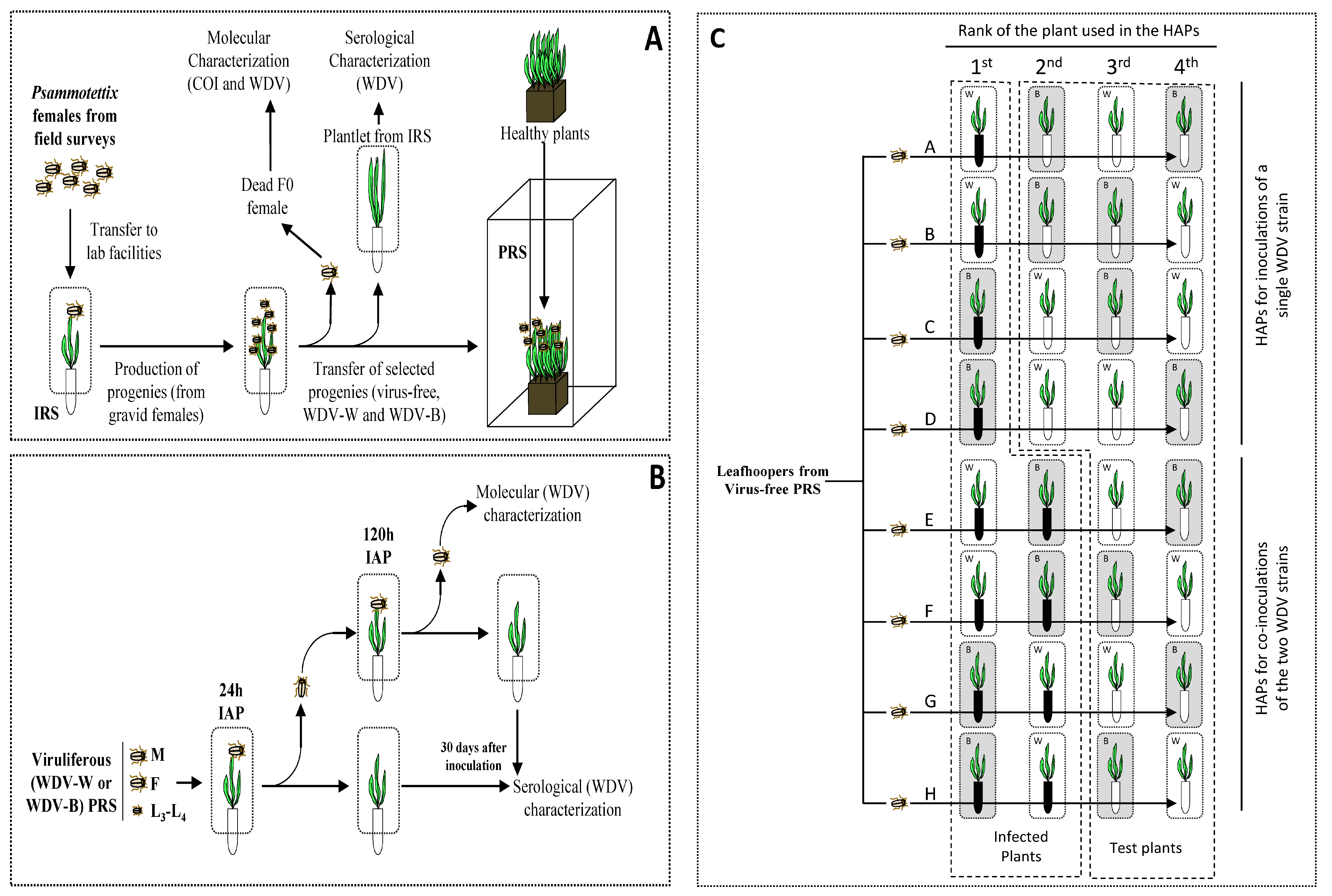
2.1. Plants, Insects and Viruses
2.2. DNA Extraction from Samples
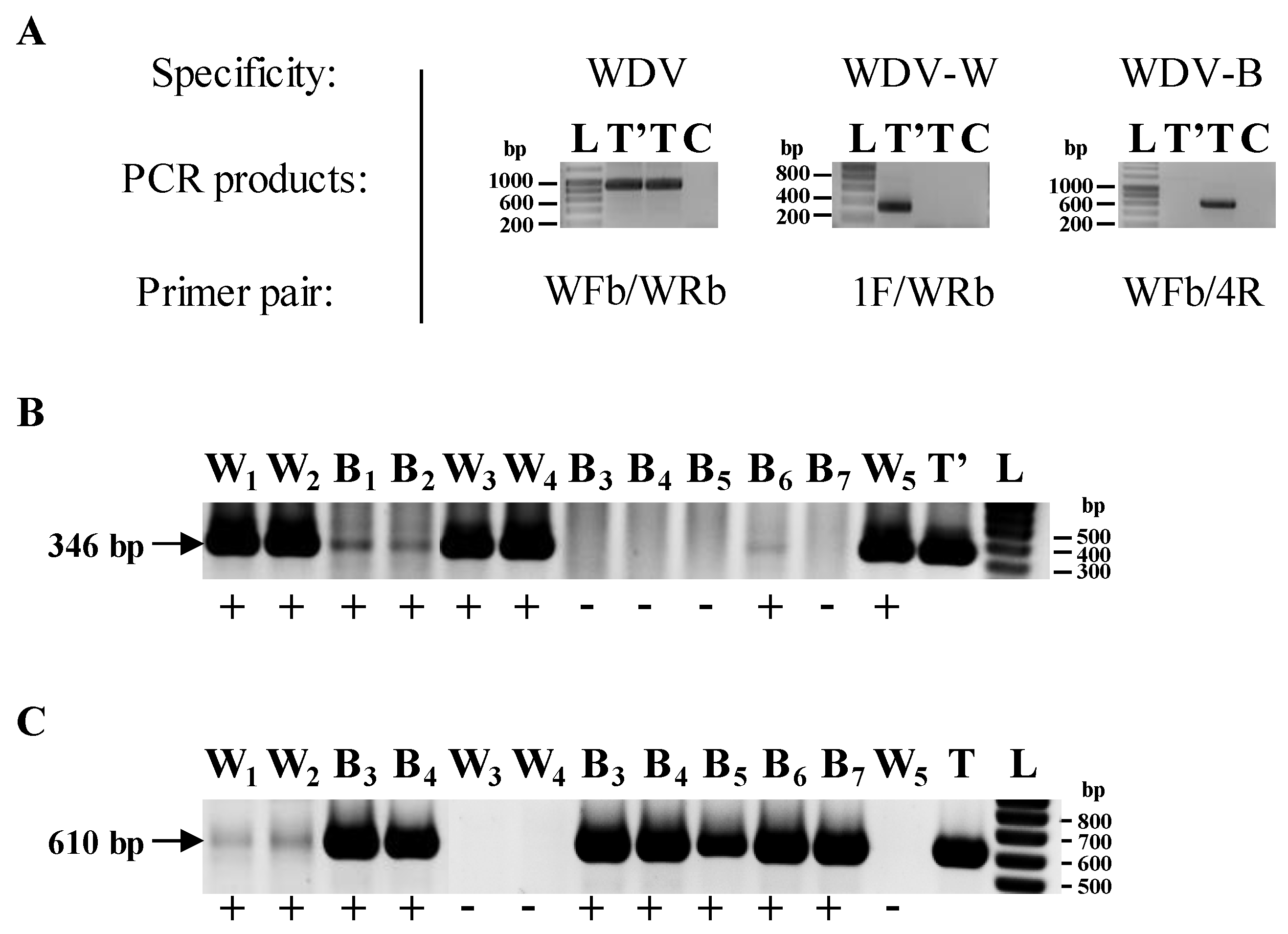
2.3. Detection of WDV and Sequencing of WDV Isolates
2.4. Characterization of Psammotettix Species Reared in PRS
2.5. Acquisition and Inoculation of WDV by P. alienus
2.6. Serological Detection of WDV in Plants
2.7. Data Analyses
3. Results
3.1. Characterization of the Leafhoppers and WDV Isolates Used in the Experiments
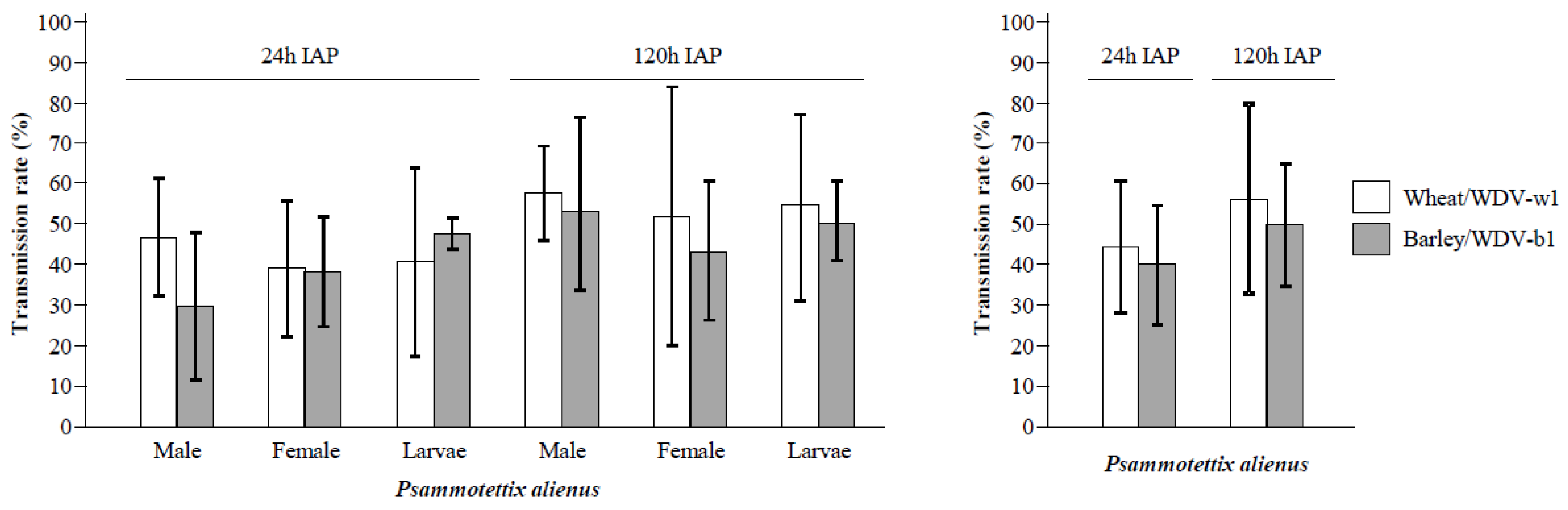
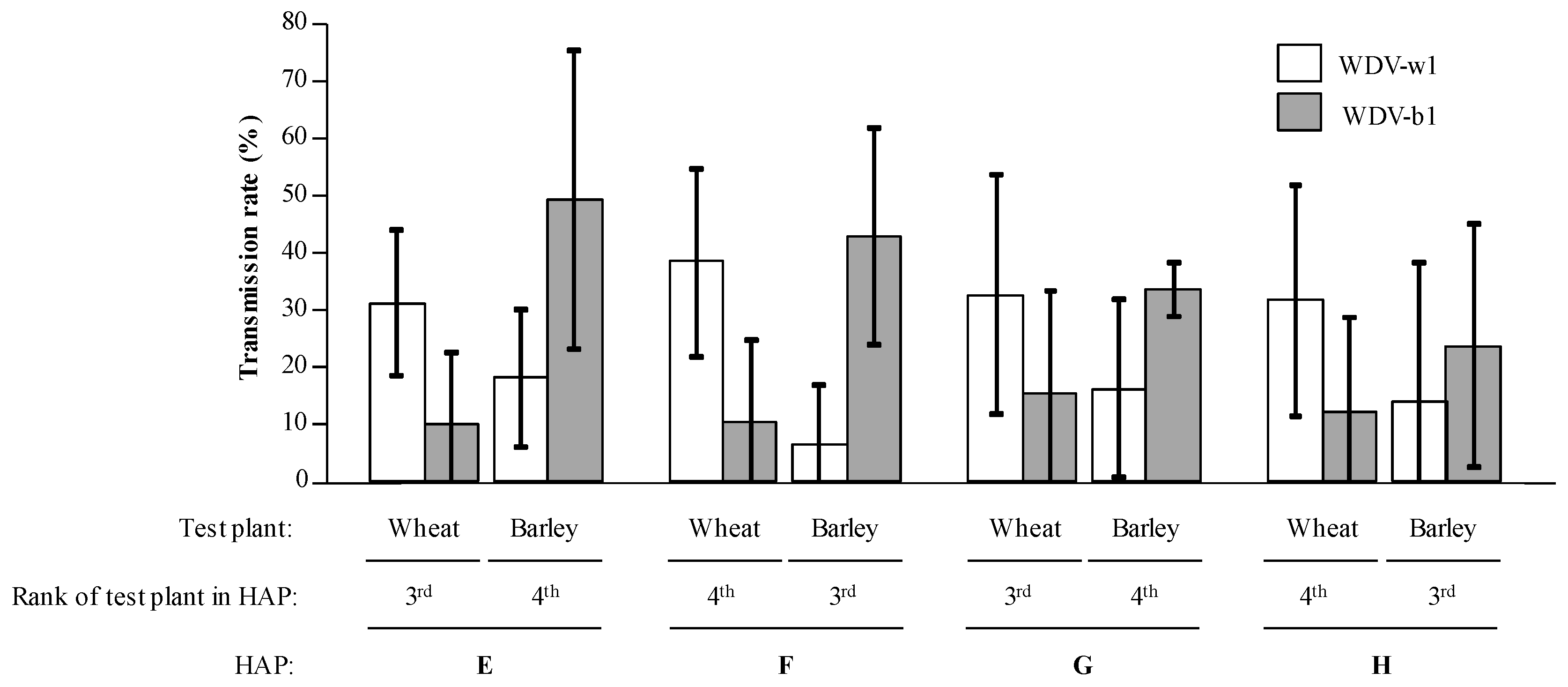
3.2. Transmission of WDV by P. alienus
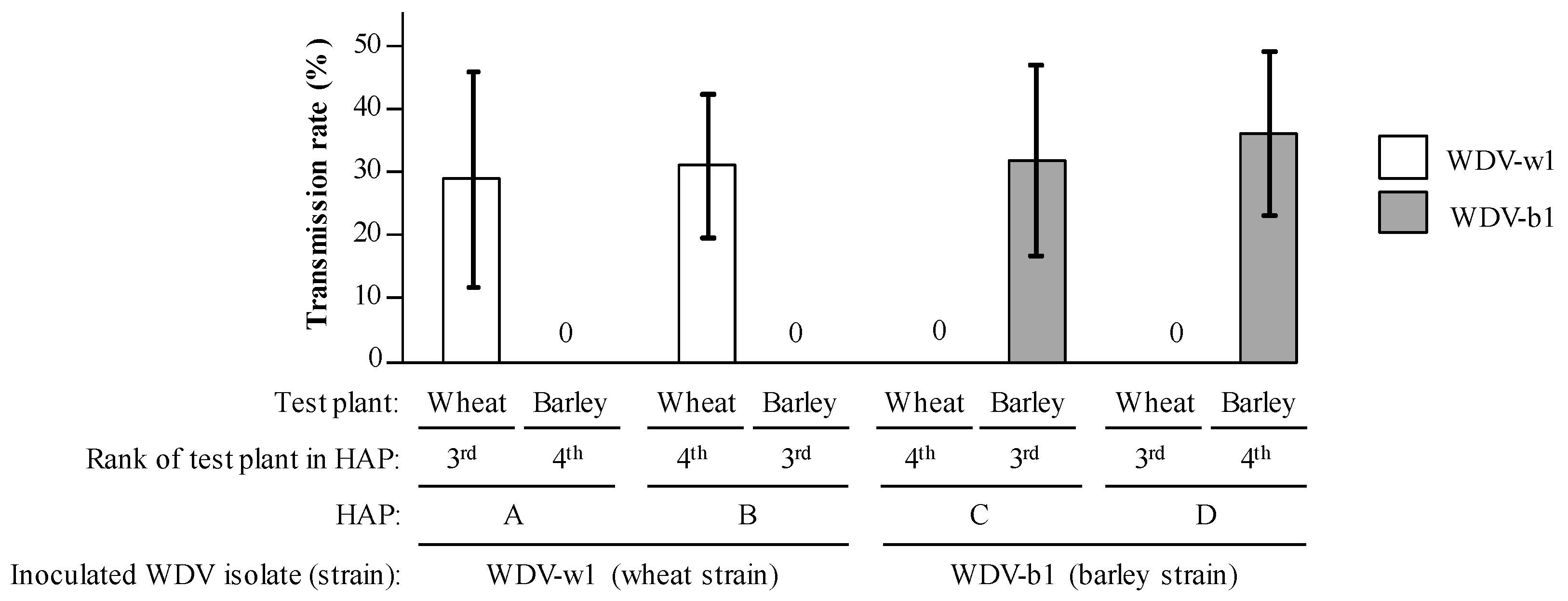
3.3. Inoculation of WDV-w1 and WDV-b1 to Barley and Wheat Plantlets
3.4. Transmission Efficiencies in Multiple Acquisition/Inoculation Procedures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Vacke, J. Wheat dwarf virus disease. Biol. Plant. 1961, 3, 228–233. [Google Scholar] [CrossRef]
- Vacke, J. Host plants range and symptoms of wheat dwarf virus. Vyzkumnych Ustavu Rostlinné Vyroby Praha Ruzyne 1972, 17, 151–162. [Google Scholar]
- Lindsten, K.; Lindsten, B. Wheat dwarf-an old disease with new outbreaks in Sweden. J. Plant Dis. Protect. 1999, 106, 325–332. [Google Scholar]
- Lindblad, M.; Waern, P. Correlation of wheat dwarf incidence to winter wheat cultivation practices. Agric. Ecosyst. Environ. 2002, 92, 115–122. [Google Scholar] [CrossRef]
- Sirlova, L.; Vacke, J.; Chaloupkova, M. Reaction of selected winter wheat varieties to autumnal infection with Wheat dwarf virus. Plant Protect. Sci. 2005, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Macdowell, S.W.; Macdonald, H.; Hamilton, W.D.O.; Coutts, R.H.A.; Buck, K.W. The nucleotide sequence of cloned Wheat dwarf virus DNA. EMBO J. 1985, 4, 2173–2180. [Google Scholar] [CrossRef]
- Hofer, J.M.I.; Dekker, E.L.; Reynolds, H.V.; Woolston, C.J.; Cox, B.S.; Mullineaux, P.M. Coordinate regulation of replication and virion sense gene expression in wheat dwarf virus. Plant Cell 1992, 4, 213–223. [Google Scholar]
- Lindsten, K.; Vacke, J. A possible barley adapted strain of wheat dwarf virus (WDV). Acta Phytopathol. Entomol. Hung. 1991, 26, 175–180. [Google Scholar]
- Commandeur, U.; Huth, W. Differentiation of strains of wheat dwarf virus in infected wheat and barley plants by means of polymerase chain reaction. J. Plant Dis. Protect. 1999, 106, 550–552. [Google Scholar]
- Muhire, B.; Martin, D.P.; Brown, J.K.; Navas-Castillo, J.; Moriones, E.; Zerbini, F.M.; Rivera-Bustamante, R.; Malathi, V.G.; Briddon, R.W.; Varsani, A. A genome-wide pairwise-identity-based proposal for the classification of viruses in the genus Mastrevirus (family Geminiviridae). Arch. Virol. 2013, 158, 1411–1424. [Google Scholar] [CrossRef] [Green Version]
- Schubert, J.; Habekuss, A.; Wu, B.L.; Thieme, T.; Wang, X.F. Analysis of complete genomes of isolates of the Wheat dwarf virus from new geographical locations and descriptions of their defective forms. Virus Genes 2014, 48, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Koklu, G.; Ramsell, J.N.E.; Kvarnheden, A. The complete genome sequence for a Turkish isolate of Wheat dwarf virus (WDV) from barley confirms the presence of two distinct WDV strains. Virus Genes 2007, 34, 359–366. [Google Scholar] [CrossRef]
- Schubert, J.; Habekuss, A.; Kazmaier, K.; Jeske, H. Surveying cereal-infecting geminiviruses in Germany–Diagnostics and direct sequencing using rolling circle amplification. Virus Res. 2007, 127, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Kvarnheden, A.; Lindblad, M.; Lindsten, K.; Valkonen, J.P.T. Genetic diversity of Wheat dwarf virus. Arch. Virol. 2002, 147, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Ramsell, J.N.E.; Lemmetty, A.; Jonasson, J.; Andersson, A.; Sigvald, R.; Kvarnheden, A. Sequence analyses of Wheat dwarf virus isolates from different hosts reveal low genetic diversity within the wheat strain. Plant Pathol. 2008, 57, 834–841. [Google Scholar] [CrossRef]
- Kundu, J.; Gadiou, S.; Cervena, G. Discrimination and genetic diversity of Wheat dwarf virus in the Czech Republic. Virus Genes 2009, 38, 468–474. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, B.; Vida, G.; Cseplo-Karolyi, M.; Wu, B.L.; Wu, Y.H.; Wang, X.F. Genomic Analysis of the Natural Population of Wheat dwarf virus in Wheat from China and Hungary. J. Integr. Agric. 2012, 11, 2020–2027. [Google Scholar] [CrossRef]
- Brault, V.; Uzest, M.; Monsion, B.; Jacquot, E.; Blanc, S. Aphids as transport devices for plant viruses. Comptes Rendus Biol. 2010, 333, 524–538. [Google Scholar] [CrossRef]
- Raatikainen, M.; Vasarainen, A. Early-and high-summer flight periods of leafhoppers. Ann. Agric. Fenn. 1973, 12, 77–94. [Google Scholar]
- Lindblad, M.; Areno, P. Temporal and spatial population dynamics of Psammotettix alienus, a vector of wheat dwarf virus. Int. J. Pest Manag. 2002, 48, 233–238. [Google Scholar] [CrossRef]
- Vilbaste, J. Preliminary key for the identification of the nymphs of North European Homoptera, Cicadinea. II. Cicadelloidea. Ann. Zool. Fenn. Vanamo 1982, 19, 1–20. [Google Scholar]
- Della Giustina, W. Homoptères Cicadellidae, Volume 3 Compléments; INRA, Ed.; Fédération Française des Sociétés de Sciences Naturelles: Faune, France, 1989; p. 350. [Google Scholar]
- Zhang, X.; Zhou, G.H.; Wang, X.F. Detection of wheat dwarf virus (WDV) in wheat and vector leafhopper (Psammotettix alienus Dahlb.) by real-time PCR. J. Virol. Methods 2010, 169, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Ekzayez, A.M.; Kumari, S.G.; Ismail, I. First Report of Wheat dwarf virus and Its Vector (Psammotettix provincialis) Affecting Wheat and Barley Crops in Syria. Plant Dis. 2011, 95, 76. [Google Scholar] [CrossRef]
- Abt, I.; Derlink, M.; Mabon, R.; Virant-Doberlet, M.; Jacquot, E. Integrating multiple criteria for the characterization of Psammotettix populations in European cereal fields. Bull. Entomol. Res. 2017, 108, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Derlink, M.; Abt, I.; Mabon, R.; Julian, C.; Virant-Doberlet, M.; Jacquot, E. Mating behavior of Psammotettix alienus (Hemiptera: Cicadellidae). Insect Sci. 2018, 25, 148–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manurung, B.; Witsack, W.; Mehner, S.; Gruntzig, M.; Fuchs, E. Studies on biology and population dynamics of the leafhopper Psammotettix alienus Dahlb. (Homoptera: Auchenorrhyncha) as vector of Wheat dwarf virus (WDV) in Saxony-Anhalt, Germany. J. Plant Dis. Protect. 2005, 112, 497–507. [Google Scholar]
- Guglielmino, A.; Virla, E.G. Postembryonic development and biology of Psammotettix alienus (Dahlbom) (Homoptera, ‘Cicadellidae) under laboratory conditions. Boll. Zool. 1997, 29, 65–80. [Google Scholar]
- Manurung, B.; Witsack, W.; Mehner, S.; Gruntzig, M.; Fuchs, E. The epidemiology of Wheat dwarf virus in relation to occurrence of the leafhopper Psammotettix alienus in Middle-Germany. Virus Res. 2004, 100, 109–113. [Google Scholar] [CrossRef]
- Abt, I.; Julian, C.; Souquet, M.; Derlink, M.; Mabon, R.; Angot, G.; Thébaud, G.; de Keyzer, A.; Letroublon, M.; Virant-Doberlet, M.; et al. Characterization of Psammotettix species, vectors of Wheat dwarf virus. Presented at 15. Rencontres de Virologie Végétale, Aussois, France, 18–22 January 2015. [Google Scholar]
- Vacke, J.; Cibulka, R. Silky bent grass (Apera spica-venti (L.) Beauv.)-A new host and reservoir of wheat dwarf virus. Plant Protect. Sci. 1999, 35, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, E.; Mehner, S.; Manurung, B.; Grüntzig, M. Strain-spectrum of Wheat dwarf virus (WDV) in Saxony-Anhalt. In Proceedings of the Fifth Congress of the European Foundation for Plant Pathology, Taormina-Giardini Naxos, Italy, 18–22 September 2000; pp. 213–215. [Google Scholar]
- Tobias, I.; Shevchenko, O.; Kiss, B.; Bysov, A.; Snihur, H.; Polischuk, V.; Salanki, K.; Palkovics, L. Comparison of the Nucleotide Sequences of Wheat Dwarf Virus (WDV) isolates from Hungary and Ukraine. Pol. J. Microbiol. 2011, 60, 125–131. [Google Scholar] [CrossRef]
- Ramsell, J.N.E.; Boulton, M.I.; Martin, D.P.; Valkonen, J.P.T.; Kvarheden, A. Studies on the host range of the barley strain of Wheat dwarf virus using an agroinfectious viral clone. Plant Pathol. 2009, 58, 1161–1169. [Google Scholar] [CrossRef]
- Wang, X.F.; Wu, B.; Wang, J.F. First report of Wheat dwarf virus infecting barley in Yunnan, China. J. Plant Pathol. 2008, 90, 400. [Google Scholar]
- Posadas, G.; Henry, M. Resistance to BYDV-PAV and CYDV-RPV in a bread wheat x agrotricum cross. In Barley Yellow Dwarf Disease: Recent Advances and Future Strategies; Henry, M., McNab, A., Eds.; CIMMYT: Texcoco, Mexico, 2002; pp. 127–129. [Google Scholar]
- Sadeghi, S.E.; Dedryver, C.A.; Riault, G.; Tanguy, S. Variation in virus content among individual leaves and roots of barley and wheat infected with a BYDV-PAV isolate. J. Agric. Sci. Technol. 2000, 2, 151–160. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.A.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control of mitochondrial-DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Clark, M.F.; Adams, A.N. Characteristics of microplate method of Enzyme-Linked ImmunoSorbent Assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Gadiou, S.; Ripl, J.; Janourova, B.; Jarosova, J.; Kundu, J.K. Real-time PCR assay for the discrimination and quantification of wheat and barley strains of Wheat dwarf virus. Virus Genes 2012, 44, 349–355. [Google Scholar] [CrossRef]
- Dedryver, C.A.; Riault, G.; Tanguy, S.; Le Gallic, J.F.; Trottet, M.; Jacquot, E. Intra specific variation and inheritance of BYDV-PAV transmission in the aphid Sitobion avenae. Eur. J. Plant Pathol. 2005, 111, 341–354. [Google Scholar] [CrossRef]
- Fox, A.; Collins, L.E.; Macarthur, R.; Blackburn, L.F.; Northing, P. New aphid vectors and efficiency of transmission of Potato virus A and strains of Potato virus Y in the UK. Plant. Pathol. 2017, 66, 325–335. [Google Scholar] [CrossRef]
- Bluemel, J.K.; King, A.R.; Virant-Doberet, M.; Symondson, W.O.C. Primers for identification of type and other archived specimens of Aphrodes leafhoppers (Hemiptera, Cicadellidae). Mol. Ecol. Resour. 2011, 11, 770–774. [Google Scholar] [CrossRef]
- Wang, Y.J.; Mao, Q.Z.; Liu, W.W.; Mar, T.; Wei, T.Y.; Liu, Y.; Wang, X.F. Localization and Distribution of Wheat dwarf virus in Its Vector Leafhopper, Psammotettix alienus. Phytopathology 2014, 104, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Latham, J.R.; Wilson, A.K. Transcomplementation and synergism in plants: Implications for viral transgenes? Mol. Plant Pathol. 2008, 9, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M.; Ryabov, E.; Fraser, G.; Taliansky, M. Mechanical transmission of Potato leafroll virus. J. Gen. Virol. 2000, 81, 2791–2795. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Fraser, G.; Mayo, M.A.; Barker, H.; Taliansky, M. Umbravirus gene expression helps potato leafroll virus to invade mesophyll tissues and to be transmitted mechanically between plants. Virology 2001, 286, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Spitsin, S.; Steplewski, K.; Fleysh, N.; Belanger, H.; Mikheeva, T.; Shivprasad, S.; Dawson, W.; Koprowski, H.; Yusibov, V. Expression of alfalfa mosaic virus coat protein in tobacco mosaic virus (TMV) deficient in the production of its native coat protein supports long-distance movement of a chimeric TMV. Proc. Natl. Acad. Sci. USA 1999, 96, 2549–2553. [Google Scholar] [CrossRef] [Green Version]
- Valkonen, J.P.T. Accumulation of potato virus Y is enhanced in Solanum brevidens also infected with tobacco mosaic virus or potato spindle tuber viroid. Ann. Appl. Biol. 1992, 121, 321–327. [Google Scholar] [CrossRef]
- Cohen, J.; Loebenstein, G.; Spiegel, S. Infection of sweet potato by cucumber mosaic virus depends on the presence of sweet potato feathery mottle virus. Phytopathology 1988, 72, 583–585. [Google Scholar] [CrossRef]
- Tribodet, M.; Glais, L.; Kerlan, C.; Jacquot, E. Characterization of Potato virus Y (PVY) molecular determinants involved in the vein necrosis symptom induced by PVYN isolates in infected Nicotiana tabacum cv. Xanthi. J. Gen. Virol. 2005, 86, 2101–2105. [Google Scholar] [CrossRef]
- Abt, I.; Jacquot, E. Wheat dwarf. In Virus Diseases of Tropical and Subtropical Crops; Tennant, P., Fermin, R., Eds.; Plant Protection Series; CAB International: Boston, MA, USA, 2015; pp. 27–41. [Google Scholar]
- Péréfarres, F.; Thébaud, G.; Lefeuvre, P.; Chiroleu, F.; Rimbaud, L.; Hoareau, M.; Reynaud, B.; Lett, J.M. Frequency-dependent assistance as a way out of competitive exclusion between two strains of an emerging virus. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133374. [Google Scholar] [CrossRef] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abt, I.; Souquet, M.; Angot, G.; Mabon, R.; Dallot, S.; Thébaud, G.; Jacquot, E. Functional Transcomplementation between Wheat Dwarf Virus Strains in Wheat and Barley. Viruses 2020, 12, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010034
Abt I, Souquet M, Angot G, Mabon R, Dallot S, Thébaud G, Jacquot E. Functional Transcomplementation between Wheat Dwarf Virus Strains in Wheat and Barley. Viruses. 2020; 12(1):34. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010034
Chicago/Turabian StyleAbt, Isabelle, Marlène Souquet, Gersende Angot, Romain Mabon, Sylvie Dallot, Gaël Thébaud, and Emmanuel Jacquot. 2020. "Functional Transcomplementation between Wheat Dwarf Virus Strains in Wheat and Barley" Viruses 12, no. 1: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010034