Role of Dendritic Cells in Exposing Latent HIV-1 for the Kill


1
Department of Infectious Diseases and Microbiology, Graduate School of Public Health, University of Pittsburgh, Pittsburgh, PA 15261, USA
2
Department of Pathology, School of Medicine, University of Pittsburgh, Pittsburgh, PA 15261, USA
*
Author to whom correspondence should be addressed.
Viruses 2020, 12(1), 37; https://0-doi-org.brum.beds.ac.uk/10.3390/v12010037
Submission received: 25 November 2019
/
Revised: 19 December 2019
/
Accepted: 24 December 2019
/
Published: 28 December 2019
(This article belongs to the Special Issue Dendritic Cells and Antiviral Defense)
Abstract
:The development of effective yet nontoxic strategies to target the latent human immunodeficiency virus-1 (HIV-1) reservoir in antiretroviral therapy (ART)-suppressed individuals poses a critical barrier to a functional cure. The ‘kick and kill’ approach to HIV eradication entails proviral reactivation during ART, coupled with generation of cytotoxic T lymphocytes (CTLs) or other immune effectors equipped to eliminate exposed infected cells. Pharmacological latency reversal agents (LRAs) that have produced modest reductions in the latent reservoir ex vivo have not impacted levels of proviral DNA in HIV-infected individuals. An optimal cure strategy incorporates methods that facilitate sufficient antigen exposure on reactivated cells following the induction of proviral gene expression, as well as the elimination of infected targets by either polyfunctional HIV-specific CTLs or other immune-based strategies. Although conventional dendritic cells (DCs) have been used extensively for the purpose of inducing antigen-specific CTL responses in HIV-1 clinical trials, their immunotherapeutic potential as cellular LRAs has been largely ignored. In this review, we discuss the challenges associated with current HIV-1 eradication strategies, as well as the unharnessed potential of ex vivo-programmed DCs for both the ‘kick and kill’ of latent HIV-1.
1. Introduction
Despite major advances in human immunodeficiency virus-1 (HIV-1) treatment and prevention since the discovery of the virus in 1983 [1], the global rate of new infections remains constant at approximately 2 million per year [2,3]. Although optimal antiretroviral therapy (ART) suppresses viremia to below the limit of detection of clinical assays, HIV-1 is managed as a chronic disease due to its persistence in a long-lived population of resting memory CD4+ T cells considered to be the major reservoir [4,5]. Early treatment can limit reservoir size but not prevent its establishment, which occurs within days of initial infection [6]. Based on reservoir decay kinetics following ART initiation, it was initially predicted that 2–3 years of therapy would be sufficient to eliminate all HIV-infected cells [7]. However, rebound of viremia to pretreatment levels in HIV-infected individuals who discontinued therapy provided evidence for a longer-lived population of latently infected cells [8]. With a t1/2 of 44 months, this pool of infected cells would require approximately 73 years of therapy to eradicate [9]. Finding a safe and effective means to specifically expose this HIV-1 cellular reservoir for immune targeting and elimination during ART remains a major barrier to a functional cure. In this review, we dissect the potential strengths and limitations of current HIV cure strategies, with a particular emphasis on recent studies employing dendritic cell (DC)-based HIV-1 immunotherapies.
2. Dendritic Cell Programming for Immunotherapy
Derived from the bone marrow, DC serve as bridge between the innate and adaptive immune systems due to their dual capacity to respond to pathogen- and danger-associated signals during acute inflammatory responses and to process and present antigens for the priming of naïve T cells [10]. Although the principal human DC subsets in vivo consist of three main types including plasmacytoid DCs (pDCs) and myeloid/conventional DC1s (cDC1s) and DC2s (cDC2s) [10], models of DC ontogeny have undergone numerous revisions since the discovery of DCs by Ralph Steinman and Zanvil Cohn [11,12] more than 40 years ago. Due to recent experimental data supporting modification of the classical model of hematopoiesis that regulates DC lineage, it is now accepted that DCs originate not from a population of multipotent progenitor cells but from progenitors that follow predestined developmental pathways determined by lineage priming [13,14,15,16].
2.1. Classification of Endogenous DCs
DCs are further classified based on differential expression of transcription factors such as interferon regulatory factors 4 and 8, which distinguish plasmacytoid DCs (pDCs, IRF4+IRF8+), myeloid/conventional DC1s (cDC1s, IRF4-IRF8+), and myeloid/conventional DC2s (cDC2s, IRF4+IRF8-) [10,17,18,19,20]. Characterized by surface expression of CD123 and CD45RA, pDCs produce high levels of type I and type III interferons in response to viral infection in the forms of single-stranded RNA and double-stranded DNA detected by endosomal toll-like receptors 7 and 9, respectively [10,21,22].
Myeloid cDC1s were initially classified as a subset of blood DCs expressing high levels of CD141 (thrombomodulin) [23,24] but also reside in numerous lymphoid and nonlymphoid tissues [10]. Identification of cDC1s is also based on shared expression of CD13 and CD33 with cDC2s and low expression of CD11b, CD11c, and CD172 [10]; unique to cDC1s is the receptor that recognizes actin exposed during cell necrosis, CLEC9A [25,26]. Myeloid DC1s have a superior capacity to cross-present viral and intracellular antigens in the context of both MHC class I and class II for activation of CD8+ T cells and the induction of T helper type 1 (TH1) and natural killer (NK) cell responses [27,28,29,30]. Like pDCs, cDC1s produce both type I and type III interferons in response to pathogen sensing via TLR3 and TLR9 [10,31,32,33].
The predominant human cDC population in blood, as well as lymphoid and nonlymphoid tissues, cDC2s are characterized by various surface antigens, including CD1c, CD2, CD172, FcεR1, CD11b, CD11c, CD13, and CD33 [10]. Similar to monocytes, cDC2s express an extensive repertoire of lectins and pattern recognition receptors that facilitate immune responses to agents as diverse as intracellular pathogens, extracellular bacteria, parasites, and fungi [10]. Furthermore, cDC2s are capable of inducing TH1, TH2, TH17, and TREG immune responses, due to their ability to secrete various pro- and anti-inflammatory mediators [10,34,35,36]. Of note, cDC2s are weak producers of type III interferons but can be stimulated to produce high levels of IL-12 compared to cDC1s [37,38,39,40].
2.2. Characterization of Ex Vivo-Programmed DCs
Ex vivo generation and programming of antigen-presenting, monocyte-derived DCs (MDCs) was first implemented in cancer immunotherapy to circumvent the dysfunction of endogenous DCs that occurs in cancer patients [41,42,43,44,45,46], capitalizing on the discovery of culture methods that were scalable to quantities required for clinical trials. In contrast to traditional vaccines that relied on antigen cross-presentation by patients’ own dysfunctional DCs, this strategy harnessed functional DCs generated outside of the tumor-associated suppressive environment to induce effective anti-cancer immunity [41,42,43,44,45,46]. This concept was based on the premise that MDCs could be imprinted during maturation with particular effector functions and homing properties for the induction of tumor-specific CD8+ T cells in vivo [47]. The first advantage of this approach is that ex vivo-generated DCs can be loaded with specific antigens that will promote effective delivery of both antigen (‘signal 1’) and costimulation (‘signal 2’) to antigen-specific T cells. However, the ability of DCs to cross-present antigen is affected by DC developmental stage and by the combination of factors used in their activation and maturation [48,49,50,51,52]. In addition, shortcomings of ‘first-generation’ DC vaccines highlighted the necessity for DCs to express high levels of costimulatory molecules and lymph node homing capacity in order to induce effective anti-cancer cytotoxic T lymphocyte (CTL) responses [47,53,54,55,56]. As a result, ‘second-generation’ DC maturation strategies incorporating PGE2 [57,58,59] were developed to induce fully mature DCs, with high expression of costimulatory molecules (CD83, CD86) and CCR7 (Table 1) [60,61,62,63,64] by activation via E-prostanoid receptors 2 (EP-2) and 4 (EP4) expressed on MDCs [65]. As a mediator of chronic inflammation, the addition of PGE2 mimics a microenvironment characteristic of a chronic inflammatory state such as that of a cancer setting. Although IL-1β/TNF-α/IL-6/PGE2-matured DCs displayed enhanced immunogenicity and migratory potential in response to lymph node-secreted chemokines CCL19 and CCL21 in healthy individuals [53,54,55,60,61], the negative impact of PGE2 on DC IL-12p70 production and their recruitment and activation of TREG cells [66,67,68,69] was linked to limited anti-tumor [70] and anti-HIV clinical responses [71].
Since high IL-12p70 production by DCs has been shown to greatly enhance their ability to induce antigen-specific TH1 cells and CTL (‘signal 3’) [75,76,77,78,79,80,81,82,83,84,85], subsequent efforts to improve the efficacy of DC-based cancer vaccines have focused on employing ‘nonexhausted’ DCs, unlike the PGE2- matured DCs [59,62,86,87]. These ‘type 1-polarized’ monocyte-derived DCs (MDC1s) could be generated in the presence of type I and type II interferons and TLR ligands, or IL-18-activated NK cells or memory CD8+ T cells [62,67,72,88,89,90,91,92,93]. Both in vitro and in mouse models, high IL-12p70-producing DCs induced strong, long-lived tumor-specific CTL responses [47]. Furthermore, addition of IFN-γ and the combination of cytokine maturation cocktail reversed maturation-associated DC exhaustion, producing polarized DCs capable of increased IL-12p70 production in response to interaction with CD40L-expressing CD4+ T cells [62,67,92,94], mimicking an acute, pro-inflammatory microenvironment typically associated with successful anti-viral responses. Subsequent findings revealed that maturing MDCs enhanced expression of CCR7 and production of chemokines (‘signal 4’) that promote the interaction of MDC1 with naïve, memory, and effector T cells (Table 1) [69]. Specifically, inclusion of IFN-α and the TLR 3 ligand polyinosinic:polycytidylic acid [poly (I:C)] in the ‘alpha type 1-polarized’ DC (αDC1) maturation cocktail was found to augment production of CXCL9, CXCL10, CXCL11, and CCL5 to promote the interaction of mature DCs with CXCR3- and CCR5-expressing CTL, TH1, and NK cells desirable for effector responses [63,69,73,74]. Likewise, IFN-α-induced maturation results in reduced production of CCL22 by MDC1, thus avoiding attraction of undesirable CCR4-expresssing TREG [69].
In sum, while differentially matured DCs can share similarities in antigen presentation, maturation status, and lymph node homing capacity, large discrepancies often exist in their capacities for optimal cytokine production, chemokine expression related to T cell trafficking, and antigen transfer (Table 1). Thus, correct ex vivo programming is critical to the design of DC-based immunotherapies in order to achieve the desired cellular mechanisms in vivo. Ex vivo generation of DCs allows manipulation of antigenic ‘signal 1’ and costimulatory ‘signal 2’ for optimal activation and expansion of both antigen-specific CD4+ and CD8+ T cells [47]. Education of DCs to further exploit polarizing ‘signal 3’ and homing ‘signal 4’ can selectively drive the desired antigen-specific T cell effector mechanisms and trafficking patterns. Together, such ex-vivo DC instruction could improve therapeutic outcomes by circumventing logistic issues related to the route of DC delivery, tissue-associated influences, cytokine exhaustion, and requirements for timely and functional interactions between DCs and T cells within lymphatic tissues [53,54,63,86,87,95,96,97,98].
3. Use of DCs to Address Hurdles to the CTL ‘Kill’ of HIV-1
Numerous clinical trials have shown that although HIV-1 proviral reactivation is achievable in vivo, the elimination of reactivated cells is minimal to nonexistent [99,100,101,102,103,104,105]. These results highlight that the ‘kill’ does not necessarily follow latency reversal, regardless of sufficient expression of viral RNA or antigens by infected cells [106,107]. CD8 depletion studies in SIV-infected nonhuman primates support the crucial role of CD8+ T cells in control of viral replication [108,109,110,111,112]. Likewise, human studies have found temporal associations between a peak in HIV-specific CD8+ T cell responses during acute infection, control of HIV viremia, and delayed disease progression [113,114,115]. A successful anti-HIV-1 response likely requires either the reactivation of memory CTL or the activation of de novo CTL responders capable of effective recognition of cells expressing peptide antigen derived from replication-competent proviruses following latency reversal [116]. While DC-based strategies may offer a route to educate and assemble an arsenal of CTLs, the challenges associated with effective targeting of latently infected cells are multifactorial.
3.1. Mutations in CTL Epitopes
CTL recognition of HIV-1 epitopes is hampered by escape mutations that are selected early in infection due to heightened immune pressure [117,118,119,120] and continue to contribute to the latent reservoir until the initiation of ART [116]. Proviruses with escape mutations to immunodominant CTL epitopes comprise a significant portion of the latent reservoir in individuals who initiate ART in the chronic stage of HIV-1 infection [121]. In addition to the establishment of CTL escape variants, minor viral modifications to CTL epitopes can also occur, leading to a partial or incomplete immune escape resulting in the induction of dysfunctional cross-reactive memory responses characterized by the production of proinflammatory cytokines by responding CTLs in the absence of target cell killing [74,122]. In a sense, this can create type of immunologic smokescreen, which could be even more problematic than the establishment of a complete CTL escape variant, since the responding effector cells can act as competitive inhibitors for the induction and effector activity of new antigen-specific CD8+ T cells. Such dysfunctional cross-reactive CTLs could negatively impact the afferent phase of the immune response by competing for antigen presented by DCs, potentially affecting their capacity to generate de novo killer CTL responses against the newly established variants, as well as to block the ability of newly activated CTLs to target the infected cells in the periphery during the efferent phase of the immune response. Moreover, cytokines produced by cross-reactive CTLs targeting these adaptive epitopes can create an environment to promote chronic inflammation and DC-mediated HIV-1 dissemination [74,123]. Using MDC1 to selectively prime HIV-1-specific CD8+ T cells from naïve T cell precursors while avoiding pre-existing, dysfunctional memory T cells has been shown effective in vitro for targeting persistent HIV in individuals on ART [122].
Thus, successful CTL-based HIV eradication strategies should target subdominant viral epitopes or epitopes for which escape mutations have not accumulated [121]. Graph-based epitope selection strategies using mathematical algorithm program tools such as Epigraph can be used to assess global epitope diversity and/or conservation when designing DC vaccine antigenic formulations [124]. An alternative to this approach is to base antigen selection according to biochemical network analyses of CD8+ T cell epitope topology based on HIV proteome data, which has revealed an inverse relationship between protective CTL epitopes targeted by HIV controllers and mutational frequency in vivo [125]. An epitope that is determined to have a high network value indicates that it is contained within an important structural region of the viral proteome, critical for maintaining the three-dimensional protein structure and function, and thus important to the overall fitness of the virus. Utilizing DCs to selectively re-focus the CTL response away from highly variable antigenic targets and towards these critical regions of the virus is an attractive notion.
3.2. Dysfunctional or Exhausted CTLs
CTL exhaustion is another critical consideration in the design of effective kill-phase strategies. T cell exhaustion can occur in the settings of chronic viral infections or cancer, as a result of persistent immune activation and antigen exposure [116,126,127,128,129]. Among defects in CTL function that arise during chronic HIV-1 infection, exhaustion cannot be fully reversed with ART [130]. Exhaustion in antigen-specific T cells involves a gradual loss of effector function, accompanied by decreased proliferative capacity [131]. Compared to terminally differentiated or memory CD8+ T cells, exhausted CD8+ T cells are characterized by decreased cytokine production and cell surface expression of inhibitory receptors including PD-1, TIM-3, CTLA4, CD160, LAG-3, and TIGIT [131,132,133,134,135,136]. Increases in inhibitory receptor expression serve as immune checkpoints (ICs) to limit excess T cell activation during a normal immune response and are downregulated upon its resolution [134,137]. Conversely, sustained expression of inhibitory receptors results from persistent antigen exposure, leading to an exhausted phenotype. Even in the context of suppressive ART, in which exposure to HIV antigen is limited, it may not be possible to completely restore CTL functionality [116]. This is illustrated by high levels of PD-1 and TIGIT expression by both follicular and nonfollicular CD8+ T cells in the lymph nodes of HIV-infected individuals [138]. However, immune checkpoint blockade combination therapies approved for use in cancer immunotherapy have been shown to reverse exhaustion to varying degrees in virus-specific T cells [128,136,139,140]. In vitro blockade of the PD-1/PD-L1 axis in follicular and nonfollicular lymph node cells was shown to restore HIV-1-specific CD8+ T cell function [138], and a phase I clinical trial of the anti-PD-L1 antibody BMS-936559 demonstrated a trend toward enhanced Gag-specific CD8+ T cell responses in virally suppressed participants [141] Importantly, correlations between HIV DNA content in memory CD4+ T cells and expression of PD-1, TIGIT, and LAG-3 inhibitory receptors have been documented, suggesting a potential role for these ICs in latency establishment [116,142,143].
3.3. Defective HIV as Antigen Decoys
Another challenge of the ‘kill’ phase is the excess of defective proviruses that occupy the latent reservoir in vivo. These defective proviruses (HIV-1def) can result from hypermutations, large internal deletions, packaging signal deletions, major splice donor (MSD) mutations, or inactivating point mutations [144,145,146], and significantly outnumber cells harboring replication-competent proviruses. As a result of intact LTR promoter function, integration into actively transcribed genes, and lack of promoter methylation, some HIV-1def can be transcribed, translated, and recognized by HIV-specific CTLs [145,146,147], thus serving as decoy targets that act to divert the CTL-mediated elimination of the true latent reservoir [145,148]. It has been postulated that preferential targeting of HIV-1def may be a function of their ability to present antigen to CTLs more efficiently as a result of a defect in Nef and the inability to downregulate MHC class I [145]. Evidence that clonal expansion of cells containing both defective and replication-competent HIV-1 shapes the pool of latently infected cells in individuals on long-term ART [149,150,151,152,153] poses an additional challenge to targeting only those proviruses.
3.4. Spatial Separation of CTL and Target Cells
One of the greatest challenges of targeting the HIV reservoir for CTL-mediated elimination is the spatial separation of target and effector cells in anatomical ‘sanctuaries’ [154] that serve as a persistent source of latent provirus. For example, the CD4+ T follicular helper (TFH) and T follicular regulatory (TFR) cell subsets within B cell follicles of lymph nodes are more permissive to HIV-1 infection than extrafollicular subsets ex vivo [155,156,157] and comprise a major portion of the latent reservoir during both chronic SIV and HIV infection [158,159,160,161]. Whereas CXCR5+ CD8+ T cells have been identified within the B cell follicles of secondary lymphoid organs during chronic viral infections [162,163,164], CTLs could be excluded from the B cell follicle because they usually lack the follicular homing receptor CXCR5, which is not expressed until late in untreated infection [158]. As a result, target cells within the follicle are protected from CTL-mediated cytolysis, whereas viral replication in extrafollicular zones is efficiently controlled [165]. Indeed, ongoing SIV replication within TFH cells of elite controller macaques has been attributed to CTL exclusion from B cell follicles, as depletion of SIV-specific CD8+ T cells resulted in increased viral replication in non-TFH CD4+ T cells [154,166].
Mechanisms governing CTL migration to and cytolytic activity within B cell follicles are poorly understood, and novel strategies to address barriers to CTL targeting of this compartment are being explored. Among these, temporary disruption of the B cell follicle with depleting antibodies (anti-CD20/rituximab), blockade of T and B cell interactions using anti-CD40L, and therapeutic vaccination with engineered CTL or chimeric antigen receptor T cells expressing CXCR5 have been proposed [154,165]. Transduction of CTLs with CXCR5 to facilitate B cell follicle homing [167] and in vitro stimulation with TGF-β to induce CXCR5 expression in CXCR5-CD8+ T cells has been demonstrated in rhesus macaques [168]. Another promising approach involves the use of bispecific antibodies targeting CD3 and HIV gp120 that have the dual capacity to act as latency reversal agents (LRAs), through a CD3-mediated activation of CD4+ T cells containing HIV provirus, and to facilitate CD8+ T cell-mediated ADCC of the reactivated HIV-infected CD4+ T cells induced to express Env protein. [169]. Although this remains to be tested in vivo, lymph node-homing CXCR5+CD8+ T cells could be transduced to produce these antibodies in order to facilitate killing of latently infected cells within the B cell follicle [165]. A better understanding of the signals required to drive CXCR5 expression in CTLs during priming could lead to the optimal design of ex-vivo-educated DCs purposely programmed to provide such needed factors.
4. Driving HIV-1 out of Hiding: Current Status of Latency Reversal Approaches
Beyond spatial separation of target and effector cells, the greatest obstacle to the ‘kill’ is HIV-1 latency itself. The ‘kick and kill’ approach to HIV eradication entails proviral reactivation during ART, coupled with generation of CTLs or other immune effectors equipped to eliminate exposed infected cells [170]. Effective yet nontoxic strategies to target the latent reservoir remain elusive, due to limited post-integration proviral gene expression that spares infected cells from viral cytopathic effects or effector-mediated clearance [171].
4.1. Common LRA Strategies
Perhaps the largest class of LRAs in current use, epigenetic modifiers, include histone deacetylase inhibitors (HDACi), histone methyltransferase inhibitors (HMTi), DNA methyltransferase inhibitors (DNMTi), bromodomain and extraterminal (BET) bromodomain inhibitors (BETi), positive transcription elongation factor b (P-TEFb) activators, and hexamethylene bisacetamide (HMBA) [172,173]. All of these compounds operate at various levels of transcriptional control to mediate changes in chromatin structure that permit proviral reactivation. Among these, HDACi have been most extensively studied, and clinical trials of vorinostat, panobinostat, and romidepsin have all demonstrated HIV reactivation in vivo [99,102,104,105,174]. However, none of these agents impacted levels of HIV-1 DNA.
PKC agonists have been shown to induce HIV LR through activation of NF-κB signaling by intracellular PKC isoforms [173]. Within this class of LRAs, the phorbol esters prostratin and 12-deoxyphorbol 13-phenylacetate (DPP) have both demonstrated potential as T cell activators and LRAs in latently infected primary cells ex vivo [175,176]. In addition, the PKC agonist bryostsin-1 has been used to achieve potent T cell stimulation both in vitro and ex vivo [177,178]. However, its association with adverse effects in clinical oncology trials [179,180,181,182] prompted conservative drug dosing of bryostatin-1 in a subsequent HIV-1 clinical trial, resulting in neither PKC activation nor HIV reactivation [103].
Importantly, both ex vivo studies and clinical trials have demonstrated that no single pharmacological LRA in current use has been able to reactivate a significant proportion of the latent HIV-1 reservoir [183,184]; therefore, combination LRA approaches are being explored [173]. An additional complication of proposed pharmacological LRAs is that many demonstrate the capacity to negatively impact CTL and NK cell effector function [178,185,186,187].
4.2. Next Generation Pharmacological LRAs?
Among LRAs that have shown the most promise, TLR agonists have been investigated in several ex vivo and in vivo studies [188,189,190]. The TLR7 agonist GS-9620 (Vesatolimod) in combination with the HIV envelope-specific broadly neutralizing antibody (bNAb) PGT121 induced extracellular HIV-1 RNA and increased cytolytic activity in PBMCs of virally suppressed individuals [188]. Though the mechanism of HIV reactivation in this study has not been fully elucidated, both latency reversal and T cell activation were dependent on type I IFNs produced through TLR7 stimulation of pDCs. Co-administration of PGT121 and Vesatolimod during ART also delayed viral rebound in SHIV-infected rhesus macaques upon ART discontinuation [190]. Finally, combined Ad26/MVA therapeutic vaccination/TLR7 stimulation was shown to increase the breadth of SIV-specific CTL responses, decrease levels of SIV DNA, and delay viral rebound following ART interruption in SIV-infected rhesus macaques [189]. Vesatolimod has since advanced to human trials in both ART-suppressed participants (NCT02858401) and HIV-1-infected controllers on ART and during ATI (NCT03060447).
The efficacy of TLR9 agonist MGN1703 as an innate immunity enhancer and LRA was evaluated in HIV-1-infected, ART-suppressed individuals (http://www.clinicaltrials.gov NCT02443935) [191]. Whereas activation of pDCs, increases in plasma IFN-2α and HIV-1 RNA, upregulation of ISG transcription, and increased proportions of activated NK and CD8+ T cells resulted from MGN1703 treatment, dosing did not impact reservoir size [191]. The TLR9 agonist lefitolimod in combination with bNAbs is currently being evaluated in HIV-1-infected individuals on ART and during ATI for its impact on HIV-1 reservoir reduction (NCT03837756).
Finally, in addition to its ability to enhance HIV-specific CTL responses [165,192], IL-15 is being investigated for its LRA potential as a novel LRA. IL-15 is a product of activated DCs, which stimulates antiviral NK and CD8+ T cell responses [193,194,195] when presented in trans as part of a membrane-bound IL-15:IL-15Rα complex [194,196]. IL-15 superagonists recapitulating this biologically potent heterodimer functionality are being explored as potential LRAs [192]. Both IL-15 and the IL-15 superagonist ALT-803 induced LR activity in a primary CD4+ T cell model of HIV latency, and ALT-803 also enhanced CTL killing of HIV-infected cells ex vivo. In addition to being evaluated in human cancer trials (NCT01946789, NCT01885897, NCT02099539), dose escalation studies of ALT-803 are being performed to assess whether it can be tolerated at doses deemed safe in nonhuman primates.
5. Dual Role for DCs in the ‘Kick and Kill’?
5.1. DCs as a Therapeutic Tool to Drive HIV-1-Specific Killer T cells
A revolutionary study by Lu et al. in SIV-infected rhesus macaques revealed the promise of therapeutic dendritic cell vaccination using inactivated SIV-loaded autologous DCs [197]. Three immunizations elicited a 50-fold decrease in SIV DNA and a 1000-fold decrease in SIV RNA in peripheral blood that were sustained throughout the study and correlated with increased SIV-specific cellular and humoral responses. These impressive results were replicated in a subsequent trial in chronically HIV-infected, untreated individuals who exhibited prolonged post-vaccination suppression of viral load that was attributed to strong virus-specific CD4+ T helper and CD8+ effector responses [198].
An early DC-based HIV immunization strategy developed by our group implemented autologous mature DCs pulsed with HLA*A02-restricted HIV-1 Gag, Pol, and Env peptides and influenza A matrix protein peptide administered to participants intravenously or subcutaneously [199]. Although the peptide-DC vaccine elicited HIV-specific IFN-γ responses at two weeks following the second immunization, the DCs used were suboptimal for the induction of long-lived, broadly reactive CTL responses. However, one of the most impressive HIV immunotherapy trials to date utilized DCs pulsed with inactivated autologous HIV, which resulted in a 1 log10 decrease in HIV RNA setpoint and was associated with increased anti-HIV CD8+ T cell IFN-γ responses [200]. Nonetheless, as with many of these earlier DC-based studies, this trial implemented DC generation methods that yield IL-12p70-deficient DCs incapable of inducing sustained HIV-specific effector responses. In an attempt to address this issue, Argos Therapeutics investigated ex vivo genetic manipulation of DCs as a strategy to deliver a constitutive CD40L helper signal to the DCs in an HIV immunotherapy to treat acute and chronic infections [201,202,203]. Autologous monocyte-derived DCs were co-electroporated with synthetic CD40L RNA and HIV RNA encoding Gag, Nef, Vpr, and Rev derived from individuals’ pre-ART plasma to create the personalized AGS-004 vaccine [204]. Nevertheless, this approach was unsuccessful, which may have been due to the fact that constitutive CD40L signaling induces an early burst of IL-12p70 production, but ultimately creates IL-12p70-exhausted DCs that are unresponsive to CD4+ TH cell interaction [122].
A novel therapy proposed by Guardo et al. combined TRIMIX adjuvant and an HIV T cell immunogen (HTI) for in vivo targeting of DCs by intranodal injections [205]. The previously described TRIMIX adjuvant consists of three mRNAs encoding CD40L, the costimulatory molecule CD70, and constitutively activated TLR4 [206]. The HTI vaccine component consists of an mRNA expressing epitopes of Gag, Pol, Vif, and Nef proteins, chosen on the basis of antigen-specific CD4+ and CD8+ T cell reactivity [207]. Monocyte-derived DCs electroporated with this preparation were shown to induce T cell proliferation and IFN-γ responses in vitro, and intranodal injection of TRIMIX/HTI induced antigen-specific CTL responses in mice [205]. In addition, human lymph node explants treated with TRIMIX/HTI activated DCs and induced proinflammatory mediator production. However, the IL-12-producing capacity of the mRNA/DC-based formulation was not investigated in this study, therefore providing no information regarding its potential to induce broadly reactive CTLs required for the long-term control of viremia in the absence of ART [208].
More recently, Surenaud et al. reported increased HIV-specific CD8+ and CD4+ T cell responses in patients on ART following therapeutic vaccination with DCs generated ex vivo in the presence of IFN-α and loaded with a combination of Gag, Pol, and Nef peptides (LIPO-5) [209]. HIV-specific CD8+ and CD4+ T cells targeted both dominant and subdominant epitopes of Gag, Pol, and Nef and elicited IL-2 and IL-13 production by CD4+ T cells that significantly correlated with control of viremia upon analytic treatment interruption (ATI). In addition to predicted epitopes, the vaccine also elicited CD4+ T cell responses against previously unidentified HIV-1 HLA-DR-restricted CD4+ T cell epitopes, highlighting the importance of CD4+ T cell responses in control of HIV-1 viremia.
Overall, although DC-based HIV-1 immunotherapies have proven to be safe and well tolerated, they have achieved a success rate of only 38% according to meta-analyses [210]. Furthermore, while there have been many DC-based therapeutic approaches explored in HIV clinical trials, there has been little consensus with regards to the selection of subjects, methods of DC generation and preparation, choice of immunogen, and assessment of immunologic and virologic responses [211]. These parameters will be critical to the design of future trials, and although beyond the scope of this review, efforts are being made to compare and interpret outcomes of DC-based HIV vaccination strategies to date.
5.2. LRA Potential of DCs
Although DCs have been used extensively for the induction of antigen-specific T cell responses in HIV-1 clinical trials [199,200,212,213], their capacity to function as an LRA has only gained recent attention. In fact, as DCs are intended to engage in cognate interaction with CD4+ T cells, it is not surprising that they have been implicated in both HIV trans infection [214,215] and latency establishment [216,217]. For the same reason, it is plausible that DCs could be utilized to reactivate provirus from these same cells. Interestingly, results of a recent phase I/II clinical trial developed by our group linked the administration of DCs loaded with autologous HIV that were designed to induce CTL responses [218] with post-vaccination increases in plasma viremia in ART-suppressed participants [219]. As the aim of that study was not to evaluate the efficacy of the DC therapeutic as an LRA, pertinent questions regarding the underlying mechanisms of the observed proviral reactivation remain.
Historically, most DC-based strategies aimed at the ‘kick’ of latent HIV in vitro have employed monocyte-derived immature DCs (iDCs) and DCs matured by exposure to bacterial antigens [220,221,222]. Among these studies, several have purported that DC-mediated LR could be induced by DC–T cell contact in the absence of antigen presentation and/or mediated by DC-secreted soluble factors. For example, van der Sluis et al. reported that the interaction of MDC with actively proliferating primary T cells resulted in secretion of unidentified components by DCs to induce latent provirus [220]. However, in this study, iDCs were cultured with T cells that had been preactivated using either PHA or anti-CD3/CD28 beads. These treatments alone have been shown to reverse HIV-1 latency [184,223,224,225,226], and thus, it is difficult to dissect the clinical relevance.
In another study, CD40L-transduced DCs were reported to induce latency reversal in J-Lat and ACH-2 cell lines, independent of DC–T cell contact and mediated by TNF-α [227]. Similarly, MDCs matured by microbial components related to AIDS-associated pathogens, including Mycobacterium bovis, Bacillus Calmette-Guérin (BCG), and lipopolysaccharide (LPS), were shown to reactivate latent HIV in Jurkat T cells through secretion of TNF-α [222]. Although the incorporation of antigen in DC-mediated LR has been underexplored, Marini et al. successfully utilized iDCs loaded with SEB superantigen for reactivation of HIV-1 in in vitro–infected CD4+ T cell lines [221]. Furthermore, van Montfort and colleagues determined that iDCs in combination with TCR stimulation more efficiently reversed latency in primary HIV-infected cells from aviremic individuals than TCR activation alone, a phenomenon that was dependent on PI3K-Akt-mTOR pathway activation [228]. Finally, a study investigating the latency activation potential of various DC subsets demonstrated that tissue-resident and blood-derived myeloid DCs reactivated virus in vitro from latently infected effector T cells with different efficiencies [229]. Nevertheless, none of these DCs therapeutics was designed with the dual purpose of creating sufficient HIV-1 antigen exposure for successful CTL targeting of the CD4+ T cell reservoir. Information regarding the antigen specificity of CD4+ T cells harboring HIV would be an important step in designing a DC-based therapy to directly activate viral production from these cells.
5.3. Using DCs to Expose Pathogen-Specific HIV Reservoirs?
Previous studies have shown that HIV-1 preferentially infects and depletes HIV-specific CD4+ T cells of HIV infected individuals [230,231], but some of these latently infected cells survive and retain their antigen specificity during ART [232]. Therefore, HIV-1 specific CD4+ T cells may themselves harbor a portion of the long-lived latent reservoir and can potentially be exposed upon their antigen-specific activation. This could partially explain why our early HIV-1 antigen-loaded DC-based therapy was found to subsequently drive residual viremia in ART-suppressed individuals following ATI [219].
Despite that it has been reported that CMV-specific CD4+ T cells are less susceptible to HIV-1 infection in vivo [233], they were still found to be susceptible to a degree. Moreover, susceptibility to infection may not be directly related to the capacity of these cells to survive and clonally expand to harbor persistent infection over time. In fact, a large body of data exists to support the notion that there is a sizeable contribution of CMV antigen-specific CD4+ T cells to the latent reservoir. For example, the seroprevalence of CMV among HIV-infected individuals is extremely high, with conservative estimates being greater than 95% [234], and levels of CMV-specific CD4+ T cells as great as 25% have been documented in those who are coinfected [235,236,237,238]. In the mucosal and peripheral tissues of HIV-1-infected individuals, CMV hijacks human cytokine/chemokine signaling to fuel inflammation, thereby augmenting its own replication [239]. Subclinical CMV replication not only creates an environment conducive to reservoir seeding by recruiting target cells to sites of inflammation that may be shared sites of HIV persistence [240,241,242,243], but ultimately contributes to T cell dysfunction, impaired immune cell recovery, and chronic immune activation during ART [244,245,246,247]. Interestingly, Agudelo-Hernandez et al. [248] found a negative correlation between higher rates of subclinical herpes virus shedding and levels of inflammation and immune activation in virally suppressed, HIV-infected men. Nevertheless, CMV replication in the gut of ART-suppressed individuals has been associated with inflammation, mucosal barrier damage, and microbial translocation [240]. Finally, upregulation of CCR5 expression on susceptible target cells in the shared inflammatory environment triggered by CMV [249] and its ability to manipulate AP-1 and NF-κB signaling for the induction of HIV-1 gene expression [250,251,252,253] also serve to amplify HIV infection in CMV-coinfected individuals.
If established within CMV-specific CD4+ T cells, the latent HIV-1 reservoir could undergo cellular proliferation or clonal expansion in response to chronic CMV antigenic stimulation [242]. Data from several studies corroborate this theory. For example, a cross-sectional study of ART-naïve and ART-suppressed participants revealed a correlation between elevated levels of HIV-1 DNA and CMV replication in semen and peripheral blood [254,255]; another associated delayed decay of HIV DNA reservoirs with CMV replication in PBMC of men initiating ART during acute infection [243]. In addition, proviral and integration site analyses point to clonal expansion of latently infected cells as a major driver of HIV persistence under suppressive ART [149,150,153,256,257]. Evidence of selective increases in HIV DNA within CMV- and EBV-specific CD4+ T cells after immune reconstitution in HIV-infected patients who received chemotherapy for CMV- and EBV-associated malignancies illustrates this phenomenon [258]. Finally, self-renewal of stem cell memory T cells (TSCM), which were reported to contain the most copies of integrated provirus per cell in HIV-1-infected individuals, has been implicated to play a role in expansion of the latent HIV reservoir [259]. Of note, CMV-/HIV-coinfected persons possess functional CMV-specific TSCM that may undergo homeostatic proliferation during ART to promote reservoir expansion [260]. Coincidentally, expanded CMV-specific CD4+ T cells harboring latent HIV could be protected from elimination as a consequence of numerous immune evasion strategies that CMV has evolved, including production of decoy viral homologues of HLA class I molecules [261,262] and cytokines [263,264], hijacking of cell signaling pathways [142,263,264,265], and inhibition of apoptosis [266,267]. These findings suggest that CMV coinfection contributes to establishment and maintenance of the HIV-1 reservoir, and that CMV antigen-loaded DCs could offer a means to selectively expose a portion of the HIV-1 reservoir contained within CMV specific CD4+ T cells.
5.4. DC Potential as an All-in-One ‘Kick and Kill’ Tool
In a recent study from our group, we demonstrated the ability of a clinically applicable DC-based therapeutic strategy to facilitate both the ‘kick’ and ‘kill’ of latent HIV-1 in CD4+ T cells from chronically infected, ART-suppressed individuals (Figure 1). Antigen-presenting type 1-polarized DC (MDC1) induced the transcription of replication-competent provirus, as well as HIV-1-specific CTL responses that targeted exposed infected cells for immune elimination [268]. The induction of strong antigen-specific CTL responses is attributed to uniquely high IL-12 production by MDC1 in response to interaction with CD40L-expressing CD4+ T helper cells [64]. Such MDC1s have already been successfully implemented in clinical oncology trials [47,80,269,270,271,272] due to their superior ability to drive long-lived CTL responses from naive T cell precursors [273,274] and have been approved for use in phase I HIV clinical trials. Likewise, strategic loading of MDC1 with MHC class II viral antigens (CMV, HIV-1) to facilitate interaction with CD4+ TH cells reactivated latent provirus. Thus, this dual therapeutic approach could prove a safer, more targeted strategy for purging the latent HIV-1 cellular reservoir.
Notably, there were some caveats and limitations to our study. Although the underlying mechanisms of MDC1-mediated latency reversal have not been fully elucidated, bidirectional cross-talk between MDC1 and CD4+ T cells during cognate antigen-driven interaction involving CD40/CD40L signaling plays a role in the observed LR activity [268]. Furthermore, while antigen-driven, we cannot exclude the possibility that the MDC1-mediated proviral reactivation demonstrated represents nonspecific bystander effects of a potent antigen-specific response, instead of a direct impact on an antigen-specific HIV reservoir. As such, these questions warrant further investigation.
6. Concluding Remarks
While current data from our group and others indicate that the pool of both CMV- and HIV-1-specific CD4+ T cells contributes to the latent HIV-1 reservoir, it is probable that CD4+ T cells specific to other viruses that manifest as chronic infections, especially EBV and other herpes viruses, also harbor replication-competent provirus. It is well documented that secondary lymphoid organs such as the spleen, lymph nodes, and GALT are the major sites of HIV replication and persistence, and targeting the true latent reservoir within B cell follicles of these anatomical sanctuaries poses numerous challenges with regard to antigen delivery by a DC vaccine in vivo [160]. Furthermore, studies in nonhuman primates have clearly demonstrated that viral RNA may be found in a diverse array of organ systems not limited to lymphoid tissues, including the brain, kidneys, liver, and lungs [275,276,277]. As previously detailed, novel strategies to address immune targeting of these compartments are being developed [138,154,165,169,192]. As such, the LRA potential of other antigen-presenting cells such as B cells could be considered as strategy for targeting TFH within lymph node follicles. Additionally, to broadly target other anatomical sites, exploring the use of DC-derived exosomes might prove useful, since they may be able to widely circulate and present DC-associated proteins and/or antigens to T cells throughout the body, and have been shown to have T cell activating capacity [278]. Alternatively, developing methods for in vivo antigen delivery to resident antigen-presenting cells in these compartments should be pursued. Nevertheless, successful cure strategies will employ nontoxic, directed inducers of latency reversal, coupled with methods to augment immune effector functions. Mounting evidence suggests that this goal will entail identification of the antigen specificities of latent cellular reservoirs and may be best achieved through incorporation of antigen-presenting ex vivo-educated DCs.
Author Contributions
J.K. and R.B.M. contributed to the writing and original draft preparation; C.R.R. contributed to the review and editing; C.R.R. and R.B.M. contributed to the conceptualization, supervision, and funding and resources. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by NIH grants R21-A131763, U01-AI35041, UM1-AI126603, and T32-AI065380.
Acknowledgments
The authors would like to thank Phalguni Gupta for his support and thought-provoking comments and discussions.
Conflicts of Interest
The authors declare no conflict of interest, and the funders had no role in the writing of the manuscript or in the decision to publish this review.
References
- Barre-Sinoussi, F.; Chermann, J.C.; Rey, F.; Nugeyre, M.T.; Chamaret, S.; Gruest, J.; Dauguet, C.; Axler-Blin, C.; Vezinet-Brun, F.; Rouzioux, C.; et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science 1983, 220, 868–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- UNAIDS. Global AIDS Update 2018. Available online: https://www.unaids.org/en/20180718_GR2018 (accessed on 19 November 2019).
- UNAIDS. UNAIDS Data 2018. Available online: https://www.unaids.org/sites/default/files/media_asset/unaids-data-2018_en.pdf (accessed on 19 November 2019).
- Ruelas, D.S.; Greene, W.C. An integrated overview of HIV-1 latency. Cell 2013, 155, 519–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finzi, D.; Blankson, J.; Siliciano, J.D.; Margolick, J.B.; Chadwick, K.; Pierson, T.; Smith, K.; Lisziewicz, J.; Lori, F.; Flexner, C.; et al. Latent infection of CD4+ T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nat. Med. 1999, 5, 512. [Google Scholar] [CrossRef] [PubMed]
- Chun, T.W.; Engel, D.; Berrey, M.M.; Shea, T.; Corey, L.; Fauci, A.S. Early establishment of a pool of latently infected, resting CD4+ T cells during primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 1998, 95, 8869–8873. [Google Scholar] [CrossRef] [Green Version]
- Perelson, A.S.; Essunger, P.; Cao, Y.; Vesanen, M.; Hurley, A.; Saksela, K.; Markowitz, M.; Ho, D.D. Decay characteristics of HIV-1-infected compartments during combination therapy. Nature 1997, 387, 188–191. [Google Scholar] [CrossRef]
- Davey, R.T., Jr.; Bhat, N.; Yoder, C.; Chun, T.W.; Metcalf, J.A.; Dewar, R.; Natarajan, V.; Lempicki, R.A.; Adelsberger, J.W.; Miller, K.D.; et al. HIV-1 and T cell dynamics after interruption of highly active antiretroviral therapy (HAART) in patients with a history of sustained viral suppression. Proc. Natl. Acad. Sci. USA 1999, 96, 15109–15114. [Google Scholar] [CrossRef] [Green Version]
- Siliciano, J.D.; Kajdas, J.; Finzi, D.; Quinn, T.C.; Chadwick, K.; Margolick, J.B.; Kovacs, C.; Gange, S.J.; Siliciano, R.F. Long-term follow-up studies confirm the stability of the latent reservoir for HIV-1 in resting CD4+ T cells. Nat. Med. 2003, 9, 727. [Google Scholar] [CrossRef]
- Collin, M.; Bigley, V. Human dendritic cell subsets: An update. Immunology 2018, 154, 3–20. [Google Scholar] [CrossRef]
- Steinman, R.M.; Cohn, Z.A. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J. Exp. Med. 1973, 137, 1142–1162. [Google Scholar] [CrossRef]
- Steinman, R.M.; Cohn, Z.A. Identification of a novel cell type in peripheral lymphoid organs of mice. II. Functional properties in vitro. J. Exp. Med. 1974, 139, 380–397. [Google Scholar] [CrossRef] [Green Version]
- Paul, F.; Arkin, Y.; Giladi, A.; Jaitin, D.A.; Kenigsberg, E.; Keren-Shaul, H.; Winter, D.; Lara-Astiaso, D.; Gury, M.; Weiner, A.; et al. Transcriptional Heterogeneity and Lineage Commitment in Myeloid Progenitors. Cell 2015, 163, 1663–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notta, F.; Zandi, S.; Takayama, N.; Dobson, S.; Gan, O.I.; Wilson, G.; Kaufmann, K.B.; McLeod, J.; Laurenti, E.; Dunant, C.F.; et al. Distinct routes of lineage development reshape the human blood hierarchy across ontogeny. Science 2016, 351, aab2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velten, L.; Haas, S.F.; Raffel, S.; Blaszkiewicz, S.; Islam, S.; Hennig, B.P.; Hirche, C.; Lutz, C.; Buss, E.C.; Nowak, D.; et al. Human haematopoietic stem cell lineage commitment is a continuous process. Nat. Cell Biol. 2017, 19, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamitros, D.; Stoilova, B.; Aboukhalil, Z.; Hamey, F.; Reinisch, A.; Samitsch, M.; Quek, L.; Otto, G.; Repapi, E.; Doondeea, J.; et al. Single-cell analysis reveals the continuum of human lympho-myeloid progenitor cells. Nat. Immunol. 2018, 19, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Dutertre, C.A.; Scott, C.L.; McGovern, N.; Sichien, D.; Chakarov, S.; Van Gassen, S.; Chen, J.; Poidinger, M.; De Prijck, S.; et al. Unsupervised High-Dimensional Analysis Aligns Dendritic Cells across Tissues and Species. Immunity 2016, 45, 669–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilliams, M.; Ginhoux, F.; Jakubzick, C.; Naik, S.H.; Onai, N.; Schraml, B.U.; Segura, E.; Tussiwand, R.; Yona, S. Dendritic cells, monocytes and macrophages: A unified nomenclature based on ontogeny. Nat. Rev. Immunol. 2014, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Heidkamp, G.F.; Sander, J.; Lehmann, C.H.K.; Heger, L.; Eissing, N.; Baranska, A.; Luhr, J.J.; Hoffmann, A.; Reimer, K.C.; Lux, A.; et al. Human lymphoid organ dendritic cell identity is predominantly dictated by ontogeny, not tissue microenvironment. Sci. Immunol. 2016, 1, eaai7677. [Google Scholar] [CrossRef]
- Granot, T.; Senda, T.; Carpenter, D.J.; Matsuoka, N.; Weiner, J.; Gordon, C.L.; Miron, M.; Kumar, B.V.; Griesemer, A.; Ho, S.H.; et al. Dendritic Cells Display Subset and Tissue-Specific Maturation Dynamics over Human Life. Immunity 2017, 46, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Bao, M.; Liu, Y.J. Regulation of TLR7/9 signaling in plasmacytoid dendritic cells. Protein Cell 2013, 4, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef]
- Dzionek, A.; Fuchs, A.; Schmidt, P.; Cremer, S.; Zysk, M.; Miltenyi, S.; Buck, D.W.; Schmitz, J. BDCA-2, BDCA-3, and BDCA-4: Three markers for distinct subsets of dendritic cells in human peripheral blood. J. Immunol. 2000, 165, 6037–6046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, K.P.; Munster, D.J.; Clark, G.J.; Dzionek, A.; Schmitz, J.; Hart, D.N. Characterization of human blood dendritic cell subsets. Blood 2002, 100, 4512–4520. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, S.; Zelenay, S.; Sancho, D.; Hanc, P.; Kjaer, S.; Feest, C.; Fletcher, G.; Durkin, C.; Postigo, A.; Skehel, M.; et al. F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells. Immunity 2012, 36, 635–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.G.; Czabotar, P.E.; Policheni, A.N.; Caminschi, I.; Wan, S.S.; Kitsoulis, S.; Tullett, K.M.; Robin, A.Y.; Brammananth, R.; van Delft, M.F.; et al. The dendritic cell receptor Clec9A binds damaged cells via exposed actin filaments. Immunity 2012, 36, 646–657. [Google Scholar] [CrossRef] [Green Version]
- Haniffa, M.; Shin, A.; Bigley, V.; McGovern, N.; Teo, P.; See, P.; Wasan, P.S.; Wang, X.N.; Malinarich, F.; Malleret, B.; et al. Human tissues contain CD141hi cross-presenting dendritic cells with functional homology to mouse CD103+ nonlymphoid dendritic cells. Immunity 2012, 37, 60–73. [Google Scholar] [CrossRef] [Green Version]
- Poulin, L.F.; Salio, M.; Griessinger, E.; Anjos-Afonso, F.; Craciun, L.; Chen, J.L.; Keller, A.M.; Joffre, O.; Zelenay, S.; Nye, E.; et al. Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8alpha+ dendritic cells. J. Exp. Med. 2010, 207, 1261–1271. [Google Scholar] [CrossRef]
- Jongbloed, S.L.; Kassianos, A.J.; McDonald, K.J.; Clark, G.J.; Ju, X.; Angel, C.E.; Chen, C.J.; Dunbar, P.R.; Wadley, R.B.; Jeet, V.; et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J. Exp. Med. 2010, 207, 1247–1260. [Google Scholar] [CrossRef] [Green Version]
- Bachem, A.; Guttler, S.; Hartung, E.; Ebstein, F.; Schaefer, M.; Tannert, A.; Salama, A.; Movassaghi, K.; Opitz, C.; Mages, H.W.; et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J. Exp. Med. 2010, 207, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Hemont, C.; Neel, A.; Heslan, M.; Braudeau, C.; Josien, R. Human blood mDC subsets exhibit distinct TLR repertoire and responsiveness. J. Leukoc. Biol. 2013, 93, 599–609. [Google Scholar] [CrossRef]
- Colletti, N.J.; Liu, H.; Gower, A.C.; Alekseyev, Y.O.; Arendt, C.W.; Shaw, M.H. TLR3 Signaling Promotes the Induction of Unique Human BDCA-3 Dendritic Cell Populations. Front. Immunol. 2016, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Lauterbach, H.; Bathke, B.; Gilles, S.; Traidl-Hoffmann, C.; Luber, C.A.; Fejer, G.; Freudenberg, M.A.; Davey, G.M.; Vremec, D.; Kallies, A.; et al. Mouse CD8alpha+ DCs and human BDCA3+ DCs are major producers of IFN-lambda in response to poly IC. J. Exp. Med. 2010, 207, 2703–2717. [Google Scholar] [CrossRef]
- Sittig, S.P.; Bakdash, G.; Weiden, J.; Skold, A.E.; Tel, J.; Figdor, C.G.; de Vries, I.J.; Schreibelt, G. A Comparative Study of the T Cell Stimulatory and Polarizing Capacity of Human Primary Blood Dendritic Cell Subsets. Mediat. Inflamm. 2016, 2016, 3605643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nizzoli, G.; Larghi, P.; Paroni, M.; Crosti, M.C.; Moro, M.; Neddermann, P.; Caprioli, F.; Pagani, M.; De Francesco, R.; Abrignani, S.; et al. IL-10 promotes homeostatic proliferation of human CD8+ memory T cells and, when produced by CD1c+ DCs, shapes naive CD8+ T-cell priming. Eur. J. Immunol. 2016, 46, 1622–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Blasio, S.; Wortel, I.M.; van Bladel, D.A.; de Vries, L.E.; Duiveman-de Boer, T.; Worah, K.; de Haas, N.; Buschow, S.I.; de Vries, I.J.; Figdor, C.G.; et al. Human CD1c+ DCs are critical cellular mediators of immune responses induced by immunogenic cell death. Oncoimmunology 2016, 5, e1192739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittag, D.; Proietto, A.I.; Loudovaris, T.; Mannering, S.I.; Vremec, D.; Shortman, K.; Wu, L.; Harrison, L.C. Human dendritic cell subsets from spleen and blood are similar in phenotype and function but modified by donor health status. J. Immunol. 2011, 186, 6207–6217. [Google Scholar] [CrossRef] [Green Version]
- Segura, E.; Durand, M.; Amigorena, S. Similar antigen cross-presentation capacity and phagocytic functions in all freshly isolated human lymphoid organ-resident dendritic cells. J. Exp. Med. 2013, 210, 1035–1047. [Google Scholar] [CrossRef] [Green Version]
- Nizzoli, G.; Krietsch, J.; Weick, A.; Steinfelder, S.; Facciotti, F.; Gruarin, P.; Bianco, A.; Steckel, B.; Moro, M.; Crosti, M.; et al. Human CD1c+ dendritic cells secrete high levels of IL-12 and potently prime cytotoxic T-cell responses. Blood 2013, 122, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Cohn, L.; Chatterjee, B.; Esselborn, F.; Smed-Sorensen, A.; Nakamura, N.; Chalouni, C.; Lee, B.C.; Vandlen, R.; Keler, T.; Lauer, P.; et al. Antigen delivery to early endosomes eliminates the superiority of human blood BDCA3+ dendritic cells at cross presentation. J. Exp. Med. 2013, 210, 1049–1063. [Google Scholar] [CrossRef] [Green Version]
- Bell, D.; Chomarat, P.; Broyles, D.; Netto, G.; Harb, G.M.; Lebecque, S.; Valladeau, J.; Davoust, J.; Palucka, K.A.; Banchereau, J. In breast carcinoma tissue, immature dendritic cells reside within the tumor, whereas mature dendritic cells are located in peritumoral areas. J. Exp. Med. 1999, 190, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Toriyama, K.; Wen, D.R.; Paul, E.; Cochran, A.J. Variations in the distribution, frequency, and phenotype of Langerhans cells during the evolution of malignant melanoma of the skin. J. Investig. Dermatol. 1993, 100, S269–S273. [Google Scholar] [CrossRef] [Green Version]
- Troy, A.J.; Summers, K.L.; Davidson, P.J.; Atkinson, C.H.; Hart, D.N. Minimal recruitment and activation of dendritic cells within renal cell carcinoma. Clin. Cancer Res. 1998, 4, 585–593. [Google Scholar] [PubMed]
- Zou, W.; Machelon, V.; Coulomb-L’Hermin, A.; Borvak, J.; Nome, F.; Isaeva, T.; Wei, S.; Krzysiek, R.; Durand-Gasselin, I.; Gordon, A.; et al. Stromal-derived factor-1 in human tumors recruits and alters the function of plasmacytoid precursor dendritic cells. Nat. Med. 2001, 7, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Almand, B.; Resser, J.R.; Lindman, B.; Nadaf, S.; Clark, J.I.; Kwon, E.D.; Carbone, D.P.; Gabrilovich, D.I. Clinical significance of defective dendritic cell differentiation in cancer. Clin. Cancer Res. 2000, 6, 1755–1766. [Google Scholar] [PubMed]
- Della Bella, S.; Gennaro, M.; Vaccari, M.; Ferraris, C.; Nicola, S.; Riva, A.; Clerici, M.; Greco, M.; Villa, M.L. Altered maturation of peripheral blood dendritic cells in patients with breast cancer. Br. J. Cancer 2003, 89, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Kalinski, P.; Edington, H.; Zeh, H.J.; Okada, H.; Butterfield, L.H.; Kirkwood, J.M.; Bartlett, D.L. Dendritic cells in cancer immunotherapy: Vaccines or autologous transplants? Immunol. Res. 2011, 50, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Amigorena, S.; Savina, A. Intracellular mechanisms of antigen cross presentation in dendritic cells. Curr. Opin. Immunol. 2010, 22, 109–117. [Google Scholar] [CrossRef]
- Segura, E.; Villadangos, J.A. Antigen presentation by dendritic cells in vivo. Curr. Opin. Immunol. 2009, 21, 105–110. [Google Scholar] [CrossRef]
- Shortman, K.; Heath, W.R. The CD8+ dendritic cell subset. Immunol. Rev. 2010, 234, 18–31. [Google Scholar] [CrossRef]
- Manfredi, A.A.; Capobianco, A.; Bianchi, M.E.; Rovere-Querini, P. Regulation of dendritic- and T-cell fate by injury-associated endogenous signals. Crit. Rev. Immunol. 2009, 29, 69–86. [Google Scholar] [CrossRef]
- Lopez-Albaitero, A.; Mailliard, R.; Hackman, T.; Andrade Filho, P.A.; Wang, X.; Gooding, W.; Ferrone, S.; Kalinski, P.; Ferris, R.L. Maturation pathways of dendritic cells determine TAP1 and TAP2 levels and cross-presenting function. J. Immunother. 2009, 32, 465. [Google Scholar] [CrossRef] [Green Version]
- De Vries, I.J.; Krooshoop, D.J.; Scharenborg, N.M.; Lesterhuis, W.J.; Diepstra, J.H.; Van Muijen, G.N.; Strijk, S.P.; Ruers, T.J.; Boerman, O.C.; Oyen, W.J.; et al. Effective migration of antigen-pulsed dendritic cells to lymph nodes in melanoma patients is determined by their maturation state. Cancer Res. 2003, 63, 12–17. [Google Scholar] [PubMed]
- De Vries, I.J.; Lesterhuis, W.J.; Scharenborg, N.M.; Engelen, L.P.; Ruiter, D.J.; Gerritsen, M.J.; Croockewit, S.; Britten, C.M.; Torensma, R.; Adema, G.J.; et al. Maturation of dendritic cells is a prerequisite for inducing immune responses in advanced melanoma patients. Clin. Cancer Res. 2003, 9, 5091–5100. [Google Scholar] [PubMed]
- Dhodapkar, M.V.; Steinman, R.M.; Sapp, M.; Desai, H.; Fossella, C.; Krasovsky, J.; Donahoe, S.M.; Dunbar, P.R.; Cerundolo, V.; Nixon, D.F.; et al. Rapid generation of broad T-cell immunity in humans after a single injection of mature dendritic cells. J. Clin. Investig. 1999, 104, 173–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adema, G.J.; de Vries, I.J.; Punt, C.J.; Figdor, C.G. Migration of dendritic cell based cancer vaccines: In vivo veritas? Curr. Opin. Immunol. 2005, 17, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Sapp, M.; Schuler, G.; Steinman, R.M.; Bhardwaj, N. Improved methods for the generation of dendritic cells from nonproliferating progenitors in human blood. J. Immunol. Methods 1996, 196, 121–135. [Google Scholar] [CrossRef]
- Reddy, A.; Sapp, M.; Feldman, M.; Subklewe, M.; Bhardwaj, N. A monocyte conditioned medium is more effective than defined cytokines in mediating the terminal maturation of human dendritic cells. Blood 1997, 90, 3640–3646. [Google Scholar] [CrossRef]
- Jonuleit, H.; Kuhn, U.; Muller, G.; Steinbrink, K.; Paragnik, L.; Schmitt, E.; Knop, J.; Enk, A.H. Pro-inflammatory cytokines and prostaglandins induce maturation of potent immunostimulatory dendritic cells under fetal calf serum-free conditions. Eur. J. Immunol. 1997, 27, 3135–3142. [Google Scholar] [CrossRef]
- Luft, T.; Jefford, M.; Luetjens, P.; Toy, T.; Hochrein, H.; Masterman, K.A.; Maliszewski, C.; Shortman, K.; Cebon, J.; Maraskovsky, E. Functionally distinct dendritic cell (DC) populations induced by physiologic stimuli: Prostaglandin E2 regulates the migratory capacity of specific DC subsets. Blood 2002, 100, 1362–1372. [Google Scholar] [CrossRef] [Green Version]
- Scandella, E.; Men, Y.; Gillessen, S.; Forster, R.; Groettrup, M. Prostaglandin E2 is a key factor for CCR7 surface expression and migration of monocyte-derived dendritic cells. Blood 2002, 100, 1354–1361. [Google Scholar] [CrossRef]
- Mailliard, R.B.; Wankowicz-Kalinska, A.; Cai, Q.; Wesa, A.; Hilkens, C.M.; Kapsenberg, M.L.; Kirkwood, J.M.; Storkus, W.J.; Kalinski, P. alpha-type-1 polarized dendritic cells: A novel immunization tool with optimized CTL-inducing activity. Cancer Res. 2004, 64, 5934–5937. [Google Scholar] [CrossRef] [Green Version]
- Muthuswamy, R.; Mueller-Berghaus, J.; Haberkorn, U.; Reinhart, T.A.; Schadendorf, D.; Kalinski, P. PGE2 transiently enhances DC expression of CCR7 but inhibits the ability of DCs to produce CCL19 and attract naive T cells. Blood 2010, 116, 1454–1459. [Google Scholar] [CrossRef] [PubMed]
- Zaccard, C.R.; Watkins, S.C.; Kalinski, P.; Fecek, R.J.; Yates, A.L.; Salter, R.D.; Ayyavoo, V.; Rinaldo, C.R.; Mailliard, R.B. CD40L induces functional tunneling nanotube networks exclusively in dendritic cells programmed by mediators of type 1 immunity. J. Immunol. 2015, 194, 1047–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, S.; Takahashi, H.K.; Takei, M.; Iwagaki, H.; Yoshino, T.; Tanaka, N.; Mori, S.; Nishibori, M. E-prostanoid (EP)2/EP4 receptor-dependent maturation of human monocyte-derived dendritic cells and induction of helper T2 polarization. J. Pharmacol. Exp. Ther. 2004, 309, 1213–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinski, P.; Schuitemaker, J.H.; Hilkens, C.M.; Kapsenberg, M.L. Prostaglandin E2 induces the final maturation of IL-12-deficient CD1a+CD83+ dendritic cells: The levels of IL-12 are determined during the final dendritic cell maturation and are resistant to further modulation. J. Immunol. 1998, 161, 2804–2809. [Google Scholar] [PubMed]
- Vieira, P.L.; de Jong, E.C.; Wierenga, E.A.; Kapsenberg, M.L.; Kalinski, P. Development of Th1-inducing capacity in myeloid dendritic cells requires environmental instruction. J. Immunol. 2000, 164, 4507–4512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinski, P.; Vieira, P.L.; Schuitemaker, J.H.; de Jong, E.C.; Kapsenberg, M.L. Prostaglandin E2 is a selective inducer of interleukin-12 p40 (IL-12p40) production and an inhibitor of bioactive IL-12p70 heterodimer. Blood 2001, 97, 3466–3469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthuswamy, R.; Urban, J.; Lee, J.J.; Reinhart, T.A.; Bartlett, D.; Kalinski, P. Ability of mature dendritic cells to interact with regulatory T cells is imprinted during maturation. Cancer Res. 2008, 68, 5972–5978. [Google Scholar] [CrossRef] [Green Version]
- Schadendorf, D.; Ugurel, S.; Schuler-Thurner, B.; Nestle, F.O.; Enk, A.; Brocker, E.B.; Grabbe, S.; Rittgen, W.; Edler, L.; Sucker, A.; et al. Dacarbazine (DTIC) versus vaccination with autologous peptide-pulsed dendritic cells (DC) in first-line treatment of patients with metastatic melanoma: A randomized phase III trial of the DC study group of the DeCOG. Ann. Oncol. 2006, 17, 563–570. [Google Scholar] [CrossRef]
- Macatangay, B.J.; Szajnik, M.E.; Whiteside, T.L.; Riddler, S.A.; Rinaldo, C.R. Regulatory T cell suppression of Gag-specific CD8 T cell polyfunctional response after therapeutic vaccination of HIV-1-infected patients on ART. PLoS ONE 2010, 5, e9852. [Google Scholar] [CrossRef] [Green Version]
- Wesa, A.; Kalinski, P.; Kirkwood, J.M.; Tatsumi, T.; Storkus, W.J. Polarized type-1 dendritic cells (DC1) producing high levels of IL-12 family members rescue patient TH1-type antimelanoma CD4+ T cell responses in vitro. J. Immunother. 2007, 30, 75–82. [Google Scholar] [CrossRef]
- Gustafsson, K.; Junevik, K.; Werlenius, O.; Holmgren, S.; Karlsson-Parra, A.; Andersson, P.O. Tumour-loaded alpha-type 1-polarized dendritic cells from patients with chronic lymphocytic leukaemia produce a superior NK-, NKT- and CD8+ T cell-attracting chemokine profile. Scand. J. Immunol. 2011, 74, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Mailliard, R.B.; Smith, K.N.; Fecek, R.J.; Rappocciolo, G.; Nascimento, E.J.; Marques, E.T.; Watkins, S.C.; Mullins, J.I.; Rinaldo, C.R. Selective induction of CTL helper rather than killer activity by natural epitope variants promotes dendritic cell-mediated HIV-1 dissemination. J. Immunol. 2013, 191, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Mayordomo, J.I.; Tjandrawan, T.; DeLeo, A.B.; Clarke, M.R.; Lotze, M.T.; Storkus, W.J. Therapy of murine tumors with tumor peptide-pulsed dendritic cells: Dependence on T cells, B7 costimulation, and T helper cell 1-associated cytokines. J. Exp. Med. 1996, 183, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Robbins, P.D.; Storkus, W.J.; Clarke, M.R.; Maeurer, M.J.; Campbell, R.L.; Davis, C.G.; Tahara, H.; Schreiber, R.D.; Lotze, M.T. Interleukin-12 and B7.1 co-stimulation cooperate in the induction of effective antitumor immunity and therapy of established tumors. Eur. J. Immunol. 1996, 26, 1335–1341. [Google Scholar] [CrossRef]
- Furumoto, K.; Arii, S.; Yamasaki, S.; Mizumoto, M.; Mori, A.; Inoue, N.; Isobe, N.; Imamura, M. Spleen-derived dendritic cells engineered to enhance interleukin-12 production elicit therapeutic antitumor immune responses. Int. J. Cancer 2000, 87, 665–672. [Google Scholar] [CrossRef]
- Furumoto, K.; Mori, A.; Yamasaki, S.; Inoue, N.; Yang, W.; Nakau, M.; Yasuda, S.; Arii, S.; Imamura, M. Interleukin-12-gene transduction makes DCs from tumor-bearing mice an effective inducer of tumor-specific immunity in a peritoneal dissemination model. Immunol. Lett. 2002, 83, 13–20. [Google Scholar] [CrossRef]
- Nishioka, Y.; Hirao, M.; Robbins, P.D.; Lotze, M.T.; Tahara, H. Induction of systemic and therapeutic antitumor immunity using intratumoral injection of dendritic cells genetically modified to express interleukin 12. Cancer Res. 1999, 59, 4035–4041. [Google Scholar]
- Okada, H.; Kalinski, P.; Ueda, R.; Hoji, A.; Kohanbash, G.; Donegan, T.E.; Mintz, A.H.; Engh, J.A.; Bartlett, D.L.; Brown, C.K.; et al. Induction of CD8+ T-cell responses against novel glioma-associated antigen peptides and clinical activity by vaccinations with {alpha}-type 1 polarized dendritic cells and polyinosinic-polycytidylic acid stabilized by lysine and carboxymethylcellulose in patients with recurrent malignant glioma. J. Clin. Oncol. 2011, 29, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Redlinger, R.E., Jr.; Mailliard, R.B.; Barksdale, E.M., Jr. Advanced neuroblastoma impairs dendritic cell function in adoptive immunotherapy. J. Pediatr. Surg. 2003, 38, 857–862. [Google Scholar] [CrossRef]
- Satoh, Y.; Esche, C.; Gambotto, A.; Shurin, G.V.; Yurkovetsky, Z.R.; Robbins, P.D.; Watkins, S.C.; Todo, S.; Herberman, R.B.; Lotze, M.T.; et al. Local administration of IL-12-transfected dendritic cells induces antitumor immune responses to colon adenocarcinoma in the liver in mice. J. Exp. Ther. Oncol. 2002, 2, 337–349. [Google Scholar] [CrossRef]
- Shimizu, T.; Berhanu, A.; Redlinger, R.E., Jr.; Watkins, S.; Lotze, M.T.; Barksdale, E.M., Jr. Interleukin-12 transduced dendritic cells induce regression of established murine neuroblastoma. J. Pediatr. Surg. 2001, 36, 1285–1292. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, R.; Zullo, S.A.; Ramsey, J.; Yajima, N.; Tsuchiya, N.; Tanaka, R.; Blaese, M.; Xanthopoulos, K.G. Marked enhancement of antitumor immune responses in mouse brain tumor models by genetically modified dendritic cells producing Semliki Forest virus-mediated interleukin-12. J. Neurosurg. 2002, 97, 611–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zeng, G.; Wilkes, D.S.; Reed, G.E.; McGarry, R.C.; Eble, J.N.; Cheng, L. Dendritic cells transfected with interleukin-12 and pulsed with tumor extract inhibit growth of murine prostatic carcinoma in vivo. Prostate 2003, 55, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Kalinski, P.; Schuitemaker, J.H.; Hilkens, C.M.; Wierenga, E.A.; Kapsenberg, M.L. Final maturation of dendritic cells is associated with impaired responsiveness to IFN-gamma and to bacterial IL-12 inducers: Decreased ability of mature dendritic cells to produce IL-12 during the interaction with Th cells. J. Immunol. 1999, 162, 3231–3236. [Google Scholar] [PubMed]
- Langenkamp, A.; Messi, M.; Lanzavecchia, A.; Sallusto, F. Kinetics of dendritic cell activation: Impact on priming of TH1, TH2 and nonpolarized T cells. Nat. Immunol. 2000, 1, 311–316. [Google Scholar] [CrossRef]
- Kalinski, P.; Giermasz, A.; Nakamura, Y.; Basse, P.; Storkus, W.J.; Kirkwood, J.M.; Mailliard, R.B. Helper role of NK cells during the induction of anticancer responses by dendritic cells. Mol. Immunol. 2005, 42, 535–539. [Google Scholar] [CrossRef]
- Kalinski, P.; Mailliard, R.B.; Giermasz, A.; Zeh, H.J.; Basse, P.; Bartlett, D.L.; Kirkwood, J.M.; Lotze, M.T.; Herberman, R.B. Natural killer-dendritic cell cross-talk in cancer immunotherapy. Expert Opin. Biol. Ther. 2005, 5, 1303–1315. [Google Scholar] [CrossRef]
- Mailliard, R.B.; Egawa, S.; Cai, Q.; Kalinska, A.; Bykovskaya, S.N.; Lotze, M.T.; Kapsenberg, M.L.; Storkus, W.J.; Kalinski, P. Complementary dendritic cell-activating function of CD8+ and CD4+ T cells: Helper role of CD8+ T cells in the development of T helper type 1 responses. J. Exp. Med. 2002, 195, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Mailliard, R.B.; Son, Y.I.; Redlinger, R.; Coates, P.T.; Giermasz, A.; Morel, P.A.; Storkus, W.J.; Kalinski, P. Dendritic cells mediate NK cell help for Th1 and CTL responses: Two-signal requirement for the induction of NK cell helper function. J. Immunol. 2003, 171, 2366–2373. [Google Scholar] [CrossRef]
- Xu, S.; Koski, G.K.; Faries, M.; Bedrosian, I.; Mick, R.; Maeurer, M.; Cheever, M.A.; Cohen, P.A.; Czerniecki, B.J. Rapid high efficiency sensitization of CD8+ T cells to tumor antigens by dendritic cells leads to enhanced functional avidity and direct tumor recognition through an IL-12-dependent mechanism. J. Immunol. 2003, 171, 2251–2261. [Google Scholar] [CrossRef] [Green Version]
- Kalinski, P.; Nakamura, Y.; Watchmaker, P.; Giermasz, A.; Muthuswamy, R.; Mailliard, R.B. Helper roles of NK and CD8+ T cells in the induction of tumor immunity. Polarized dendritic cells as cancer vaccines. Immunol. Res. 2006, 36, 137–146. [Google Scholar] [CrossRef]
- Ten Brinke, A.; Karsten, M.L.; Dieker, M.C.; Zwaginga, J.J.; van Ham, S.M. The clinical grade maturation cocktail monophosphoryl lipid A plus IFNgamma generates monocyte-derived dendritic cells with the capacity to migrate and induce Th1 polarization. Vaccine 2007, 25, 7145–7152. [Google Scholar] [CrossRef] [PubMed]
- Camporeale, A.; Boni, A.; Iezzi, G.; Degl’Innocenti, E.; Grioni, M.; Mondino, A.; Bellone, M. Critical impact of the kinetics of dendritic cells activation on the in vivo induction of tumor-specific T lymphocytes. Cancer Res. 2003, 63, 3688–3694. [Google Scholar] [PubMed]
- Watchmaker, P.B.; Berk, E.; Muthuswamy, R.; Mailliard, R.B.; Urban, J.A.; Kirkwood, J.M.; Kalinski, P. Independent regulation of chemokine responsiveness and cytolytic function versus CD8+ T cell expansion by dendritic cells. J. Immunol. 2010, 184, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Barratt-Boyes, S.M.; Figdor, C.G. Current issues in delivering DCs for immunotherapy. Cytotherapy 2004, 6, 105–110. [Google Scholar] [CrossRef]
- Jusforgues-Saklani, H.; Uhl, M.; Blachere, N.; Lemaitre, F.; Lantz, O.; Bousso, P.; Braun, D.; Moon, J.J.; Albert, M.L. Antigen persistence is required for dendritic cell licensing and CD8+ T cell cross-priming. J. Immunol. 2008, 181, 3067–3076. [Google Scholar] [CrossRef] [Green Version]
- Archin, N.M.; Liberty, A.L.; Kashuba, A.D.; Choudhary, S.K.; Kuruc, J.D.; Crooks, A.M.; Parker, D.C.; Anderson, E.M.; Kearney, M.F.; Strain, M.C.; et al. Administration of vorinostat disrupts HIV-1 latency in patients on antiretroviral therapy. Nature 2012, 487, 482–485. [Google Scholar] [CrossRef] [Green Version]
- Archin, N.M.; Kirchherr, J.L.; Sung, J.A.; Clutton, G.; Sholtis, K.; Xu, Y.; Allard, B.; Stuelke, E.; Kashuba, A.D.; Kuruc, J.D.; et al. Interval dosing with the HDAC inhibitor vorinostat effectively reverses HIV latency. J. Clin. Investig. 2017, 127, 3126–3135. [Google Scholar] [CrossRef] [Green Version]
- Elliott, J.H.; McMahon, J.H.; Chang, C.C.; Lee, S.A.; Hartogensis, W.; Bumpus, N.; Savic, R.; Roney, J.; Hoh, R.; Solomon, A.; et al. Short-term administration of disulfiram for reversal of latent HIV infection: A phase 2 dose-escalation study. Lancet HIV 2015, 2, e520–e529. [Google Scholar] [CrossRef] [Green Version]
- Elliott, J.H.; Wightman, F.; Solomon, A.; Ghneim, K.; Ahlers, J.; Cameron, M.J.; Smith, M.Z.; Spelman, T.; McMahon, J.; Velayudham, P.; et al. Activation of HIV transcription with short-course vorinostat in HIV-infected patients on suppressive antiretroviral therapy. PLoS Pathog. 2014, 10, e1004473. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, C.; Serrano-Villar, S.; Madrid-Elena, N.; Perez-Elias, M.J.; Martin, M.E.; Barbas, C.; Ruiperez, J.; Munoz, E.; Munoz-Fernandez, M.A.; Castor, T.; et al. Bryostatin-1 for latent virus reactivation in HIV-infected patients on antiretroviral therapy. Aids 2016, 30, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.A.; Tolstrup, M.; Brinkmann, C.R.; Olesen, R.; Erikstrup, C.; Solomon, A.; Winckelmann, A.; Palmer, S.; Dinarello, C.; Buzon, M.; et al. Panobinostat, a histone deacetylase inhibitor, for latent-virus reactivation in HIV-infected patients on suppressive antiretroviral therapy: A phase 1/2, single group, clinical trial. Lancet HIV 2014, 1, e13–e21. [Google Scholar] [CrossRef]
- Sogaard, O.S.; Graversen, M.E.; Leth, S.; Olesen, R.; Brinkmann, C.R.; Nissen, S.K.; Kjaer, A.S.; Schleimann, M.H.; Denton, P.W.; Hey-Cunningham, W.J.; et al. The Depsipeptide Romidepsin Reverses HIV-1 Latency In Vivo. PLoS Pathog. 2015, 11, e1005142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak, A.M.; Planelles, V. HIV-1 Eradication: Early Trials (and Tribulations). Trends Mol. Med. 2016, 22, 10–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.A.; Tolstrup, M.; Sogaard, O.S. Reversal of Latency as Part of a Cure for HIV-1. Trends Microbiol. 2016, 24, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Bauer, D.E.; Tuttleton, S.E.; Lewin, S.; Gettie, A.; Blanchard, J.; Irwin, C.E.; Safrit, J.T.; Mittler, J.; Weinberger, L.; et al. Dramatic rise in plasma viremia after CD8+ T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 1999, 189, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, J.E.; Kuroda, M.J.; Santra, S.; Sasseville, V.G.; Simon, M.A.; Lifton, M.A.; Racz, P.; Tenner-Racz, K.; Dalesandro, M.; Scallon, B.J.; et al. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science 1999, 283, 857–860. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, A.; Hayes, T.L.; Bosinger, S.E.; Lawson, B.O.; Vanderford, T.; Schmitz, J.E.; Paiardini, M.; Betts, M.; Chahroudi, A.; Estes, J.D.; et al. Differential Impact of In Vivo CD8+ T Lymphocyte Depletion in Controller versus Progressor Simian Immunodeficiency Virus-Infected Macaques. J. Virol. 2015, 89, 8677–8686. [Google Scholar] [CrossRef] [Green Version]
- Cartwright, E.K.; Spicer, L.; Smith, S.A.; Lee, D.; Fast, R.; Paganini, S.; Lawson, B.O.; Nega, M.; Easley, K.; Schmitz, J.E.; et al. CD8+ Lymphocytes Are Required for Maintaining Viral Suppression in SIV-Infected Macaques Treated with Short-Term Antiretroviral Therapy. Immunity 2016, 45, 656–668. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Cartwright, E.K.; Silvestri, G.; Perelson, A.S. CD8+ lymphocyte control of SIV infection during antiretroviral therapy. PLoS Pathog. 2018, 14, e1007350. [Google Scholar] [CrossRef] [Green Version]
- Borrow, P.; Lewicki, H.; Hahn, B.H.; Shaw, G.M.; Oldstone, M.B. Virus-specific CD8+ cytotoxic T-lymphocyte activity associated with control of viremia in primary human immunodeficiency virus type 1 infection. J. Virol. 1994, 68, 6103–6110. [Google Scholar] [PubMed]
- Koup, R.A.; Safrit, J.T.; Cao, Y.; Andrews, C.A.; McLeod, G.; Borkowsky, W.; Farthing, C.; Ho, D.D. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus type 1 syndrome. J. Virol. 1994, 68, 4650–4655. [Google Scholar] [PubMed]
- Musey, L.; Hughes, J.; Schacker, T.; Shea, T.; Corey, L.; McElrath, M.J. Cytotoxic-T-cell responses, viral load, and disease progression in early human immunodeficiency virus type 1 infection. N. Engl. J. Med. 1997, 337, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Siliciano, R.F. Targeting the Latent Reservoir for HIV-1. Immunity 2018, 48, 872–895. [Google Scholar] [CrossRef] [Green Version]
- Borrow, P.; Lewicki, H.; Wei, X.; Horwitz, M.S.; Peffer, N.; Meyers, H.; Nelson, J.A.; Gairin, J.E.; Hahn, B.H.; Oldstone, M.B.; et al. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med. 1997, 3, 205–211. [Google Scholar] [CrossRef]
- Goonetilleke, N.; Liu, M.K.; Salazar-Gonzalez, J.F.; Ferrari, G.; Giorgi, E.; Ganusov, V.V.; Keele, B.F.; Learn, G.H.; Turnbull, E.L.; Salazar, M.G.; et al. The first T cell response to transmitted/founder virus contributes to the control of acute viremia in HIV-1 infection. J. Exp. Med. 2009, 206, 1253–1272. [Google Scholar] [CrossRef]
- Price, D.A.; Goulder, P.J.; Klenerman, P.; Sewell, A.K.; Easterbrook, P.J.; Troop, M.; Bangham, C.R.; Phillips, R.E. Positive selection of HIV-1 cytotoxic T lymphocyte escape variants during primary infection. Proc. Natl. Acad. Sci. USA 1997, 94, 1890–1895. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.; McMichael, A. The T-cell response to HIV. Cold Spring Harb. Perspect. Med. 2012, 2, a007054. [Google Scholar] [CrossRef] [Green Version]
- Deng, K.; Pertea, M.; Rongvaux, A.; Wang, L.; Durand, C.M.; Ghiaur, G.; Lai, J.; McHugh, H.L.; Hao, H.; Zhang, H.; et al. Broad CTL response is required to clear latent HIV-1 due to dominance of escape mutations. Nature 2015, 517, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.N.; Mailliard, R.B.; Piazza, P.A.; Fischer, W.; Korber, B.T.; Fecek, R.J.; Ratner, D.; Gupta, P.; Mullins, J.I.; Rinaldo, C.R. Effective Cytotoxic T Lymphocyte Targeting of Persistent HIV-1 during Antiretroviral Therapy Requires Priming of Naive CD8+ T Cells. Mbio 2016, 7, e00473-16, Erratum in: MBio. 2016;7(4). pii: e01012-16. doi: 10.1128/mBio.01012-16. [Google Scholar] [CrossRef] [Green Version]
- Qin, K.; Boppana, S.; Du, V.Y.; Carlson, J.M.; Yue, L.; Dilernia, D.A.; Hunter, E.; Mailliard, R.B.; Mallal, S.A.; Bansal, A.; et al. CD8 T cells targeting adapted epitopes in chronic HIV infection promote dendritic cell maturation and CD4 T cell trans-infection. PLoS Pathog. 2019, 15, e1007970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theiler, J.; Korber, B. Graph-based optimization of epitope coverage for vaccine antigen design. Stat. Med. 2018, 37, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaiha, G.D.; Rossin, E.J.; Urbach, J.; Landeros, C.; Collins, D.R.; Nwonu, C.; Muzhingi, I.; Anahtar, M.N.; Waring, O.M.; Piechocka-Trocha, A.; et al. Structural topology defines protective CD8+ T cell epitopes in the HIV proteome. Science 2019, 364, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Bucks, C.M.; Norton, J.A.; Boesteanu, A.C.; Mueller, Y.M.; Katsikis, P.D. Chronic antigen stimulation alone is sufficient to drive CD8+ T cell exhaustion. J. Immunol. 2009, 182, 6697–6708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockerham, L.R.; Jain, V.; Sinclair, E.; Glidden, D.V.; Hartogenesis, W.; Hatano, H.; Hunt, P.W.; Martin, J.N.; Pilcher, C.D.; Sekaly, R.; et al. Programmed death-1 expression on CD4+ and CD8+ T cells in treated and untreated HIV disease. Aids 2014, 28, 1749–1758. [Google Scholar] [CrossRef] [Green Version]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.J.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350–354. [Google Scholar] [CrossRef]
- Streeck, H.; Brumme, Z.L.; Anastario, M.; Cohen, K.W.; Jolin, J.S.; Meier, A.; Brumme, C.J.; Rosenberg, E.S.; Alter, G.; Allen, T.M.; et al. Antigen load and viral sequence diversification determine the functional profile of HIV-1-specific CD8+ T cells. PLoS Med. 2008, 5, e100. [Google Scholar] [CrossRef]
- Kalams, S.A.; Buchbinder, S.P.; Rosenberg, E.S.; Billingsley, J.M.; Colbert, D.S.; Jones, N.G.; Shea, A.K.; Trocha, A.K.; Walker, B.D. Association between virus-specific cytotoxic T-lymphocyte and helper responses in human immunodeficiency virus type 1 infection. J. Virol. 1999, 73, 6715–6720. [Google Scholar]
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T cell exhaustion during persistent viral infections. Virology 2015, 479, 180–193. [Google Scholar] [CrossRef]
- Trautmann, L.; Janbazian, L.; Chomont, N.; Said, E.A.; Gimmig, S.; Bessette, B.; Boulassel, M.R.; Delwart, E.; Sepulveda, H.; Balderas, R.S.; et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat. Med. 2006, 12, 1198–1202. [Google Scholar] [CrossRef]
- Petrovas, C.; Casazza, J.P.; Brenchley, J.M.; Price, D.A.; Gostick, E.; Adams, W.C.; Precopio, M.L.; Schacker, T.; Roederer, M.; Douek, D.C.; et al. PD-1 is a regulator of virus-specific CD8+ T cell survival in HIV infection. J. Exp. Med. 2006, 203, 2281–2292. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Ha, S.J.; Kaech, S.M.; Haining, W.N.; Sarkar, S.; Kalia, V.; Subramaniam, S.; Blattman, J.N.; Barber, D.L.; Ahmed, R. Molecular signature of CD8+ T cell exhaustion during chronic viral infection. Immunity 2007, 27, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.B.; Ndhlovu, L.C.; Barbour, J.D.; Sheth, P.M.; Jha, A.R.; Long, B.R.; Wong, J.C.; Satkunarajah, M.; Schweneker, M.; Chapman, J.M.; et al. Tim-3 expression defines a novel population of dysfunctional T cells with highly elevated frequencies in progressive HIV-1 infection. J. Exp. Med. 2008, 205, 2763–2779. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, S.D.; Shin, H.; Haining, W.N.; Zou, T.; Workman, C.J.; Polley, A.; Betts, M.R.; Freeman, G.J.; Vignali, D.A.; Wherry, E.J. Coregulation of CD8+ T cell exhaustion by multiple inhibitory receptors during chronic viral infection. Nat. Immunol. 2009, 10, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Petrovas, C.; Ferrando-Martinez, S.; Gerner, M.Y.; Casazza, J.P.; Pegu, A.; Deleage, C.; Cooper, A.; Hataye, J.; Andrews, S.; Ambrozak, D.; et al. Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies. Sci. Transl. Med. 2017, 9, eaag2285. [Google Scholar] [CrossRef] [Green Version]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 2006, 439, 682–687. [Google Scholar] [CrossRef]
- Blackburn, S.D.; Shin, H.; Freeman, G.J.; Wherry, E.J. Selective expansion of a subset of exhausted CD8 T cells by alphaPD-L1 blockade. Proc. Natl. Acad. Sci. USA 2008, 105, 15016–15021. [Google Scholar] [CrossRef] [Green Version]
- Gay, C.L.; Bosch, R.J.; Ritz, J.; Hataye, J.M.; Aga, E.; Tressler, R.L.; Mason, S.W.; Hwang, C.K.; Grasela, D.M.; Ray, N.; et al. Clinical Trial of the Anti-PD-L1 Antibody BMS-936559 in HIV-1 Infected Participants on Suppressive Antiretroviral Therapy. J. Infect. Dis. 2017, 215, 1725–1733. [Google Scholar] [CrossRef] [Green Version]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef]
- Fromentin, R.; Bakeman, W.; Lawani, M.B.; Khoury, G.; Hartogensis, W.; DaFonseca, S.; Killian, M.; Epling, L.; Hoh, R.; Sinclair, E.; et al. CD4+ T Cells Expressing PD-1, TIGIT and LAG-3 Contribute to HIV Persistence during ART. PLoS Pathog. 2016, 12, e1005761. [Google Scholar] [CrossRef] [PubMed]
- Bruner, K.M.; Murray, A.J.; Pollack, R.A.; Soliman, M.G.; Laskey, S.B.; Capoferri, A.A.; Lai, J.; Strain, M.C.; Lada, S.M.; Hoh, R.; et al. Defective proviruses rapidly accumulate during acute HIV-1 infection. Nat. Med. 2016, 22, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- Pollack, R.A.; Jones, R.B.; Pertea, M.; Bruner, K.M.; Martin, A.R.; Thomas, A.S.; Capoferri, A.A.; Beg, S.A.; Huang, S.H.; Karandish, S.; et al. Defective HIV-1 Proviruses Are Expressed and Can Be Recognized by Cytotoxic T Lymphocytes, which Shape the Proviral Landscape. Cell Host Microbe 2017, 21, 494–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.C.; Shan, L.; Hosmane, N.N.; Wang, J.; Laskey, S.B.; Rosenbloom, D.I.; Lai, J.; Blankson, J.N.; Siliciano, J.D.; Siliciano, R.F. Replication-competent noninduced proviruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013, 155, 540–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamichi, H.; Dewar, R.L.; Adelsberger, J.W.; Rehm, C.A.; O’Doherty, U.; Paxinos, E.E.; Fauci, A.S.; Lane, H.C. Defective HIV-1 proviruses produce novel protein-coding RNA species in HIV-infected patients on combination antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2016, 113, 8783–8788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.H.; Ren, Y.; Thomas, A.S.; Chan, D.; Mueller, S.; Ward, A.R.; Patel, S.; Bollard, C.M.; Cruz, C.R.; Karandish, S.; et al. Latent HIV reservoirs exhibit inherent resistance to elimination by CD8+ T cells. J. Clin. Investig. 2018, 128, 876–889. [Google Scholar] [CrossRef]
- Maldarelli, F.; Wu, X.; Su, L.; Simonetti, F.R.; Shao, W.; Hill, S.; Spindler, J.; Ferris, A.L.; Mellors, J.W.; Kearney, M.F.; et al. HIV latency. Specific HIV integration sites are linked to clonal expansion and persistence of infected cells. Science 2014, 345, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.A.; McLaughlin, S.; Garg, K.; Cheung, C.Y.; Larsen, B.B.; Styrchak, S.; Huang, H.C.; Edlefsen, P.T.; Mullins, J.I.; Frenkel, L.M. HIV latency. Proliferation of cells with HIV integrated into cancer genes contributes to persistent infection. Science 2014, 345, 570–573. [Google Scholar] [CrossRef] [Green Version]
- Bui, J.K.; Sobolewski, M.D.; Keele, B.F.; Spindler, J.; Musick, A.; Wiegand, A.; Luke, B.T.; Shao, W.; Hughes, S.H.; Coffin, J.M.; et al. Proviruses with identical sequences comprise a large fraction of the replication-competent HIV reservoir. PLoS Pathog. 2017, 13, e1006283. [Google Scholar] [CrossRef]
- Hosmane, N.N.; Kwon, K.J.; Bruner, K.M.; Capoferri, A.A.; Beg, S.; Rosenbloom, D.I.; Keele, B.F.; Ho, Y.C.; Siliciano, J.D.; Siliciano, R.F. Proliferation of latently infected CD4+ T cells carrying replication-competent HIV-1: Potential role in latent reservoir dynamics. J. Exp. Med. 2017, 214, 959–972. [Google Scholar] [CrossRef]
- Cohn, L.B.; Silva, I.T.; Oliveira, T.Y.; Rosales, R.A.; Parrish, E.H.; Learn, G.H.; Hahn, B.H.; Czartoski, J.L.; McElrath, M.J.; Lehmann, C.; et al. HIV-1 integration landscape during latent and active infection. Cell 2015, 160, 420–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukazawa, Y.; Lum, R.; Okoye, A.A.; Park, H.; Matsuda, K.; Bae, J.Y.; Hagen, S.I.; Shoemaker, R.; Deleage, C.; Lucero, C.; et al. B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers. Nat. Med. 2015, 21, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amet, T.; Son, Y.M.; Jiang, L.; Cheon, I.S.; Huang, S.; Gupta, S.K.; Dent, A.L.; Montaner, L.J.; Yu, Q.; Sun, J. BCL6 represses antiviral resistance in follicular T helper cells. J. Leukoc. Biol. 2017, 102, 527–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohler, S.L.; Pham, M.N.; Folkvord, J.M.; Arends, T.; Miller, S.M.; Miles, B.; Meditz, A.L.; McCarter, M.; Levy, D.N.; Connick, E. Germinal Center T Follicular Helper Cells Are Highly Permissive to HIV-1 and Alter Their Phenotype during Virus Replication. J. Immunol. 2016, 196, 2711–2722. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.M.; Miles, B.; Guo, K.; Folkvord, J.; Meditz, A.L.; McCarter, M.D.; Levy, D.N.; MaWhinney, S.; Santiago, M.L.; Connick, E. Follicular Regulatory T Cells Are Highly Permissive to R5-Tropic HIV-1. J. Virol. 2017, 91, e00430-17. [Google Scholar] [CrossRef] [Green Version]
- Connick, E.; Mattila, T.; Folkvord, J.M.; Schlichtemeier, R.; Meditz, A.L.; Ray, M.G.; McCarter, M.D.; Mawhinney, S.; Hage, A.; White, C.; et al. CTL fail to accumulate at sites of HIV-1 replication in lymphoid tissue. J. Immunol. 2007, 178, 6975–6983. [Google Scholar] [CrossRef]
- Folkvord, J.M.; Armon, C.; Connick, E. Lymphoid follicles are sites of heightened human immunodeficiency virus type 1 (HIV-1) replication and reduced antiretroviral effector mechanisms. AIDS Res. Hum. Retrovir. 2005, 21, 363–370. [Google Scholar] [CrossRef]
- Perreau, M.; Savoye, A.L.; De Crignis, E.; Corpataux, J.M.; Cubas, R.; Haddad, E.K.; De Leval, L.; Graziosi, C.; Pantaleo, G. Follicular helper T cells serve as the major CD4 T cell compartment for HIV-1 infection, replication, and production. J. Exp. Med. 2013, 210, 143–156. [Google Scholar] [CrossRef]
- Hufert, F.T.; van Lunzen, J.; Janossy, G.; Bertram, S.; Schmitz, J.; Haller, O.; Racz, P.; von Laer, D. Germinal centre CD4+ T cells are an important site of HIV replication in vivo. Aids 1997, 11, 849–857. [Google Scholar] [CrossRef]
- Leong, Y.A.; Chen, Y.; Ong, H.S.; Wu, D.; Man, K.; Deleage, C.; Minnich, M.; Meckiff, B.J.; Wei, Y.; Hou, Z.; et al. CXCR5+ follicular cytotoxic T cells control viral infection in B cell follicles. Nat. Immunol. 2016, 17, 1187–1196. [Google Scholar] [CrossRef]
- Im, S.J.; Hashimoto, M.; Gerner, M.Y.; Lee, J.; Kissick, H.T.; Burger, M.C.; Shan, Q.; Hale, J.S.; Lee, J.; Nasti, T.H.; et al. Defining CD8+ T cells that provide the proliferative burst after PD-1 therapy. Nature 2016, 537, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Quigley, M.F.; Gonzalez, V.D.; Granath, A.; Andersson, J.; Sandberg, J.K. CXCR5+ CCR7− CD8 T cells are early effector memory cells that infiltrate tonsil B cell follicles. Eur. J. Immunol. 2007, 37, 3352–3362. [Google Scholar] [CrossRef] [PubMed]
- Bronnimann, M.P.; Skinner, P.J.; Connick, E. The B-Cell Follicle in HIV Infection: Barrier to a Cure. Front. Immunol. 2018, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Folkvord, J.M.; Rakasz, E.G.; Abdelaal, H.M.; Wagstaff, R.K.; Kovacs, K.J.; Kim, H.O.; Sawahata, R.; MaWhinney, S.; Masopust, D.; et al. Simian Immunodeficiency Virus-Producing Cells in Follicles Are Partially Suppressed by CD8+ Cells In Vivo. J. Virol. 2016, 90, 11168–11180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, V.I.; Deleage, C.; Trivett, M.T.; Jain, S.; Coren, L.V.; Breed, M.W.; Kramer, J.A.; Thomas, J.A.; Estes, J.D.; Lifson, J.D.; et al. CXCR5-Dependent Entry of CD8 T Cells into Rhesus Macaque B-Cell Follicles Achieved through T-Cell Engineering. J. Virol. 2017, 91, e02507-16. [Google Scholar] [CrossRef] [Green Version]
- Mylvaganam, G.H.; Rios, D.; Abdelaal, H.M.; Iyer, S.; Tharp, G.; Mavigner, M.; Hicks, S.; Chahroudi, A.; Ahmed, R.; Bosinger, S.E.; et al. Dynamics of SIV-specific CXCR5+ CD8 T cells during chronic SIV infection. Proc. Natl. Acad. Sci. USA 2017, 114, 1976–1981. [Google Scholar] [CrossRef] [Green Version]
- Pegu, A.; Asokan, M.; Wu, L.; Wang, K.; Hataye, J.; Casazza, J.P.; Guo, X.; Shi, W.; Georgiev, I.; Zhou, T.; et al. Activation and lysis of human CD4 cells latently infected with HIV-1. Nat. Commun. 2015, 6, 8447. [Google Scholar] [CrossRef]
- Deeks, S.G. HIV: Shock and kill. Nature 2012, 487, 439–440. [Google Scholar] [CrossRef]
- Shan, L.; Deng, K.; Gao, H.; Xing, S.; Capoferri, A.A.; Durand, C.M.; Rabi, S.A.; Laird, G.M.; Kim, M.; Hosmane, N.N.; et al. Transcriptional Reprogramming during Effector-to-Memory Transition Renders CD4+ T Cells Permissive for Latent HIV-1 Infection. Immunity 2017, 47, 766–775. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Anderson, J.L.; Lewin, S.R. Getting the “Kill” into “Shock and Kill”: Strategies to Eliminate Latent HIV. Cell Host Microbe 2018, 23, 14–26. [Google Scholar] [CrossRef]
- Spivak, A.M.; Planelles, V. Novel Latency Reversal Agents for HIV-1 Cure. Annu. Rev. Med. 2018, 69, 421–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, T.A.; Tolstrup, M.; Moller, H.J.; Brinkmann, C.R.; Olesen, R.; Erikstrup, C.; Laursen, A.L.; Ostergaard, L.; Sogaard, O.S. Activation of latent human immunodeficiency virus by the histone deacetylase inhibitor panobinostat: A pilot study to assess effects on the central nervous system. Open Forum Infect. Dis. 2015, 2, ofv037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkosky, J.; Sullivan, J.; Xu, Y.; Souder, E.; Hamer, D.H.; Pomerantz, R.J. Expression of latent HAART-persistent HIV type 1 induced by novel cellular activating agents. AIDS Res. Hum. Retrovir. 2004, 20, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.A.; Chen, L.F.; Kwon, H.; Fenard, D.; Bisgrove, D.; Verdin, E.; Greene, W.C. Prostratin antagonizes HIV latency by activating NF-kappaB. J. Biol. Chem. 2004, 279, 42008–42017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullen, C.K.; Laird, G.M.; Durand, C.M.; Siliciano, J.D.; Siliciano, R.F. New ex vivo approaches distinguish effective and ineffective single agents for reversing HIV-1 latency in vivo. Nat. Med. 2014, 20, 425–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clutton, G.; Xu, Y.; Baldoni, P.L.; Mollan, K.R.; Kirchherr, J.; Newhard, W.; Cox, K.; Kuruc, J.D.; Kashuba, A.; Barnard, R.; et al. The differential short- and long-term effects of HIV-1 latency-reversing agents on T cell function. Sci. Rep. 2016, 6, 30749. [Google Scholar] [CrossRef]
- Lam, A.P.; Sparano, J.A.; Vinciguerra, V.; Ocean, A.J.; Christos, P.; Hochster, H.; Camacho, F.; Goel, S.; Mani, S.; Kaubisch, A. Phase II study of paclitaxel plus the protein kinase C inhibitor bryostatin-1 in advanced pancreatic carcinoma. Am. J. Clin. Oncol. 2010, 33, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.D.; Jones, R.J.; Cho, E.; Kowalski, J.; Karp, J.E.; Gore, S.D.; Vala, M.; Meade, B.; Baker, S.D.; Zhao, M.; et al. Differentiation therapy in poor risk myeloid malignancies: Results of a dose finding study of the combination bryostatin-1 and GM-CSF. Leuk. Res. 2011, 35, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.J., Jr.; Leong, L.; Chow, W.; Gandara, D.; Frankel, P.; Garcia, A.; Lenz, H.J.; Doroshow, J.H. Phase II trial of bryostatin-1 in combination with cisplatin in patients with recurrent or persistent epithelial ovarian cancer: A California cancer consortium study. Investig. New Drugs 2012, 30, 723–728. [Google Scholar] [CrossRef] [Green Version]
- Ajani, J.A.; Jiang, Y.; Faust, J.; Chang, B.B.; Ho, L.; Yao, J.C.; Rousey, S.; Dakhil, S.; Cherny, R.C.; Craig, C.; et al. A multi-center phase II study of sequential paclitaxel and bryostatin-1 (NSC 339555) in patients with untreated, advanced gastric or gastroesophageal junction adenocarcinoma. Investig. New Drugs 2006, 24, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Laird, G.M.; Bullen, C.K.; Rosenbloom, D.I.; Martin, A.R.; Hill, A.L.; Durand, C.M.; Siliciano, J.D.; Siliciano, R.F. Ex vivo analysis identifies effective HIV-1 latency-reversing drug combinations. J. Clin. Investig. 2015, 125, 1901–1912. [Google Scholar] [CrossRef] [PubMed]
- Cillo, A.R.; Sobolewski, M.D.; Bosch, R.J.; Fyne, E.; Piatak, M., Jr.; Coffin, J.M.; Mellors, J.W. Quantification of HIV-1 latency reversal in resting CD4+ T cells from patients on suppressive antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2014, 111, 7078–7083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker-Sperling, V.E.; Pohlmeyer, C.W.; Tarwater, P.M.; Blankson, J.N. The Effect of Latency Reversal Agents on Primary CD8+ T Cells: Implications for Shock and Kill Strategies for Human Immunodeficiency Virus Eradication. EBioMedicine 2016, 8, 217–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pace, M.; Williams, J.; Kurioka, A.; Gerry, A.B.; Jakobsen, B.; Klenerman, P.; Nwokolo, N.; Fox, J.; Fidler, S.; Frater, J.; et al. Histone Deacetylase Inhibitors Enhance CD4 T Cell Susceptibility to NK Cell Killing but Reduce NK Cell Function. PLoS Pathog. 2016, 12, e1005782. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.B.; O’Connor, R.; Mueller, S.; Foley, M.; Szeto, G.L.; Karel, D.; Lichterfeld, M.; Kovacs, C.; Ostrowski, M.A.; Trocha, A.; et al. Histone deacetylase inhibitors impair the elimination of HIV-infected cells by cytotoxic T-lymphocytes. PLoS Pathog. 2014, 10, e1004287. [Google Scholar] [CrossRef] [Green Version]
- Tsai, A.; Irrinki, A.; Kaur, J.; Cihlar, T.; Kukolj, G.; Sloan, D.D.; Murry, J.P. Toll-Like Receptor 7 Agonist GS-9620 Induces HIV Expression and HIV-Specific Immunity in Cells from HIV-Infected Individuals on Suppressive Antiretroviral Therapy. J. Virol. 2017, 91, e02166-16. [Google Scholar] [CrossRef] [Green Version]
- Borducchi, E.N.; Cabral, C.; Stephenson, K.E.; Liu, J.; Abbink, P.; Ng’ang’a, D.; Nkolola, J.P.; Brinkman, A.L.; Peter, L.; Lee, B.C.; et al. Ad26/MVA therapeutic vaccination with TLR7 stimulation in SIV-infected rhesus monkeys. Nature 2016, 540, 284. [Google Scholar] [CrossRef]
- Borducchi, E.N.; Liu, J.; Nkolola, J.P.; Cadena, A.M.; Yu, W.H.; Fischinger, S.; Broge, T.; Abbink, P.; Mercado, N.B.; Chandrashekar, A.; et al. Publisher Correction: Antibody and TLR7 agonist delay viral rebound in SHIV-infected monkeys. Nature 2018, 564, E8. [Google Scholar] [CrossRef]
- Vibholm, L.; Schleimann, M.H.; Hojen, J.F.; Benfield, T.; Offersen, R.; Rasmussen, K.; Olesen, R.; Dige, A.; Agnholt, J.; Grau, J.; et al. Short-Course Toll-Like Receptor 9 Agonist Treatment Impacts Innate Immunity and Plasma Viremia in Individuals With Human Immunodeficiency Virus Infection. Clin. Infect. Dis. 2017, 64, 1686–1695. [Google Scholar] [CrossRef]
- Jones, R.B.; Mueller, S.; O’Connor, R.; Rimpel, K.; Sloan, D.D.; Karel, D.; Wong, H.C.; Jeng, E.K.; Thomas, A.S.; Whitney, J.B.; et al. A Subset of Latency-Reversing Agents Expose HIV-Infected Resting CD4+ T-Cells to Recognition by Cytotoxic T-Lymphocytes. PLoS Pathog. 2016, 12, e1005545. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Catalfamo, M.; Li, Y.; Henkart, P.A.; Weng, N.P. IL-15 mimics T cell receptor crosslinking in the induction of cellular proliferation, gene expression, and cytotoxicity in CD8+ memory T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 6192–6197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, M.; Schachterle, W.; Oberle, K.; Aichele, P.; Diefenbach, A. Dendritic cells prime natural killer cells by trans-presenting interleukin 15. Immunity 2007, 26, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortier, E.; Woo, T.; Advincula, R.; Gozalo, S.; Ma, A. IL-15Ralpha chaperones IL-15 to stable dendritic cell membrane complexes that activate NK cells via trans presentation. J. Exp. Med. 2008, 205, 1213–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldmann, T.A. The biology of interleukin-2 and interleukin-15: Implications for cancer therapy and vaccine design. Nat. Rev. Immunol. 2006, 6, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Wu, X.; Lu, Y.; Guo, W.; Andrieu, J.M. Therapeutic dendritic-cell vaccine for simian AIDS. Nat. Med. 2003, 9, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Arraes, L.C.; Ferreira, W.T.; Andrieu, J.M. Therapeutic dendritic-cell vaccine for chronic HIV-1 infection. Nat. Med. 2004, 10, 1359–1365. [Google Scholar] [CrossRef]
- Connolly, N.C.; Whiteside, T.L.; Wilson, C.; Kondragunta, V.; Rinaldo, C.R.; Riddler, S.A. Therapeutic immunization with human immunodeficiency virus type 1 (HIV-1) peptide-loaded dendritic cells is safe and induces immunogenicity in HIV-1-infected individuals. Clin. Vaccine Immunol. 2008, 15, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.; Climent, N.; Guardo, A.C.; Gil, C.; Leon, A.; Autran, B.; Lifson, J.D.; Martinez-Picado, J.; Dalmau, J.; Clotet, B.; et al. A dendritic cell-based vaccine elicits T cell responses associated with control of HIV-1 replication. Sci. Transl. Med. 2013, 5, 166ra2. [Google Scholar] [CrossRef]
- Jacobson, J.M.; Routy, J.P.; Welles, S.; DeBenedette, M.; Tcherepanova, I.; Angel, J.B.; Asmuth, D.M.; Stein, D.K.; Baril, J.G.; McKellar, M.; et al. Dendritic Cell Immunotherapy for HIV-1 Infection Using Autologous HIV-1 RNA: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Acquir. Immune Defic. Syndr. 2016, 72, 31. [Google Scholar] [CrossRef] [Green Version]
- Gay, C.L.; DeBenedette, M.A.; Tcherepanova, I.Y.; Gamble, A.; Lewis, W.E.; Cope, A.B.; Kuruc, J.D.; McGee, K.S.; Kearney, M.F.; Coffin, J.M.; et al. Immunogenicity of AGS-004 Dendritic Cell Therapy in Patients Treated During Acute HIV Infection. AIDS Res. Hum. Retrovir. 2018, 34, 111–122. [Google Scholar] [CrossRef]
- Routy, J.P.; Nicolette, C. Arcelis AGS-004 dendritic cell-based immunotherapy for HIV infection. Immunotherapy 2010, 2, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Routy, J.P.; Boulassel, M.R.; Yassine-Diab, B.; Nicolette, C.; Healey, D.; Jain, R.; Landry, C.; Yegorov, O.; Tcherepanova, I.; Monesmith, T.; et al. Immunologic activity and safety of autologous HIV RNA-electroporated dendritic cells in HIV-1 infected patients receiving antiretroviral therapy. Clin. Immunol. 2010, 134, 140–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guardo, A.C.; Joe, P.T.; Miralles, L.; Bargalló, M.E.; Mothe, B.; Krasniqi, A.; Heirman, C.; García, F.; Thielemans, K.; Brander, C.; et al. Preclinical evaluation of an mRNA HIV vaccine combining rationally selected antigenic sequences and adjuvant signals (HTI-TriMix). Aids 2017, 31, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Bonehill, A.; Tuyaerts, S.; Van Nuffel, A.M.; Heirman, C.; Bos, T.J.; Fostier, K.; Neyns, B.; Thielemans, K. Enhancing the T-cell stimulatory capacity of human dendritic cells by co-electroporation with CD40L, CD70 and constitutively active TLR4 encoding mRNA. Mol. Ther. 2008, 16, 1170–1180. [Google Scholar] [CrossRef]
- Mothe, B.; Hu, X.; Llano, A.; Rosati, M.; Olvera, A.; Kulkarni, V.; Valentin, A.; Alicea, C.; Pilkington, G.R.; Sardesai, N.Y.; et al. A human immune data-informed vaccine concept elicits strong and broad T-cell specificities associated with HIV-1 control in mice and macaques. J. Transl. Med. 2015, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Rinaldo, C.R. A novel anti-HIV immunotherapy to cure HIV. Aids 2017, 31, 447–449. [Google Scholar] [CrossRef]
- Surenaud, M.; Montes, M.; Arlehamn, C.S.L.; Sette, A.; Banchereau, J.; Palucka, K.; Lelievre, J.D.; Lacabaratz, C.; Levy, Y. Anti-HIV potency of T-cell responses elicited by dendritic cell therapeutic vaccination. PLoS Pathog. 2019, 15, e1008011. [Google Scholar] [CrossRef]
- Coelho, A.V.; de Moura, R.R.; Kamada, A.J.; da Silva, R.C.; Guimaraes, R.L.; Brandao, L.A.; de Alencar, L.C.; Crovella, S. Dendritic Cell-Based Immunotherapies to Fight HIV: How Far from a Success Story? A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2016, 17, 1985. [Google Scholar] [CrossRef] [Green Version]
- Garcia, F.; Routy, J.P. Challenges in dendritic cells-based therapeutic vaccination in HIV-1 infection Workshop in dendritic cell-based vaccine clinical trials in HIV-1. Vaccine 2011, 29, 6454–6463. [Google Scholar] [CrossRef]
- Brezar, V.; Ruffin, N.; Richert, L.; Surenaud, M.; Lacabaratz, C.; Palucka, K.; Thiebaut, R.; Banchereau, J.; Levy, Y.; Seddiki, N. Decreased HIV-specific T-regulatory responses are associated with effective DC-vaccine induced immunity. PLoS Pathog. 2015, 11, e1004752. [Google Scholar] [CrossRef]
- Levy, Y.; Thiebaut, R.; Montes, M.; Lacabaratz, C.; Sloan, L.; King, B.; Perusat, S.; Harrod, C.; Cobb, A.; Roberts, L.K.; et al. Dendritic cell-based therapeutic vaccine elicits polyfunctional HIV-specific T-cell immunity associated with control of viral load. Eur. J. Immunol. 2014, 44, 2802–2810. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Useros, N.; Lorizate, M.; McLaren, P.J.; Telenti, A.; Krausslich, H.G.; Martinez-Picado, J. HIV-1 capture and transmission by dendritic cells: The role of viral glycolipids and the cellular receptor Siglec-1. PLoS Pathog. 2014, 10, e1004146. [Google Scholar] [CrossRef] [PubMed]
- Rappocciolo, G.; Jais, M.; Piazza, P.; Reinhart, T.A.; Berendam, S.J.; Garcia-Exposito, L.; Gupta, P.; Rinaldo, C.R. Alterations in cholesterol metabolism restrict HIV-1 trans infection in nonprogressors. MBio 2014, 5, e01031-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.A.; van der Sluis, R.M.; Mota, T.; Pascoe, R.; Evans, V.A.; Lewin, S.R.; Cameron, P.U. Myeloid Dendritic Cells Induce HIV Latency in Proliferating CD4+ T Cells. J. Immunol. 2018, 201, 1468–1477. [Google Scholar] [CrossRef] [Green Version]
- Evans, V.A.; Kumar, N.; Filali, A.; Procopio, F.A.; Yegorov, O.; Goulet, J.P.; Saleh, S.; Haddad, E.K.; da Fonseca Pereira, C.; Ellenberg, P.C.; et al. Myeloid dendritic cells induce HIV-1 latency in non-proliferating CD4+ T cells. PLoS Pathog. 2013, 9, e1003799. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.Q.; Huang, X.L.; Gupta, P.; Borowski, L.; Fan, Z.; Watkins, S.C.; Thomas, E.K.; Rinaldo, C.R., Jr. Induction of anti-human immunodeficiency virus type 1 (HIV-1) CD8+ and CD4+ T-cell reactivity by dendritic cells loaded with HIV-1 X4-infected apoptotic cells. J. Virol. 2002, 76, 3007–3014. [Google Scholar] [CrossRef] [Green Version]
- Macatangay, B.J.; Riddler, S.A.; Wheeler, N.D.; Spindler, J.; Lawani, M.; Hong, F.; Buffo, M.J.; Whiteside, T.L.; Kearney, M.F.; Mellors, J.W.; et al. Therapeutic Vaccination With Dendritic Cells Loaded With Autologous HIV Type 1-Infected Apoptotic Cells. J. Infect. Dis. 2016, 213, 1400–1409. [Google Scholar] [CrossRef] [Green Version]
- Van der Sluis, R.M.; van Montfort, T.; Pollakis, G.; Sanders, R.W.; Speijer, D.; Berkhout, B.; Jeeninga, R.E. Dendritic cell-induced activation of latent HIV-1 provirus in actively proliferating primary T lymphocytes. PLoS Pathog. 2013, 9, e1003259. [Google Scholar] [CrossRef] [Green Version]
- Marini, A.; Harper, J.M.; Romerio, F. An In Vitro System to Model the Establishment and Reactivation of HIV-1 Latency. J. Immunol. 2008, 181, 7713–7720. [Google Scholar] [CrossRef]
- Ren, X.X.; Ma, L.; Sun, W.W.; Kuang, W.D.; Li, T.S.; Jin, X.; Wang, J.H. Dendritic cells maturated by co-culturing with HIV-1 latently infected Jurkat T cells or stimulating with AIDS-associated pathogens secrete TNF-alpha to reactivate HIV-1 from latency. Virulence 2017, 8, 1732–1743. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, A.; Mailliard, R.B.; Rinaldo, C.R.; Ratner, D.; Ding, M.; Chen, Y.; Zerbato, J.M.; Giacobbi, N.S.; Venkatachari, N.J.; Patterson, B.K.; et al. Novel assay reveals a large, inducible, replication-competent HIV-1 reservoir in resting CD4+ T cells. Nat. Med. 2017, 23, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Spina, C.A.; Anderson, J.; Archin, N.M.; Bosque, A.; Chan, J.; Famiglietti, M.; Greene, W.C.; Kashuba, A.; Lewin, S.R.; Margolis, D.M.; et al. An in-depth comparison of latent HIV-1 reactivation in multiple cell model systems and resting CD4+ T cells from aviremic patients. PLoS Pathog. 2013, 9, e1003834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbato, J.M.; Serrao, E.; Lenzi, G.; Kim, B.; Ambrose, Z.; Watkins, S.C.; Engelman, A.N.; Sluis-Cremer, N. Establishment and Reversal of HIV-1 Latency in Naive and Central Memory CD4+ T Cells In Vitro. J. Virol. 2016, 90, 8059–8073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsden, M.D.; Loy, B.A.; Wu, X.; Ramirez, C.M.; Schrier, A.J.; Murray, D.; Shimizu, A.; Ryckbosch, S.M.; Near, K.E.; Chun, T.W.; et al. In vivo activation of latent HIV with a synthetic bryostatin analog effects both latent cell “kick” and “kill” in strategy for virus eradication. PLoS Pathog. 2017, 13, e1006575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, T.D.; Miller, E.A.; Bhardwaj, N.; Landau, N.R. Vpx-containing dendritic cell vaccine induces CTLs and reactivates latent HIV-1 in vitro. Gene Ther. 2015, 22, 227. [Google Scholar] [CrossRef] [Green Version]
- Van Montfort, T.; van der Sluis, R.; Darcis, G.; Beaty, D.; Groen, K.; Pasternak, A.O.; Pollakis, G.; Vink, M.; Westerhout, E.M.; Hamdi, M.; et al. Dendritic cells potently purge latent HIV-1 beyond TCR-stimulation, activating the PI3K-Akt-mTOR pathway. Ebiomedicine 2019, 42, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Van der Sluis, R.M.; van Capel, T.M.; Speijer, D.; Sanders, R.W.; Berkhout, B.; de Jong, E.C.; Jeeninga, R.E.; van Montfort, T. Dendritic cell type-specific HIV-1 activation in effector T cells: Implications for latent HIV-1 reservoir establishment. Aids 2015, 29, 1003–1014. [Google Scholar] [CrossRef]
- Douek, D.C.; Brenchley, J.M.; Betts, M.R.; Ambrozak, D.R.; Hill, B.J.; Okamoto, Y.; Casazza, J.P.; Kuruppu, J.; Kunstman, K.; Wolinsky, S.; et al. HIV preferentially infects HIV-specific CD4+ T cells. Nature 2002, 417, 95–98. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Ruff, L.E.; Casazza, J.P.; Koup, R.A.; Price, D.A.; Douek, D.C. Preferential infection shortens the life span of human immunodeficiency virus-specific CD4+ T cells in vivo. J. Virol. 2006, 80, 6801–6809. [Google Scholar] [CrossRef] [Green Version]
- Demoustier, A.; Gubler, B.; Lambotte, O.; de Goer, M.G.; Wallon, C.; Goujard, C.; Delfraissy, J.F.; Taoufik, Y. In patients on prolonged HAART, a significant pool of HIV infected CD4 T cells are HIV-specific. Aids 2002, 16, 1749–1754. [Google Scholar] [CrossRef] [Green Version]
- Casazza, J.P.; Brenchley, J.M.; Hill, B.J.; Ayana, R.; Ambrozak, D.; Roederer, M.; Douek, D.C.; Betts, M.R.; Koup, R.A. Autocrine production of beta-chemokines protects CMV-Specific CD4 T cells from HIV infection. PLoS Pathog. 2009, 5, e1000646. [Google Scholar] [CrossRef] [PubMed]
- Stone, S.F.; Price, P.; French, M.A. Cytomegalovirus (CMV)-specific CD8+ T cells in individuals with HIV infection: Correlation with protection from CMV disease. J. Antimicrob. Chemother. 2006, 57, 585–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylwester, A.W.; Mitchell, B.L.; Edgar, J.B.; Taormina, C.; Pelte, C.; Ruchti, F.; Sleath, P.R.; Grabstein, K.H.; Hosken, N.A.; Kern, F.; et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 2005, 202, 673–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abana, C.O.; Pilkinton, M.A.; Gaudieri, S.; Chopra, A.; McDonnell, W.J.; Wanjalla, C.; Barnett, L.; Gangula, R.; Hager, C.; Jung, D.K.; et al. Cytomegalovirus (CMV) Epitope-Specific CD4+ T Cells Are Inflated in HIV+ CMV+ Subjects. J. Immunol. 2017, 199, 3187–3201. [Google Scholar] [CrossRef] [PubMed]
- Komanduri, K.V.; Donahoe, S.M.; Moretto, W.J.; Schmidt, D.K.; Gillespie, G.; Ogg, G.S.; Roederer, M.; Nixon, D.F.; McCune, J.M. Direct measurement of CD4+ and CD8+ T-cell responses to CMV in HIV-1-infected subjects. Virology 2001, 279, 459–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeger, D.M.; Martin, J.N.; Sinclair, E.; Hunt, P.W.; Bangsberg, D.R.; Hecht, F.; Hsue, P.; McCune, J.M.; Deeks, S.G. Cytomegalovirus-specific T cells persist at very high levels during long-term antiretroviral treatment of HIV disease. PLoS ONE 2010, 5, e8886. [Google Scholar] [CrossRef] [Green Version]
- Soderberg-Naucler, C. Treatment of cytomegalovirus infections beyond acute disease to improve human health. Expert Rev. Anti Infect. Ther. 2014, 12, 211–222. [Google Scholar] [CrossRef]
- Maidji, E.; Somsouk, M.; Rivera, J.M.; Hunt, P.W.; Stoddart, C.A. Replication of CMV in the gut of HIV-infected individuals and epithelial barrier dysfunction. PLoS Pathog. 2017, 13, e1006202. [Google Scholar] [CrossRef] [Green Version]
- Gianella, S.; Chaillon, A.; Mutlu, E.A.; Engen, P.A.; Voigt, R.M.; Keshavarzian, A.; Losurdo, J.; Chakradeo, P.; Lada, S.M.; Nakazawa, M.; et al. Effect of cytomegalovirus and Epstein-Barr virus replication on intestinal mucosal gene expression and microbiome composition of HIV-infected and uninfected individuals. Aids 2017, 31, 2059–2067. [Google Scholar] [CrossRef]
- Christensen-Quick, A.; Vanpouille, C.; Lisco, A.; Gianella, S. Cytomegalovirus and HIV Persistence: Pouring Gas on the Fire. Aids Res. Hum. Retrovir. 2017, 33, S-23. [Google Scholar] [CrossRef]
- Gianella, S.; Anderson, C.M.; Var, S.R.; Oliveira, M.F.; Lada, S.M.; Vargas, M.V.; Massanella, M.; Little, S.J.; Richman, D.D.; Strain, M.C.; et al. Replication of Human Herpesviruses Is Associated with Higher HIV DNA Levels during Antiretroviral Therapy Started at Early Phases of HIV Infection. J. Virol. 2016, 90, 3944–3952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, M.L.; Lederman, M.M.; Gianella, S. Partners in Crime: The Role of CMV in Immune Dysregulation and Clinical Outcome During HIV Infection. Curr. HIV/AIDS Rep. 2016, 13, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.M.; Nakazawa, M.; Freeman, M.L.; Anderson, C.M.; Oliveira, M.F.; Little, S.J.; Gianella, S. Asymptomatic CMV Replication During Early Human Immunodeficiency Virus (HIV) Infection Is Associated With Lower CD4/CD8 Ratio During HIV Treatment. Clin. Infect. Dis. 2016, 63, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Poizot-Martin, I.; Allavena, C.; Duvivier, C.; Cano, C.E.; Guillouet de Salvador, F.; Rey, D.; Dellamonica, P.; Cuzin, L.; Cheret, A.; Hoen, B.; et al. CMV+ Serostatus Associates Negatively with CD4:CD8 Ratio Normalization in Controlled HIV-Infected Patients on cART. PLoS ONE 2016, 11, e0165774. [Google Scholar] [CrossRef]
- Dan, J.M.; Massanella, M.; Smith, D.M.; Spina, C.A.; Schrier, R.; Daar, E.S.; Dube, M.P.; Morris, S.R.; Gianella, S. Brief Report: Effect of CMV and HIV Transcription on CD57 and PD-1 T-Cell Expression During Suppressive ART. J. Acquir. Immune Defic. Syndr. 2016, 72, 133. [Google Scholar] [CrossRef]
- Agudelo-Hernandez, A.; Chen, Y.; Bullotta, A.; Buchanan, W.G.; Klamar-Blain, C.R.; Borowski, L.; Riddler, S.A.; Rinaldo, C.R.; Macatangay, B.J.C. Subclinical herpesvirus shedding among HIV-1-infected men on antiretroviral therapy. Aids 2017, 31, 2085–2094. [Google Scholar] [CrossRef]
- Johnson, E.L.; Howard, C.L.; Thurman, J.; Pontiff, K.; Johnson, E.S.; Chakraborty, R. Cytomegalovirus upregulates expression of CCR5 in central memory cord blood mononuclear cells, which may facilitate in utero HIV type 1 transmission. J. Infect. Dis. 2015, 211, 187–196. [Google Scholar] [CrossRef] [Green Version]
- Barry, P.A.; Pratt-Lowe, E.; Peterlin, B.M.; Luciw, P.A. Cytomegalovirus activates transcription directed by the long terminal repeat of human immunodeficiency virus type 1. J. Virol. 1990, 64, 2932–2940. [Google Scholar]
- McCarthy, M.; Auger, D.; He, J.; Wood, C. Cytomegalovirus and human herpesvirus-6 trans-activate the HIV-1 long terminal repeat via multiple response regions in human fetal astrocytes. J. Neurovirol. 1998, 4, 495–511. [Google Scholar] [CrossRef]
- Murayama, T.; Ohara, Y.; Obuchi, M.; Khabar, K.S.; Higashi, H.; Mukaida, N.; Matsushima, K. Human cytomegalovirus induces interleukin-8 production by a human monocytic cell line, THP-1, through acting concurrently on AP-1- and NF-kappaB-binding sites of the interleukin-8 gene. J. Virol. 1997, 71, 5692–5695. [Google Scholar]
- Saleh, S.; Lu, H.K.; Evans, V.; Harisson, D.; Zhou, J.; Jaworowski, A.; Sallmann, G.; Cheong, K.Y.; Mota, T.M.; Tennakoon, S.; et al. HIV integration and the establishment of latency in CCL19-treated resting CD4+ T cells require activation of NF-kappaB. Retrovirology 2016, 13, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianella, S.; Anderson, C.M.; Vargas, M.V.; Richman, D.D.; Little, S.J.; Morris, S.R.; Smith, D.M. Cytomegalovirus DNA in semen and blood is associated with higher levels of proviral HIV DNA. J. Infect. Dis. 2013, 207, 898–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gianella, S.; Massanella, M.; Richman, D.D.; Little, S.J.; Spina, C.A.; Vargas, M.V.; Lada, S.M.; Daar, E.S.; Dube, M.P.; Haubrich, R.H.; et al. Cytomegalovirus replication in semen is associated with higher levels of proviral HIV DNA and CD4+ T cell activation during antiretroviral treatment. J. Virol. 2014, 88, 7818–7827. [Google Scholar] [CrossRef] [Green Version]
- Wagner, T.A.; McKernan, J.L.; Tobin, N.H.; Tapia, K.A.; Mullins, J.I.; Frenkel, L.M. An increasing proportion of monotypic HIV-1 DNA sequences during antiretroviral treatment suggests proliferation of HIV-infected cells. J. Virol. 2013, 87, 1770–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Stockenstrom, S.; Odevall, L.; Lee, E.; Sinclair, E.; Bacchetti, P.; Killian, M.; Epling, L.; Shao, W.; Hoh, R.; Ho, T.; et al. Longitudinal Genetic Characterization Reveals That Cell Proliferation Maintains a Persistent HIV Type 1 DNA Pool During Effective HIV Therapy. J. Infect. Dis. 2015, 212, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Henrich, T.J.; Hobbs, K.S.; Hanhauser, E.; Scully, E.; Hogan, L.E.; Robles, Y.P.; Leadabrand, K.S.; Marty, F.M.; Palmer, C.D.; Jost, S.; et al. Human Immunodeficiency Virus Type 1 Persistence Following Systemic Chemotherapy for Malignancy. J. Infect. Dis. 2017, 216, 254–262. [Google Scholar] [CrossRef] [Green Version]
- Buzon, M.J.; Sun, H.; Li, C.; Shaw, A.; Seiss, K.; Ouyang, Z.; Martin-Gayo, E.; Leng, J.; Henrich, T.J.; Li, J.Z.; et al. HIV-1 persistence in CD4+ T cells with stem cell-like properties. Nat. Med. 2014, 20, 139–142. [Google Scholar] [CrossRef]
- Schmueck-Henneresse, M.; Sharaf, R.; Vogt, K.; Weist, B.J.; Landwehr-Kenzel, S.; Fuehrer, H.; Jurisch, A.; Babel, N.; Rooney, C.M.; Reinke, P.; et al. Peripheral blood-derived virus-specific memory stem T cells mature to functional effector memory subsets with self-renewal potency. J. Immunol. 2015, 194, 5559–5567. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.D.; Grundy, J.E. Down-regulation of the class I HLA heterodimer and beta 2-microglobulin on the surface of cells infected with cytomegalovirus. J. Gen. Virol. 1992, 73, 2395–2403. [Google Scholar] [CrossRef]
- Ulbrecht, M.; Martinozzi, S.; Grzeschik, M.; Hengel, H.; Ellwart, J.W.; Pla, M.; Weiss, E.H. Cutting edge: The human cytomegalovirus UL40 gene product contains a ligand for HLA-E and prevents NK cell-mediated lysis. J. Immunol. 2000, 164, 5019–5022. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Chang, P.C.; Wang, Y.; Li, M. Identification of novel viral interleukin-10 isoforms of human cytomegalovirus AD169. Virus Res. 2008, 131, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSharry, B.P.; Avdic, S.; Slobedman, B. Human cytomegalovirus encoded homologs of cytokines, chemokines and their receptors: Roles in immunomodulation. Viruses 2012, 4, 2448–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, H.; Jain, V.; Hunt, P.W.; Lee, T.H.; Sinclair, E.; Do, T.D.; Hoh, R.; Martin, J.N.; McCune, J.M.; Hecht, F.; et al. Cell-based measures of viral persistence are associated with immune activation and programmed cell death protein 1 (PD-1)-expressing CD4+ T cells. J. Infect. Dis. 2013, 208, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.; Garcia, M.; Petrovas, C.; Yamamoto, T.; Koup, R.A.; Nabel, G.J. HIV-1 causes CD4 cell death through DNA-dependent protein kinase during viral integration. Nature 2013, 498, 376–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, M.R.; Bak, R.O.; Andersen, A.; Berg, R.K.; Jensen, S.B.; Tengchuan, J.; Laustsen, A.; Hansen, K.; Ostergaard, L.; Fitzgerald, K.A.; et al. IFI16 senses DNA forms of the lentiviral replication cycle and controls HIV-1 replication. Proc. Natl. Acad. Sci. USA 2013, 110, E4571–E4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristoff, J.; Palma, M.L.; Garcia-Bates, T.M.; Shen, C.; Sluis-Cremer, N.; Gupta, P.; Rinaldo, C.R.; Mailliard, R.B. Type 1-programmed dendritic cells drive antigen-specific latency reversal and immune elimination of persistent HIV-1. EBiomedicine 2019, 43, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, Y.; Oshita, C.; Kume, A.; Iizuka, A.; Miyata, H.; Komiyama, M.; Ashizawa, T.; Yagoto, M.; Abe, Y.; Mitsuya, K.; et al. alpha-type-1 polarized dendritic cell-based vaccination in recurrent high-grade glioma: A phase I clinical trial. BMC Cancer 2012, 12, 623. [Google Scholar] [CrossRef] [Green Version]
- Fracol, M.; Xu, S.; Mick, R.; Fitzpatrick, E.; Nisenbaum, H.; Roses, R.; Fisher, C.; Tchou, J.; Fox, K.; Zhang, P.; et al. Response to HER-2 pulsed DC1 vaccines is predicted by both HER-2 and estrogen receptor expression in DCIS. Ann. Surg. Oncol. 2013, 20, 3233–3239. [Google Scholar] [CrossRef]
- Radomski, M.; Zeh, H.J.; Edington, H.D.; Pingpank, J.F.; Butterfield, L.H.; Whiteside, T.L.; Wieckowski, E.; Bartlett, D.L.; Kalinski, P. Prolonged intralymphatic delivery of dendritic cells through implantable lymphatic ports in patients with advanced cancer. J. Immunother. Cancer 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Schuler, P.J.; Harasymczuk, M.; Visus, C.; Deleo, A.; Trivedi, S.; Lei, Y.; Argiris, A.; Gooding, W.; Butterfield, L.H.; Whiteside, T.L.; et al. Phase I dendritic cell p53 peptide vaccine for head and neck cancer. Clin. Cancer Res. 2014, 20, 2433–2444. [Google Scholar] [CrossRef] [Green Version]
- Feau, S.; Arens, R.; Togher, S.; Schoenberger, S.P. Autocrine IL-2 is required for secondary population expansion of CD8+ memory T cells. Nat. Immunol. 2011, 12, 908–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feau, S.; Garcia, Z.; Arens, R.; Yagita, H.; Borst, J.; Schoenberger, S.P. The CD4+ T-cell help signal is transmitted from APC to CD8+ T-cells via CD27–CD70 interactions. Nat. Commun. 2012, 3, 948. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.D.; Kityo, C.; Ssali, F.; Swainson, L.; Makamdop, K.N.; Del Prete, G.Q.; Deeks, S.G.; Luciw, P.A.; Chipman, J.G.; Beilman, G.J.; et al. Defining total-body AIDS-virus burden with implications for curative strategies. Nat. Med. 2017, 23, 1271–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santangelo, P.J.; Rogers, K.A.; Zurla, C.; Blanchard, E.L.; Gumber, S.; Strait, K.; Connor-Stroud, F.; Schuster, D.M.; Amancha, P.K.; Hong, J.J.; et al. Whole-body immunoPET reveals active SIV dynamics in viremic and antiretroviral therapy-treated macaques. Nat. Methods 2015, 12, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Stieh, D.J.; Maric, D.; Kelley, Z.L.; Anderson, M.R.; Hattaway, H.Z.; Beilfuss, B.A.; Rothwangl, K.B.; Veazey, R.S.; Hope, T.J. Vaginal challenge with an SIV-based dual reporter system reveals that infection can occur throughout the upper and lower female reproductive tract. PLoS Pathog. 2014, 10, e1004440. [Google Scholar] [CrossRef]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef]
Figure 1.
Monocyte-derived DC (MDC1): the all-in-one ‘kick and kill’ tool. MDC1s induce antigen-specific CD8+ and CD4+ T cell responses through presentation of antigenic peptides in the context of (1) MHC class-I and (2) MHC class-II molecules, respectively (signal 1), along with costimulatory factors including CD80 and CD86 (not shown, signal 2). Responding CD4+ T cells subsequently provide MDC1 with the feedback hyperactivating ‘helper’ signal CD40L (3), necessary for MDC1 release of IL-12p70 (4), which then promotes expansion and differentiation of CD8+ human immunodeficiency virus-1 (HIV-1)-specific effector cytotoxic T lymphocytes (CTLs) (5). Activation of CMV and HIV antigen-responsive CD4+ T cells harboring latent HIV-1 (6) results in HIV latency reversal (7), with HIV-1 proteins being transcribed and expressed as surface antigen (8). As a result, exposed infected cells harboring replication-competent provirus are targeted for elimination by HIV-specific CTLs (9).
Figure 1.
Monocyte-derived DC (MDC1): the all-in-one ‘kick and kill’ tool. MDC1s induce antigen-specific CD8+ and CD4+ T cell responses through presentation of antigenic peptides in the context of (1) MHC class-I and (2) MHC class-II molecules, respectively (signal 1), along with costimulatory factors including CD80 and CD86 (not shown, signal 2). Responding CD4+ T cells subsequently provide MDC1 with the feedback hyperactivating ‘helper’ signal CD40L (3), necessary for MDC1 release of IL-12p70 (4), which then promotes expansion and differentiation of CD8+ human immunodeficiency virus-1 (HIV-1)-specific effector cytotoxic T lymphocytes (CTLs) (5). Activation of CMV and HIV antigen-responsive CD4+ T cells harboring latent HIV-1 (6) results in HIV latency reversal (7), with HIV-1 proteins being transcribed and expressed as surface antigen (8). As a result, exposed infected cells harboring replication-competent provirus are targeted for elimination by HIV-specific CTLs (9).


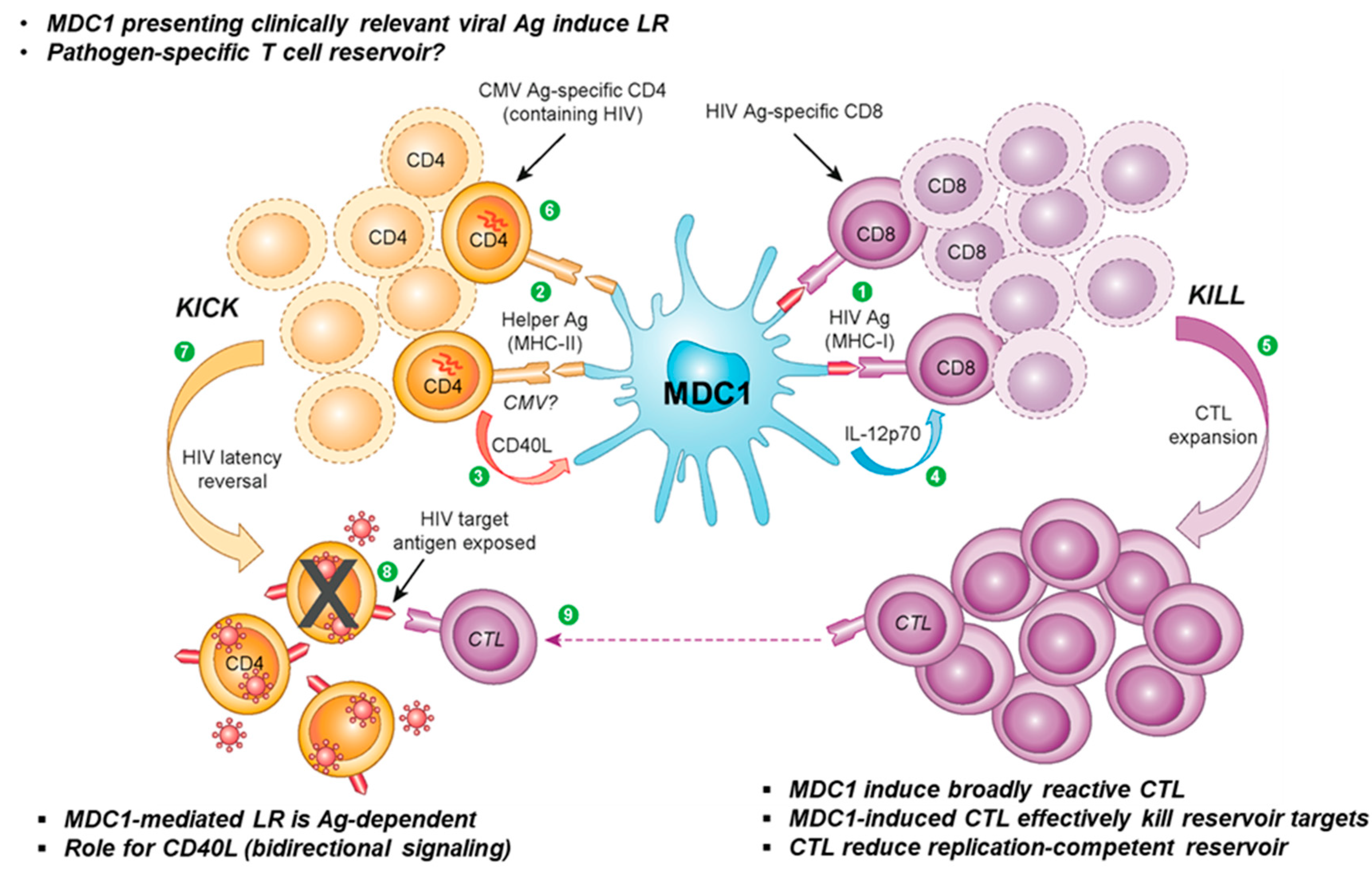{kind=link}
Table 1.
Immunotherapeutic potential of ex vivo-educated dendritic cells (DCs).
| Desired Trait | PGE2 DC | Type 1-polarized DC | References |
|---|---|---|---|
| Antigen Presentation | ++++ | ++++++ | [52] |
| Maturation status (high CD83 / CD86) | ++++++ | ++++++ | [62,64] |
| Lymph node homing (CCR7) | ++++++ | ++++ | [60,61,62,63,64] |
| IL-12p70, IL-15 production | Deficient | ++++++++ | [62,64,72] |
| T cell trafficking / chemokine expression | CCL22 (TREG) | CCL19 (TN, TCM) | [63,69,73,74] |
| CCL3-5, CXCL9-11 (TEM) | |||
| Antigen transfer ability | Deficient | ++++++ | [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kristoff, J.; Rinaldo, C.R.; Mailliard, R.B. Role of Dendritic Cells in Exposing Latent HIV-1 for the Kill. Viruses 2020, 12, 37. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010037
AMA Style
Kristoff J, Rinaldo CR, Mailliard RB. Role of Dendritic Cells in Exposing Latent HIV-1 for the Kill. Viruses. 2020; 12(1):37. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010037
Chicago/Turabian StyleKristoff, Jan, Charles R. Rinaldo, and Robbie B. Mailliard. 2020. "Role of Dendritic Cells in Exposing Latent HIV-1 for the Kill" Viruses 12, no. 1: 37. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010037
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.