Investigation of the Role of the Spike Protein in Reversing the Virulence of the Highly Virulent Taiwan Porcine Epidemic Diarrhea Virus Pintung 52 Strains and Its Attenuated Counterpart


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Cells and Viruses
2.3. Generation and Recovery of Recombinant iPEDVPT-P5, iPEDVPT-P96, iPEDVPT-P5-96S and iPEDVPT-P96-5S Viruses
2.4. In Vitro Characterization of Recombinant iPEDVPT-P5, iPEDVPT-P96, iPEDVPT-P5-96S and iPEDVPT-P96-5S Viruses
2.4.1. Immunofluorescence Assay and Syncytia Analysis
2.4.2. Sequence Analysis
2.4.3. Growth Kinetics, Viral Titration and Plaque Assay
2.5. Animal Experiment
2.5.1. Quantification of PEDV Fecal Viral Shedding
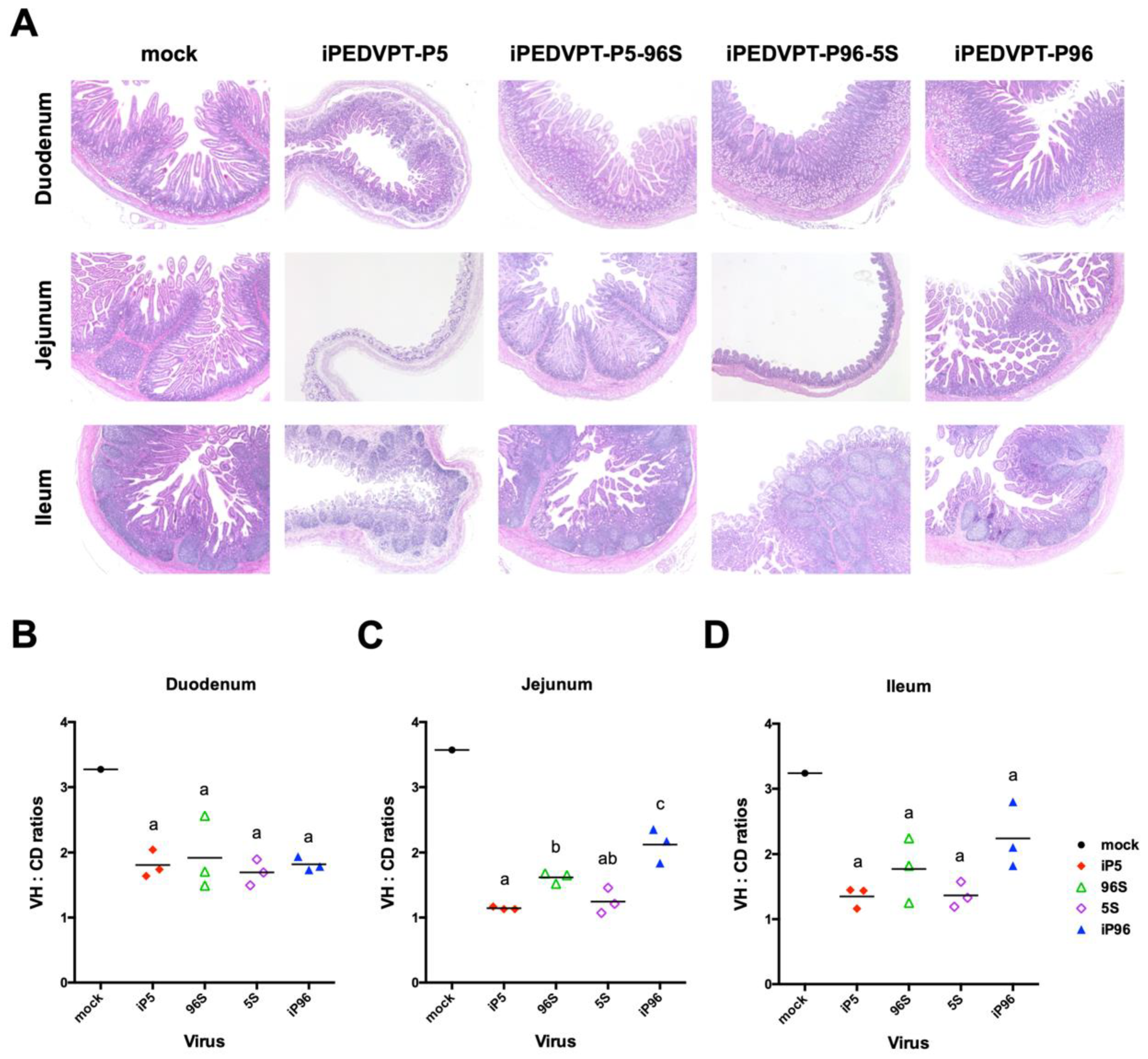
2.5.2. Histopathology and Immunohistochemistry
2.6. Statistical Analysis
3. Results
3.1. Recovery of recombinant PEDVs
3.2. In Vitro Characterization of Recombinant PEDVs
3.3. Investigation of the Role of Spike Gene on the Pathogenicity of PEDVPT 52 strain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pensaert, M.B.; de Bouck, P. A new coronavirus-like particle associated with diarrhea in swine. Arch. Virol. 1978, 58, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Li, H.; Liu, Y.; Pan, Y.; Deng, F.; Song, Y.; Tang, X.; He, Q. New variants of porcine epidemic diarrhea virus, China, 2011. Emerg. Infect. Dis. 2012, 18, 1350–1353. [Google Scholar] [CrossRef] [PubMed]
- Chiou, H.Y.; Huang, Y.L.; Deng, M.C.; Chang, C.Y.; Jeng, C.R.; Tsai, P.S.; Yang, C.; Pang, V.F.; Chang, H.W. Phylogenetic Analysis of the Spike (S) Gene of the New Variants of Porcine Epidemic Diarrhoea Virus in Taiwan. Transbound. Emerg. Dis. 2017, 64, 157–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, L.L.; Tonsor, G.T. Assessment of the economic impacts of porcine epidemic diarrhea virus in the United States. J. Anim. Sci. 2015, 93, 5111–5118. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, F.C.; Lin, C.N.; Chiou, H.Y.; Chia, M.Y.; Chiou, M.T.; Haga, T.; Kao, C.F.; Chang, Y.C.; Chang, C.Y.; Jeng, C.R.; et al. Updated phylogenetic analysis of the spike gene and identification of a novel recombinant porcine epidemic diarrhoea virus strain in Taiwan. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef]
- Duarte, M.; Gelfi, J.; Lambert, P.; Rasschaert, D.; Laude, H. Genome organization of porcine epidemic diarrhoea virus. Adv. Exp. Med. Biol. 1993, 342, 55–60. [Google Scholar] [CrossRef]
- Li, F. Structure, Function, and Evolution of Coronavirus Spike Proteins. Annu. Rev. Virol. 2016, 3, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, I.N.; de Vries, R.P.; Grone, A.; de Haan, C.A.; Verheije, M.H. Binding of avian coronavirus spike proteins to host factors reflects virus tropism and pathogenicity. J. Virol. 2011, 85, 8903–8912. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.W.; Egberink, H.F.; Halpin, R.; Spiro, D.J.; Rottier, P.J. Spike protein fusion peptide and feline coronavirus virulence. Emerg. Infect. Dis. 2012, 18, 1089–1095. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J. The coronavirus spike protein is a class I virus fusion protein: Structural and functional characterization of the fusion core complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wang, Q. Emerging Highly Virulent Porcine Epidemic Diarrhea Virus: Molecular Mechanisms of Attenuation and Rational Design of Live Attenuated Vaccines. Int. J. Mol. Sci. 2019, 20, 5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, K.; Lager, K.M.; Kulshreshtha, V.; Miller, L.C.; Faaberg, K.S. Status of vaccines for porcine epidemic diarrhea virus in the United States and Canada. Virus Res. 2016, 226, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdts, V.; Zakhartchouk, A. Vaccines for porcine epidemic diarrhea virus and other swine coronaviruses. Vet. Microbiol. 2017, 206, 45–51. [Google Scholar] [CrossRef]
- Stobart, C.C.; Moore, M.L. RNA virus reverse genetics and vaccine design. Viruses 2014, 6, 2531–2550. [Google Scholar] [CrossRef]
- Hou, Y.; Ke, H.; Kim, J.; Yoo, D.; Su, Y.; Boley, P.; Chepngeno, J.; Vlasova, A.N.; Saif, L.J.; Wang, Q. Engineering a Live Attenuated Porcine Epidemic Diarrhea Virus Vaccine Candidate via Inactivation of the Viral 2’-O-Methyltransferase and the Endocytosis Signal of the Spike Protein. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Ge, X.; Chen, D.; Li, J.; Cai, Y.; Deng, J.; Zhou, L.; Guo, X.; Han, J.; Yang, H. The S Gene Is Necessary but Not Sufficient for the Virulence of Porcine Epidemic Diarrhea Virus Novel Variant Strain BJ2011C. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Kao, C.F.; Chang, C.Y.; Jeng, C.R.; Tsai, P.S.; Pang, V.F.; Chiou, H.Y.; Peng, J.Y.; Cheng, I.C.; Chang, H.W. Evaluation and Comparison of the Pathogenicity and Host Immune Responses Induced by a G2b Taiwan Porcine Epidemic Diarrhea Virus (Strain Pintung 52) and Its Highly Cell-Culture Passaged Strain in Conventional 5-Week-Old Pigs. Viruses 2017, 9, 121. [Google Scholar] [CrossRef]
- Kao, C.F.; Chiou, H.Y.; Chang, Y.C.; Hsueh, C.S.; Jeng, C.R.; Tsai, P.S.; Cheng, I.C.; Pang, V.F.; Chang, H.W. The Characterization of Immunoprotection Induced by a cDNA Clone Derived from the Attenuated Taiwan Porcine Epidemic Diarrhea Virus Pintung 52 Strain. Viruses 2018, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Cheng, I.C.; Chang, Y.C.; Tsai, P.S.; Lai, S.Y.; Huang, Y.L.; Jeng, C.R.; Pang, V.F.; Chang, H.W. Identification of Neutralizing Monoclonal Antibodies Targeting Novel Conformational Epitopes of the Porcine Epidemic Diarrhoea Virus Spike Protein. Sci. Rep. 2019, 9, 2529. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty per cent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Peng, J.Y.; Chang, C.Y.; Kao, C.F.; Chang, Y.C.; Hsueh, C.S.; Jeng, C.R.; Cheng, I.C.; Pang, V.F.; Tsai, P.S.; Chang, H.W. Different intestinal tropism of the G2b Taiwan porcine epidemic diarrhea virus-Pintung 52 strain in conventional 7-day-old piglets. Vet. J. 2018, 237, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Lin, C.M.; Yokoyama, M.; Yount, B.L.; Marthaler, D.; Douglas, A.L.; Ghimire, S.; Qin, Y.; Baric, R.S.; Saif, L.J.; et al. Deletion of a 197-Amino-Acid Region in the N-Terminal Domain of Spike Protein Attenuates Porcine Epidemic Diarrhea Virus in Piglets. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Zhong, H.; Zhou, Q.; Du, Y.; Chen, L.; Zhang, Y.; Xue, C.; Cao, Y. A Highly Pathogenic Strain of Porcine Deltacoronavirus Caused Watery Diarrhea in Newborn Piglets. Virol. Sin. 2018, 33, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255–258. [Google Scholar] [CrossRef]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef]
- Deng, X.; van Geelen, A.; Buckley, A.C.; O’Brien, A.; Pillatzki, A.; Lager, K.M.; Faaberg, K.S.; Baker, S.C. Coronavirus Endoribonuclease Activity in Porcine Epidemic Diarrhea Virus Suppresses Type I and Type III Interferon Responses. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, J.; Jin, Z.; Gao, Y.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. Development of the full-length cDNA clones of two porcine epidemic diarrhea disease virus isolates with different virulence. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Lin, C.M.; Hou, Y.; Marthaler, D.G.; Gao, X.; Liu, X.; Zheng, L.; Saif, L.J.; Wang, Q. Attenuation of an original US porcine epidemic diarrhea virus strain PC22A via serial cell culture passage. Vet. Microbiol. 2017, 201, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Terada, Y.; Enjuanes, L.; Ohashi, S.; Kamitani, W. S1 Subunit of Spike Protein from a Current Highly Virulent Porcine Epidemic Diarrhea Virus Is an Important Determinant of Virulence in Piglets. Viruses 2018, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Li, W.; Lucio de Esesarte, E.; Guo, H.; van den Elzen, P.; Aarts, E.; van den Born, E.; Rottier, P.J.M.; Bosch, B.J. Cell Attachment Domains of the Porcine Epidemic Diarrhea Virus Spike Protein Are Key Targets of Neutralizing Antibodies. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position in Spike Gene | Nucleotide | Amino Acid | ||||
|---|---|---|---|---|---|---|
| Nucleotide | Amino Acid | Domain | iPEDVPT-P5 | iPEDVPT-P96 | iPEDVPT-P5 | iPEDVPT-P96 |
| 431 | 144 | S10 | C | T | T | I |
| 1661 | 554 | S1B | T | C | F | S |
| 2659 | 887 | S2 | A | C | S | R |
| 2902 | 968 | S2 | T | G | S | A |
| 3062 | 1021 | S2 HR1 † | T | G | I | S |
| 3077 | 1026 | S2 HR1 † | A | G | K | R |
| 3755 | 1252 | S2 | T | G | L | R |
| 4061 | 1354 | S2 CT ‡ | G | T | C | F |
| 4073 | 1358 | S2 CT ‡ | G | T | C | F |
| 4134 | 1378 | S2 CT ‡ | A | T | E | D |
| Group | Mortality Rate (%) | Peak Viral Shedding (DPI) | Peak Viral Shedding (log10 copies/mL) | Onset of Diarrhea (DPI) † | Mean Average Daily Weight Gain ‡ |
|---|---|---|---|---|---|
| iPEDVPT-P5 | 87.5 (7/8) | 1 | 10.06 ± 1.14 | 1.00 ± 0.0 | −0.116 ± 0.135 |
| iPEDVPT-P5-96S | 80.0 (4/5) | 7 | 9.15 ± 0/91 | 3.83 ± 1.47 | −0.033 ± 0.062 |
| iPEDVPT-P96-5S | 83.4 (5/6) | 1 | 8.20 ± 1.70 | 1.00 ± 0.0 | −0.035 ± 0.049 |
| iPEDVPT-P96 | 40.0 (2/5) | 10 | 9.74 ± 0.74 | 2.38 ± 1.51 | −0.016 ± 0.055 |
| Mock | 0.0 (4/4) | ND | ND | ND | 0.051 ± 0.016 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kao, C.-F.; Chang, H.-W. Investigation of the Role of the Spike Protein in Reversing the Virulence of the Highly Virulent Taiwan Porcine Epidemic Diarrhea Virus Pintung 52 Strains and Its Attenuated Counterpart. Viruses 2020, 12, 41. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010041
Kao C-F, Chang H-W. Investigation of the Role of the Spike Protein in Reversing the Virulence of the Highly Virulent Taiwan Porcine Epidemic Diarrhea Virus Pintung 52 Strains and Its Attenuated Counterpart. Viruses. 2020; 12(1):41. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010041
Chicago/Turabian StyleKao, Chi-Fei, and Hui-Wen Chang. 2020. "Investigation of the Role of the Spike Protein in Reversing the Virulence of the Highly Virulent Taiwan Porcine Epidemic Diarrhea Virus Pintung 52 Strains and Its Attenuated Counterpart" Viruses 12, no. 1: 41. https://0-doi-org.brum.beds.ac.uk/10.3390/v12010041