Binding Mechanism Elucidation of the Acute Respiratory Disease Causing Agent Adenovirus of Serotype 7 to Desmoglein-2

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prokaryotic Expression Plasmid
2.2. Mammalian Expression Plasmid
2.3. Transient Cell Transfection
2.4. Immobilized-Metal Affinity Chromatography (IMAC) Protein Purification
2.5. Sedimentation Equilibrium by Analytical Ultracentrifugation Experiments (SE-AUC)
2.6. Negative-Stain Electron Microscopy
2.7. Cryo-electron Microscopy
2.8. Image Analysis and 3D Reconstruction
2.9. Model Refinement
2.10. Bio-Layer Interferometry (BLI)
3. Results
3.1. Co-Incubation of HAd7K and EC23 Leads to a Complex Formation
3.2. Biophysical Characterization and Visualization of the HAd7K/EC23 Complex
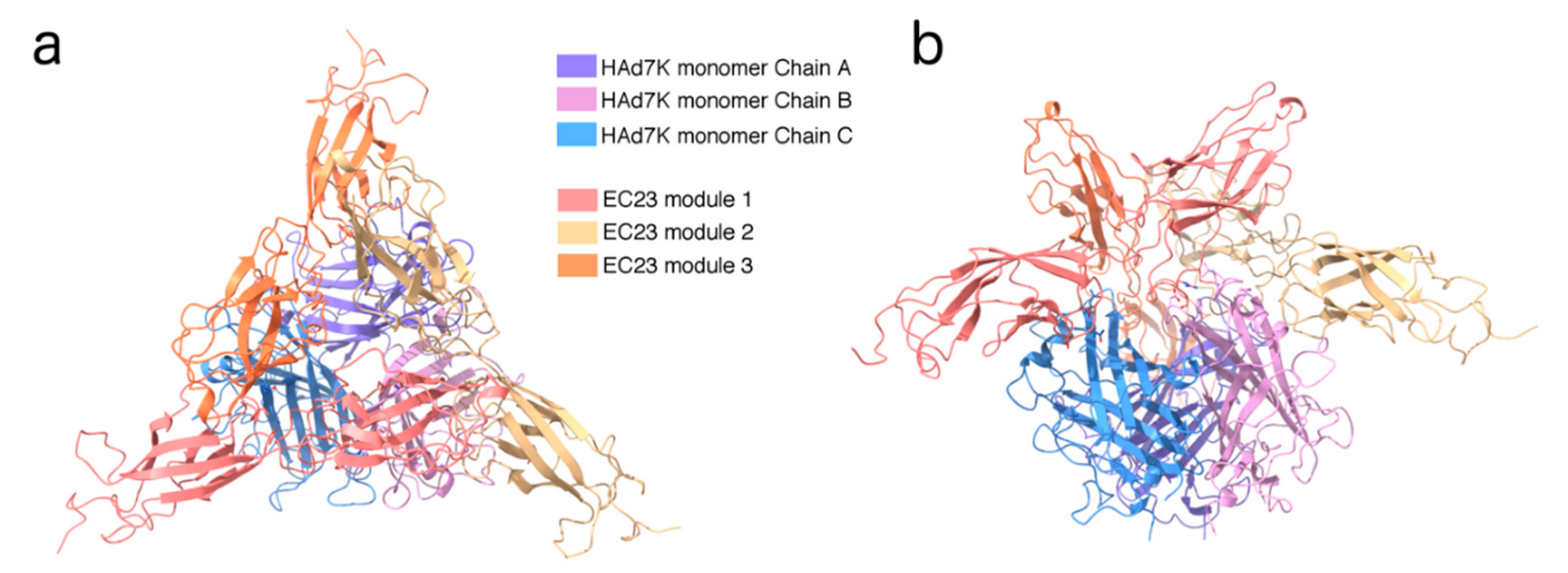
3.3. Structure Determination by Cryo-EM of the HAd7K/EC23 Complex
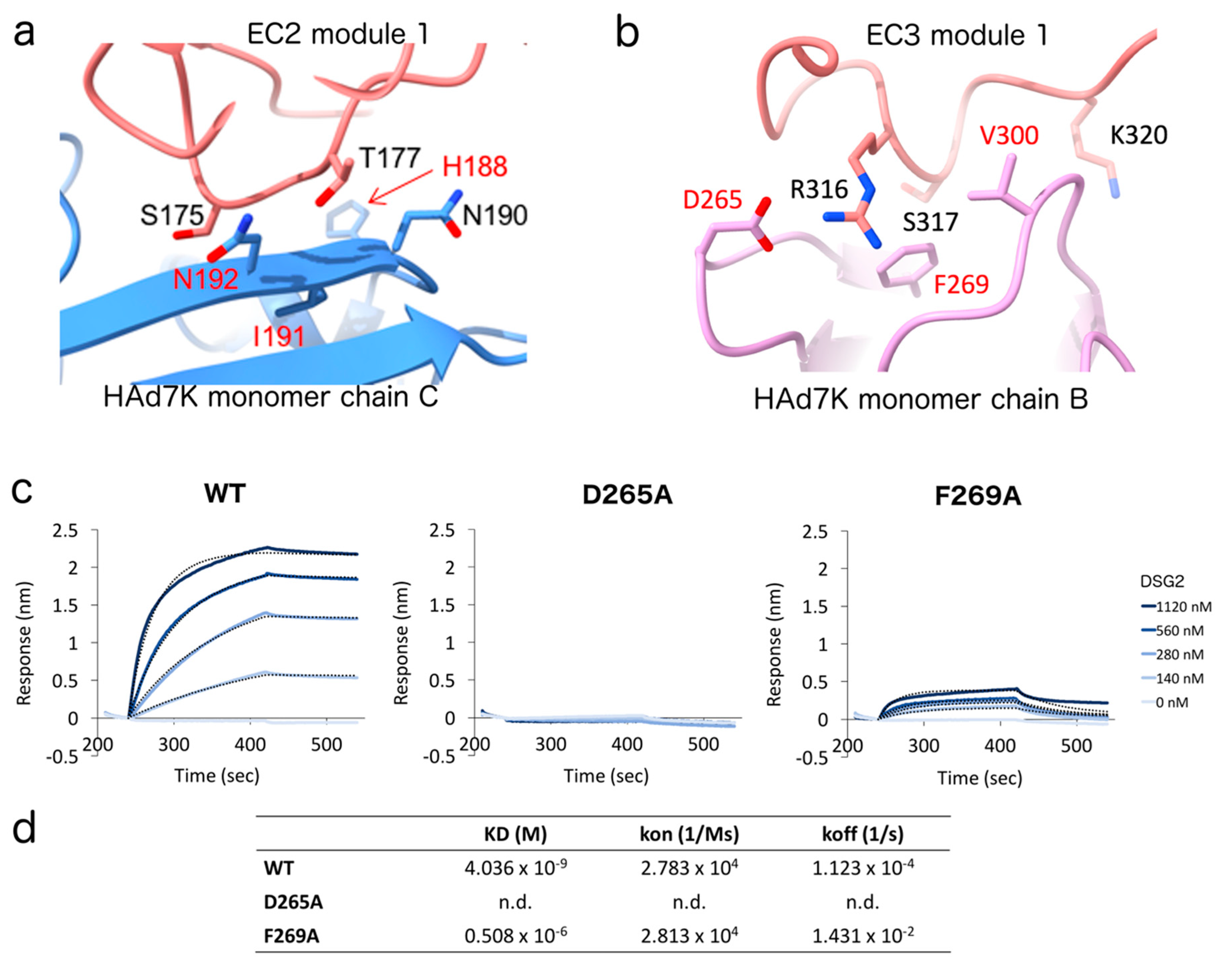
3.4. Identification and Mutagenesis of the Key Player Amino Acids
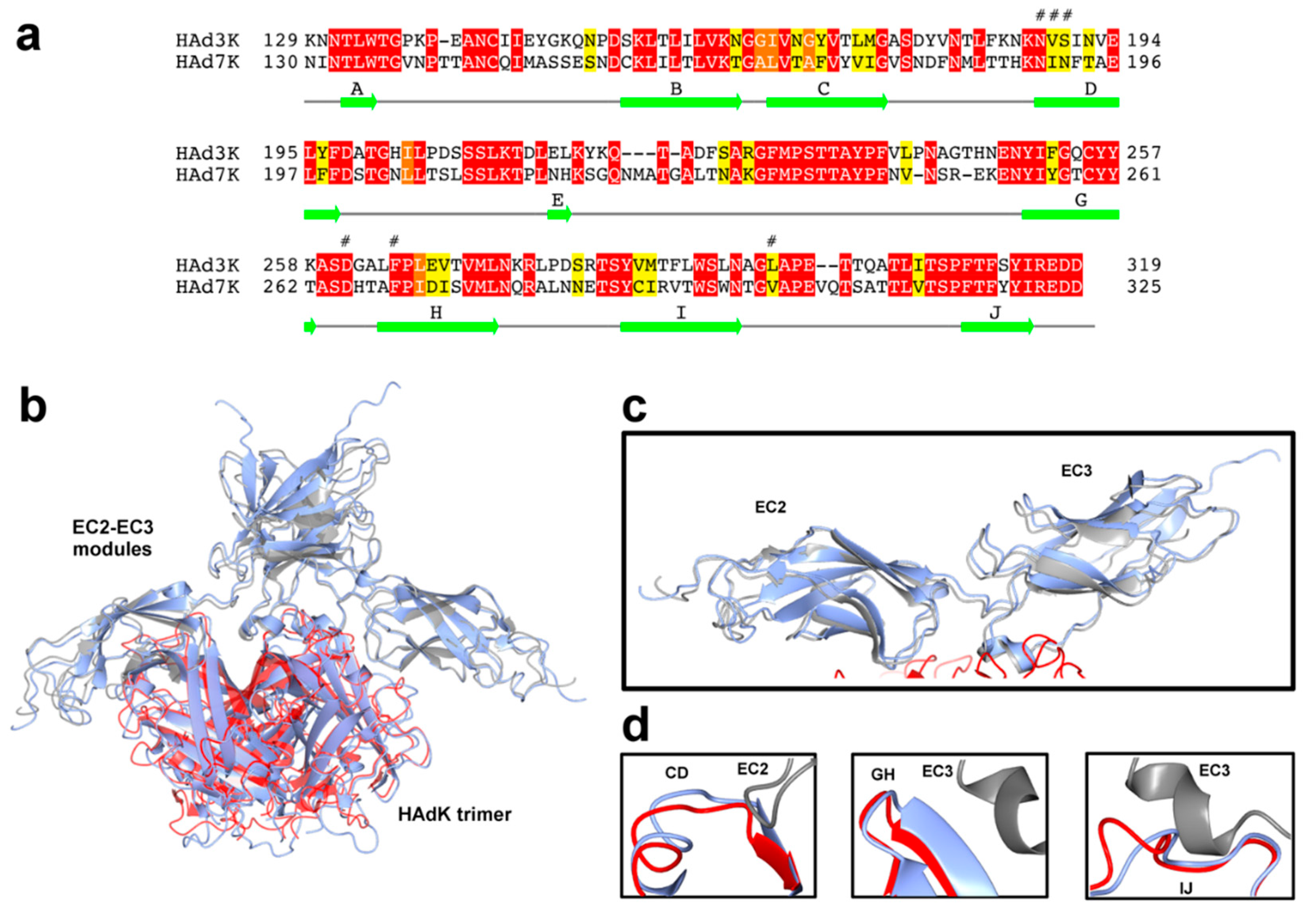
3.5. Comparison of HAd7K and HAd3K Binding to DSG2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
Abbreviations
| ARDS | Acute Respiratory Distress Syndrome |
| ARI | Acute Respiratory Infection |
| BLI | Bio-Layer Interferometry |
| CAR | Coxsackie and Adenovirus Receptor |
| CD46 | Cluster of Differentiation 46 |
| Cryo-EM | Cryo-Electron Microscopy |
| DSG2 | Desmoglein-2 |
| HAdV | Human Adenovirus |
| HAdK | Human Adenovirus Knob |
| EC23 | Extra Cellular domains 2 and 3 (from DSG2) |
| IMAC | Immobilized-Metal Affinity Chromatography |
| RMSD | Root Mean Square Deviation |
References
- Human Adenovirus Working Group. Available online: http://hadvwg.gmu.edu/ (accessed on 23 April 2020).
- Lion, T. Adenovirus Infections in Immunocompetent and Immunocompromised Patients. Clin. Microbiol. Rev. 2014, 27, 441–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Lv, J.; Qin, L.; Zou, C.; Tang, L. Severe Adenovirus Pneumonia Requiring Extracorporeal Membrane Oxygenation Support in Immunocompetent Children. Front. Pediatr. 2020, 8, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-J.; Hong, J.-Y.; Lee, H.-J.; Shin, S.-H.; Kim, Y.K.; Inada, T.; Hashido, M.; Piedra, P.A. Genome Type Analysis of Adenovirus Types 3 and 7 Isolated during Successive Outbreaks of Lower Respiratory Tract Infections in Children. J. Clin. Microbiol. 2003, 41, 4594–4599. [Google Scholar] [CrossRef] [Green Version]
- Kajon, A.E.; Lynch, J.P. Adenovirus: Epidemiology, Global Spread of Novel Serotypes, and Advances in Treatment and Prevention. Semin. Respir. Crit. Care Med. 2016, 37, 586–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauerbrei, A.; Sehr, K.; Eichhorn, U.; Reimer, K.; Wutzler, P. Inactivation of human adenovirus genome by different groups of disinfectants. J. Hosp. Infect. 2004, 57, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.-H.; Lin, Y.-C.; Chen, H.-L.; Ke, G.-M.; Chiang, C.-J.; Hwang, K.-P.; Chu, P.-Y.; Lin, J.-H.; Liu, D.-P.; Chen, H.-Y. A two decade survey of respiratory adenovirus in Taiwan: The reemergence of adenovirus types 7 and 4. J. Med. Virol. 2004, 73, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Kajon, A.E.; Lu, X.; Erdman, D.; Louie, J.; Schnurr, D.; George, K.S.; Koopmans, M.; Allibhai, T.; Metzgar, D. Molecular Epidemiology and Brief History of Emerging Adenovirus 14–Associated Respiratory Disease in the United States. J. Infect. Dis. 2010, 202, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolavic-Gray, S.A.; Binn, L.N.; Sanchez, J.L.; Cersovsky, S.B.; Polyak, C.S.; Mitchell-Raymundo, F.; Asher, L.V.; Vaughn, D.W.; Feighner, B.H.; Innis, B. Large Epidemic of Adenovirus Type 4 Infection among Military Trainees: Epidemiological, Clinical, and Laboratory Studies. Clin. Infect. Dis. 2002, 35, 808–818. [Google Scholar] [CrossRef] [Green Version]
- Crenshaw, B.J.; Jones, L.B.; Bell, C.R.; Kumar, S.; Matthews, Q.L. Perspective on Adenoviruses: Epidemiology, Pathogenicity, and Gene Therapy. Biomedicines 2019, 7, 61. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Wen, L.; Wu, Z.; Liu, N.; Yang, C.; Liu, W.; Ba, Z.; Wang, J.; Yi, S.; Li, H.; et al. Human Adenovirus Type 7 Infection Associated with Severe and Fatal Acute Lower Respiratory Illness and Nosocomial Transmission. J. Clin. Microbiol. 2014, 53, 746–749. [Google Scholar] [CrossRef] [Green Version]
- A Hakim, F.; Tleyjeh, I.M. Severe adenovirus pneumonia in immunocompetent adults: A case report and review of the literature. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 27, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Tang, Z.; Ye, Z.; Mo, S.; Tian, X.; Ni, K.; Ren, L.; Liu, E.; Zang, N. Human adenovirus type 7 infection causes a more severe disease than type 3. BMC Infect. Dis. 2019, 19, 36. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, O.; Meurman, O.; Sarkkinen, H. Adenoviral diseases in children: A study of 105 hospital cases. Pediatrics 1985, 76, 79–83. [Google Scholar] [PubMed]
- Mistchenko, A.S.; Diez, R.A.; La Mariani, A.; Robaldo, J.; Maffey, A.F.; Bayley-Bustamante, G.; Grinstein, S. Cytokines in adenoviral disease in children: Association of interleukin-6, interleukin-8, and tumor necrosis factor alpha levels with clinical outcome. J. Pediatr. 1994, 124, 714–720. [Google Scholar] [CrossRef]
- Ryan, M.A.K.; Gray, G.C.; Smith, B.; McKeehan, J.A.; Hawksworth, A.W.; Malasig, M.D. Large Epidemic of Respiratory Illness Due to Adenovirus Types 7 and 3 in Healthy Young Adults. Clin. Infect. Dis. 2002, 34, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Kajon, A.E.; Mistchenko, A.S.; Videla, C.; Hortal, M.; Wadell, G.; Avendaño, L.F. Molecular epidemiology of adenovirus acute lower respiratory infections of children in the South Cone of South America (1991–1994). J. Med. Virol. 1996, 48, 151–156. [Google Scholar] [CrossRef]
- De Andrade, H.R.; Pereira, C.C.; Gíria, M.; Prudêncio, E.; Brito, M.J.; Calé, E.; Taveira, N. Outbreak of Acute Respiratory Infection among Infants in Lisbon, Portugal, Caused by Human Adenovirus Serotype 3 and a New 7/3 Recombinant Strain. J. Clin. Microbiol. 2010, 48, 1391–1396. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.-Y.; Lee, C.-J.; Lu, C.-Y.; Lee, P.-I.; Shao, P.-L.; Wu, E.-T.; Wang, C.-C.; Tan, B.-F.; Chang, H.-Y.; Hsia, S.-H.; et al. Adenovirus Serotype 3 and 7 Infection with Acute Respiratory Failure in Children in Taiwan, 2010–2011. PLoS ONE 2013, 8, e53614. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wan, C.; Ke, C.; Seto, J.; Dehghan, S.; Zou, L.; Zhou, J.; Cheng, Z.; Jing, S.; Zeng, Z.; et al. Re-emergent Human Adenovirus Genome Type 7d Caused an Acute Respiratory Disease Outbreak in Southern China After a Twenty-one Year Absence. Sci. Rep. 2014, 4, 7365. [Google Scholar] [CrossRef] [Green Version]
- Department of Health|Communicable Disease Service|Adenovirus. Available online: https://www.nj.gov/health/cd/topics/adenovirus.shtml (accessed on 23 April 2020).
- Martinez-Aguado, P.; Serna, A.; Marrugal-Lorenzo, J.A.; Gómez-Marín, I.; Sánchez-Céspedes, J. Antiadenovirus drug discovery: Potential targets and evaluation methodologies. Drug Discov. Today 2015, 20, 1235–1242. [Google Scholar] [CrossRef]
- Ortiz, A.; Justo, P.; Sanz, A.; Melero, R.; Caramelo, C.; Guerrero, M.F.; Strutz, F.; Müller, G.; Barat, A.; Egido, J. Tubular cell apoptosis and cidofovir-induced acute renal failure. Antivir. Ther. 2005, 10, 185–190. [Google Scholar] [PubMed]
- Van Genechten, T.; Van Heerden, J.; Bauters, T.; Dhooge, C. Successful Treatment of Adenovirus Infection with Brincidofovir in an Immunocompromised Patient after Hematological Stem Cell Transplantation. Case Rep. Infect. Dis. 2020, 2020, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acton, Q.A. Food and Drug Administration Adenovirus type 4 and type 7 vaccine, live, oral. In Enteric Coated Tablets for Oral Administration; Scholary Editions: Atlanta, GA, USA, 2012. [Google Scholar]
- Wang, H.; Li, Z.-Y.; Liu, Y.; Persson, J.; Beyer, I.; Möller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.-B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2010, 17, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Vassal-Stermann, E.; Mottet, M.; Ducournau, C.; Iseni, F.; Vragniau, C.; Wang, H.; Zubieta, C.; Lieber, A.; Fender, P. Mapping of Adenovirus of serotype 3 fibre interaction to desmoglein 2 revealed a novel ’non-classical’ mechanism of viral receptor engagement. Sci. Rep. 2018, 8, 8381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassal-Stermann, E.; Effantin, G.; Zubieta, C.; Burmeister, W.P.; Iseni, F.; Wang, H.; Lieber, A.; Schoehn, G.; Fender, P. CryoEM structure of adenovirus type 3 fibre with desmoglein 2 shows an unusual mode of receptor engagement. Nat. Commun. 2019, 10, 1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, A.T.; Mundy, R.M.; Davies, J.; Rizkallah, P.J.; Parker, A.L. Human adenovirus type 26 uses sialic acid–bearing glycans as a primary cell entry receptor. Sci. Adv. 2019, 5, eaax3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bewley, M.C.; Vogel, K.S.; Klesse, L.J.; Velasco-Miguel, S.; Meyers, K.; Rushing, E.J.; Parada, L.F. Structural Analysis of the Mechanism of Adenovirus Binding to Its Human Cellular Receptor, CAR. Science 1999, 286, 1579–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burmeister, W.P.; Guilligay, D.; Cusack, S.; Wadell, G.; Arnberg, N. Crystal Structure of Species D Adenovirus Fiber Knobs and Their Sialic Acid Binding Sites. J. Virol. 2004, 78, 7727–7736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, B.D.; Reiter, D.M.; Marttila, M.; Mei, Y.-F.; Casasnovas, J.M.; Arnberg, N.; Stehle, T. Adenovirus type 11 binding alters the conformation of its receptor CD46. Nat. Struct. Mol. Boil. 2007, 14, 164–166. [Google Scholar] [CrossRef]
- Vassal-Stermann, E.; Hutin, S.; Fender, P.; Burmeister, W.P. Intermediate-resolution crystal structure of the human adenovirus B serotype 3 fibre knob in complex with the EC2-EC3 fragment of desmoglein 2. Acta Cryst. Sect. F Struct. Boil. Commun. 2019, 75, 750–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, R.C.; Shapiro, B.M.; Stadtman, E.R. Regulation of glutamine synthetase. XII. Electron microscopy of the enzyme from Escherichia coli. Biochemistry 1968, 7, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Scheres, S.H.W. RELION: Implementation of a Bayesian approach to cryo-EM structure determination. J. Struct. Boil. 2012, 180, 519–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandiah, E.; Giraud, T.; Antolinos, A.D.M.; Dobias, F.; Effantin, G.; Flot, D.; Hons, M.; Schoehn, G.; Susini, J.; Svensson, O.; et al. CM01: A facility for cryo-electron microscopy at the European Synchrotron. Acta Cryst. Sect. D Struct. Boil. 2019, 75, 528–535. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.Q.; Palovcak, E.; Armache, J.-P.; Verba, K.A.; Cheng, Y.; Agard, D.A. MotionCor2: Anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 2017, 14, 331–332. [Google Scholar] [CrossRef] [Green Version]
- Kimanius, D.; Forsberg, B.O.; Scheres, S.H.W.; Lindahl, E. Accelerated cryo-EM structure determination with parallelisation using GPUs in RELION-2. eLife 2016, 5, e18722. [Google Scholar] [CrossRef] [PubMed]
- Zivanov, J.; Nakane, T.; Forsberg, B.O.; Kimanius, D.; Hagen, W.; Lindahl, E.; Scheres, S.H.W. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 2018, 7. [Google Scholar] [CrossRef]
- Zhang, K. Gctf: Real-time CTF determination and correction. J. Struct. Boil. 2016, 193, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Persson, B.D.; Müller, S.; Reiter, D.M.; Schmitt, B.B.T.; Marttila, M.; Sumowski, C.V.; Schweizer, S.; Scheu, U.; Ochsenfeld, C.; Arnberg, N.; et al. An Arginine Switch in the Species B Adenovirus Knob Determines High-Affinity Engagement of Cellular Receptor CD46. J. Virol. 2008, 83, 673–686. [Google Scholar] [CrossRef] [Green Version]
- Harrison, O.J.; Brasch, J.; Lasso, G.; Katsamba, P.; Ahlsén, G.; Honig, B.; Shapiro, L. Structural basis of adhesive binding by desmocollins and desmogleins. Proc. Natl. Acad. Sci. USA 2016, 113, 7160–7165. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.Y.-R.; Song, Y.; Barad, B.A.; Cheng, Y.; Fraser, J.S.; DiMaio, F. Automated structure refinement of macromolecular assemblies from cryo-EM maps using Rosetta. eLife 2016, 5, e17219. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. Sect. D Boil. Cryst. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Cryst. Sect. D Boil. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2017, 27, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of Macromolecular Assemblies from Crystalline State. J. Mol. Boil. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2017, 27, 14–25. [Google Scholar] [CrossRef]
- Aricescu, A.R.; Lu, W.; Jones, E.Y. A time- and cost-efficient system for high-level protein production in mammalian cells. Acta Cryst. Sect. D Boil. Cryst. 2006, 62, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yumul, R.; Cao, H.; Ran, L.; Fan, X.; Richter, M.; Epstein, F.; Gralow, J.; Zubieta, C.; Fender, P.; et al. Structural and Functional Studies on the Interaction of Adenovirus Fiber Knobs and Desmoglein 2. J. Virol. 2013, 87, 11346–11362. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bergelson, J.M. Adenovirus Receptors. J. Virol. 2005, 79, 12125–12131. [Google Scholar] [CrossRef] [Green Version]
- Purkayastha, A.; Su, J.; Carlisle, S.; Tibbetts, C.; Seto, D. Genomic and bioinformatics analysis of HAdV-7, a human adenovirus of species B1 that causes acute respiratory disease: Implications for vector development in human gene therapy. Virology 2005, 332, 114–129. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.T.; Greenshields-Watson, A.; Coughlan, L.; Davies, J.; Uusi-Kerttula, H.; Cole, D.K.; Rizkallah, P.J.; Parker, A.L. Diversity within the adenovirus fiber knob hypervariable loops influences primary receptor interactions. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, T.O.; England, B.; Mauris, C.; Shuey, H.E.; Lennette, E.H. Etiology of acute respiratory disease among service personnel at fort ord, california. Am. J. Epidemiol. 1955, 62, 283–294. [Google Scholar] [CrossRef] [PubMed]
- Kajon, A.E.; Wadell, G. Sequence Analysis of the E3 Region and Fiber Gene of Human Adenovirus Genome Type 7h. Virology 1996, 215, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hograindleur, M.-A.; Effantin, G.; Fenel, D.; Mas, C.; Lieber, A.; Schoehn, G.; Fender, P.; Vassal-Stermann, E. Binding Mechanism Elucidation of the Acute Respiratory Disease Causing Agent Adenovirus of Serotype 7 to Desmoglein-2. Viruses 2020, 12, 1075. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101075
Hograindleur M-A, Effantin G, Fenel D, Mas C, Lieber A, Schoehn G, Fender P, Vassal-Stermann E. Binding Mechanism Elucidation of the Acute Respiratory Disease Causing Agent Adenovirus of Serotype 7 to Desmoglein-2. Viruses. 2020; 12(10):1075. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101075
Chicago/Turabian StyleHograindleur, Marc-André, Gregory Effantin, Daphna Fenel, Caroline Mas, André Lieber, Guy Schoehn, Pascal Fender, and Emilie Vassal-Stermann. 2020. "Binding Mechanism Elucidation of the Acute Respiratory Disease Causing Agent Adenovirus of Serotype 7 to Desmoglein-2" Viruses 12, no. 10: 1075. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101075