An Outbreak of a Respiratory Disorder at a Russian Swine Farm Associated with the Co-Circulation of PRRSV1 and PRRSV2

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. ELISA
2.3. PCR and Sequencing
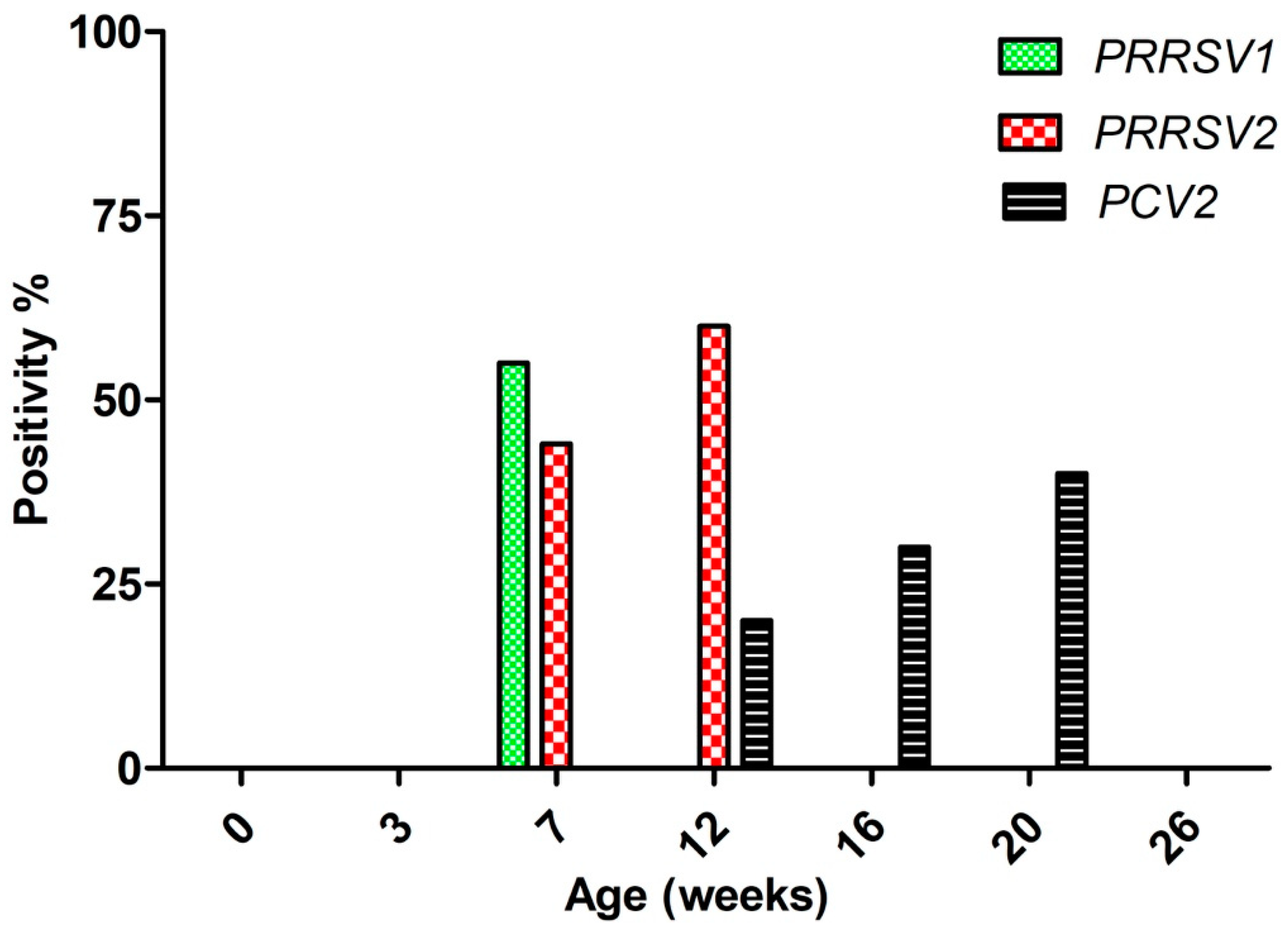
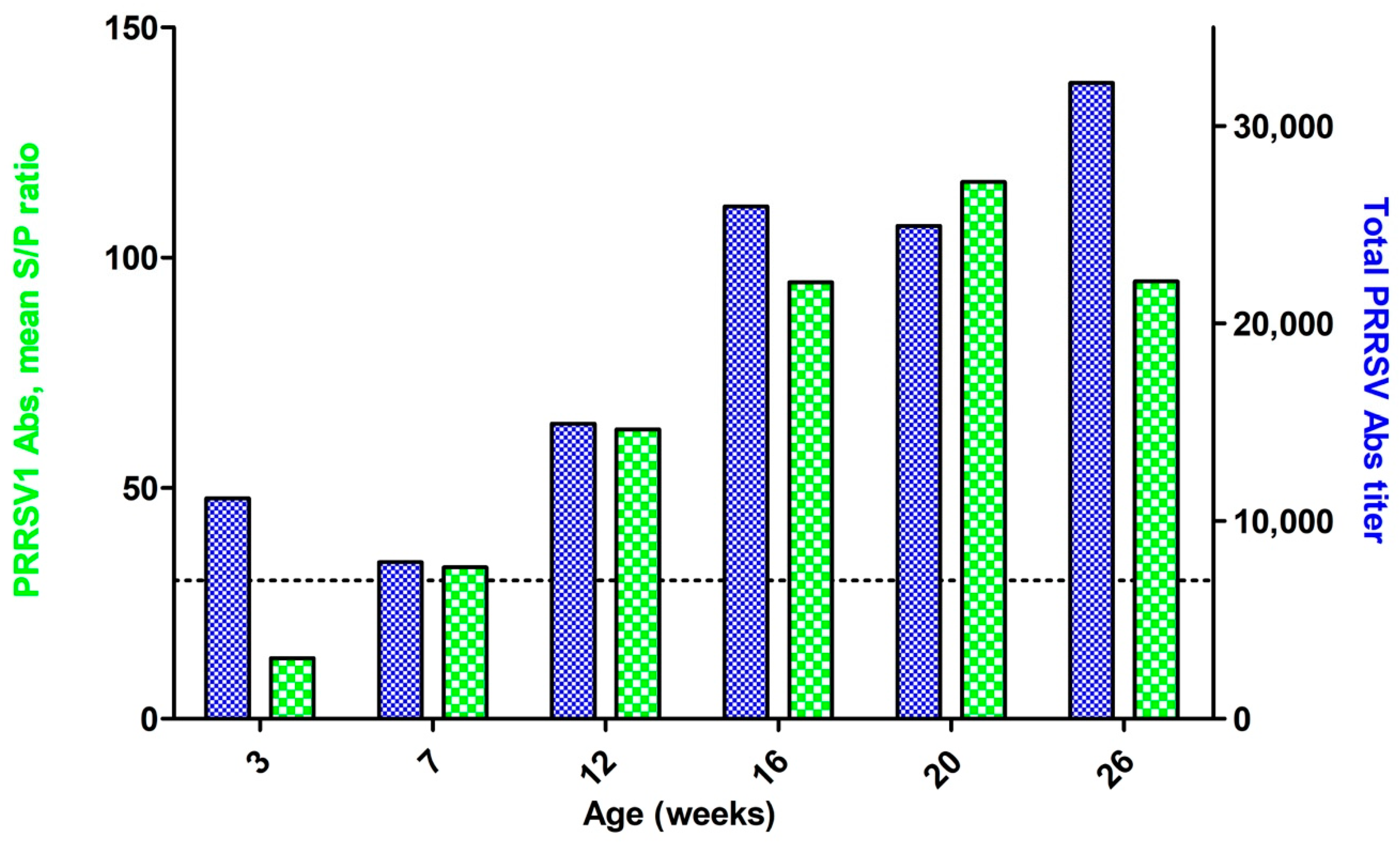
3. Results
3.1. Viremial ELISA
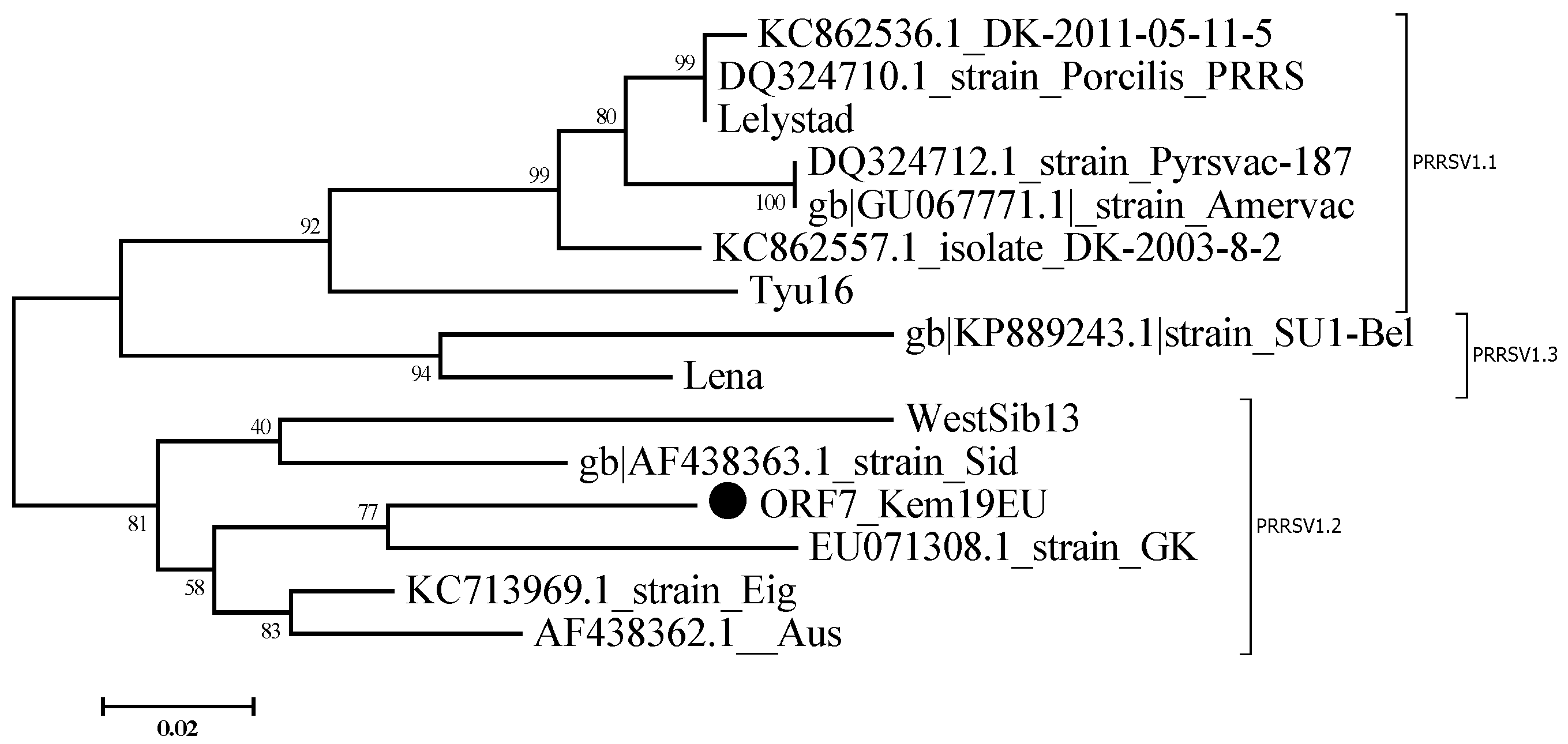
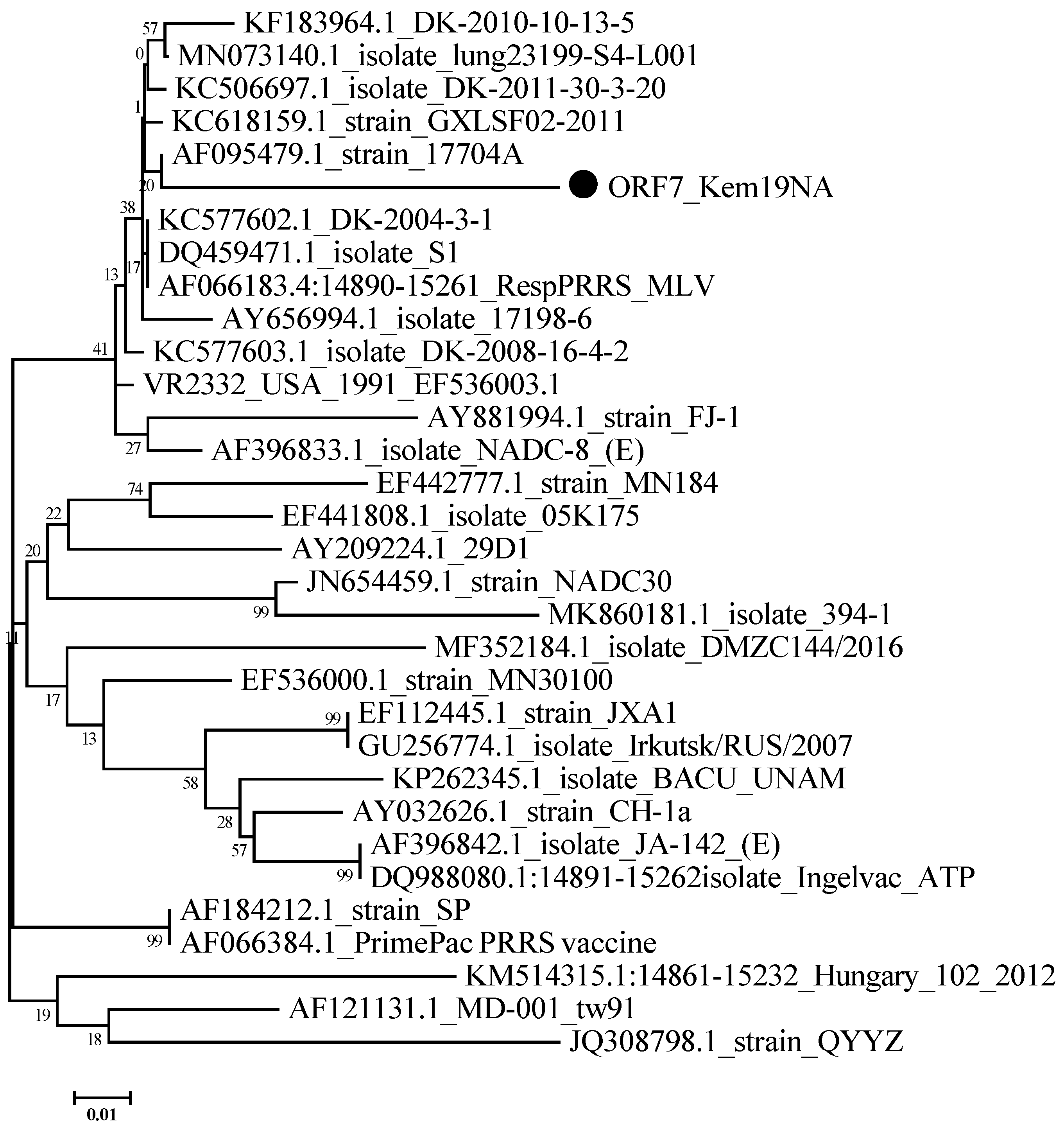
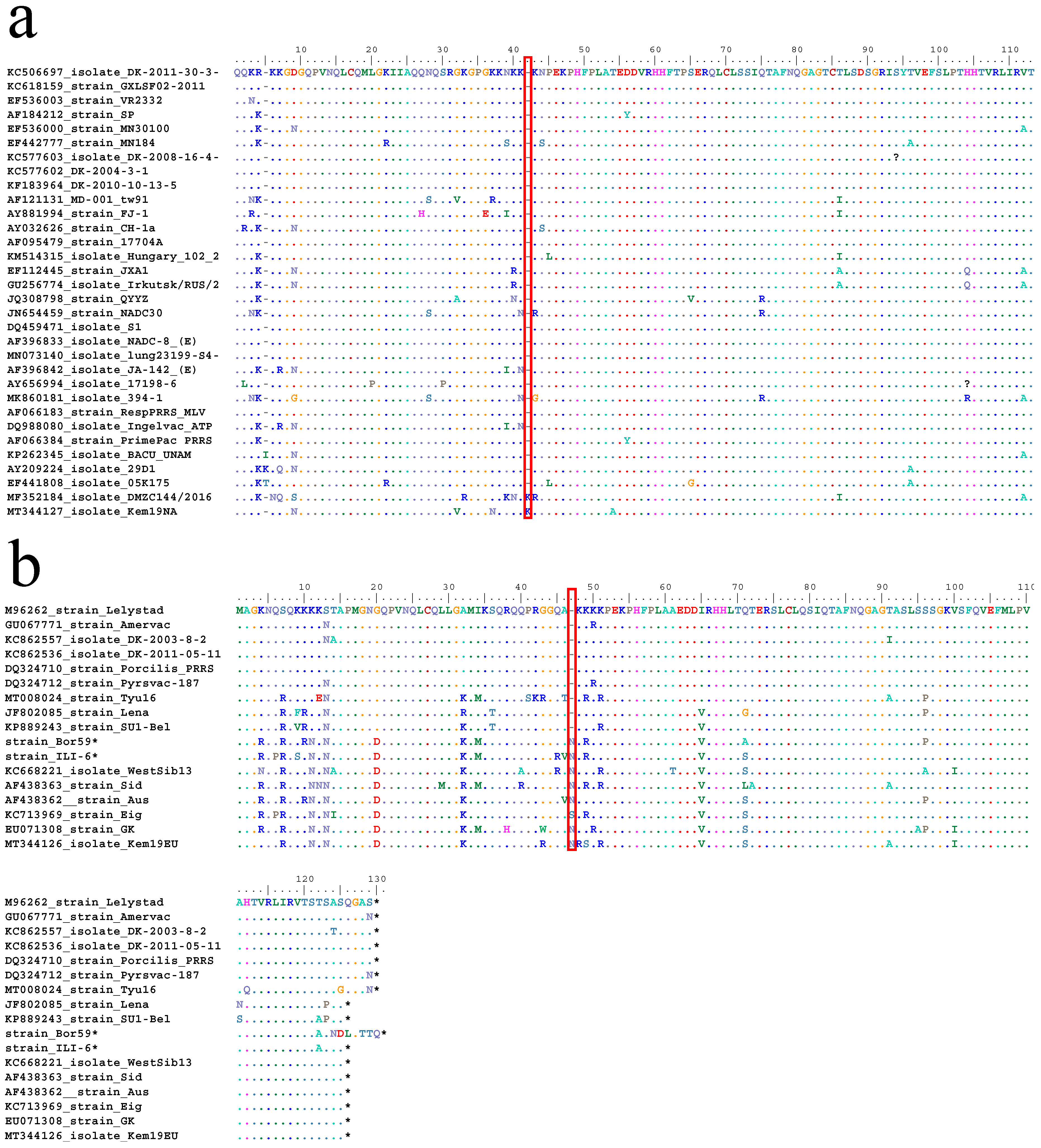
3.2. Sequencing Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Done, S.H.; Paton, D.J. Porcine reproductive and respiratory syndrome: Clinical disease, pathology and immunosuppression. Vet. Rec. 1995, 136, 32–35. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV)Release EC 50, Washington, DC, July 2018 Email Ratification February 2019 (MSL #34). Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=201851832 (accessed on 16 April 2020).
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998, 79, 961–979. [Google Scholar] [CrossRef]
- Balka, G.; Hornyák, Á.; Bálint, Á.; Kiss, I.; Kecskeméti, S.; Bakonyi, T.; Rusvai, M. Genetic diversity of porcine reproductive and respiratory syndrome virus strains circulating in Hungarian swine herds. Vet. Microbiol. 2008, 127, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Dewey, C.; Charbonneau, G.; Carman, S.; Hamel, A.; Nayar, G.; Friendship, R.; Eernisse, K.; Swenson, S. Lelystad-like strain of porcine reproductive and respiratory syndrome virus (PRRSV) identified in Canadian swine. Can. Vet. J. La Rev. Veter- Can. 2000, 41, 493–494. [Google Scholar]
- Ropp, S.L.; Wees, C.E.M.; Fang, Y.; Nelson, E.A.; Rossow, K.D.; Bien, M.; Arndt, B.; Preszler, S.; Steen, P.; Christopher-Hennings, J.; et al. Characterization of Emerging European-Like Porcine Reproductive and Respiratory Syndrome Virus Isolates in the United States. J. Virol. 2004, 78, 3684–3703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Lam, T.T.-Y.; Hon, C.-C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.-H.; Li, J.; Wong, L.T.-W.; Yip, C.-W.; Jiang, J.-W.; et al. Phylogeny-Based Evolutionary, Demographical, and Geographical Dissection of North American Type 2 Porcine Reproductive and Respiratory Syndrome Viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef] [Green Version]
- Stadejek, T.; Oleksiewicz, M.B.; Scherbakov, A.V.; Timina, A.M.; Krabbe, J.S.; Chabros, K.; Potapchuk, D. Definition of subtypes in the European genotype of porcine reproductive and respiratory syndrome virus: Nucleocapsid characteristics and geographical distribution in Europe. Arch. Virol. 2008, 153, 1479–1488. [Google Scholar] [CrossRef]
- Stadejek, T.; Stankevicius, A.; Murtaugh, M.P.; Oleksiewicz, M.B. Molecular evolution of PRRSV in Europe: Current state of play. Vet. Microbiol. 2013, 165, 21–28. [Google Scholar] [CrossRef]
- Kang, H.; Yu, J.E.; Shin, J.-E.; Kang, A.; Kim, W.-I.; Lee, C.; Lee, J.; Cho, I.-S.; Choe, S.-E.; Cha, S.-H. Geographic distribution and molecular analysis of porcine reproductive and respiratory syndrome viruses circulating in swine farms in the Republic of Korea between 2013 and 2016. BMC Vet. Res. 2018, 14, 160. [Google Scholar] [CrossRef] [Green Version]
- Yuzhakov, A.G.; Raev, S.A.; Shchetinin, A.M.; Gushchin, V.A.; Alekseev, K.P.; Stafford, V.V.; Komina, A.K.; Zaberezhny, A.D.; Gulyukin, A.M.; Aliper, T.I. Full-genome analysis and pathogenicity of a genetically distinct Russian PRRSV-1 Tyu16 strain. Vet. Microbiol. 2020, 247, 108784. [Google Scholar] [CrossRef]
- Yuzhakov, A.G.; Raev, S.A.; Skrylev, A.N.; Mishin, A.M.; Grebennikova, T.V.; Verkhovsky, O.A.; Zaberezhny, A.D.; Trus, I.; Nauwynck, H.J.; Aliper, T.I. Genetic and pathogenic characterization of a Russian subtype 2 PRRSV-1 isolate. Vet. Microbiol. 2017, 211, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Kukushkin, S.A.; Baybikov, T.Z.; Scherbakov, A.V.; Timina, A.M.; Baborenko, E.P.; Puzankova, O.P.; Pronin, I.A.; Fomin, A.E. First Outbreak of Atypical Porcine Reproductive and Respiratory Syndrome in Russia Caused by Highly Pathogenic Chinese-Like Prrs Virus. NA PRRSV Symp. 2008, 2008, 23–26. [Google Scholar]
- Storgaard, T.; Oleksiewicz, M.; Bøtner, A. Examination of the selective pressures on a live PRRS vaccine virus. Arch. Virol. 1999, 144, 2389–2401. [Google Scholar] [CrossRef]
- Sørensen, K. Blocking ELISA’s for the distinction between antibodies against European and American strains of porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 1998, 60, 169–177. [Google Scholar] [CrossRef]
- Choi, K.; Lee, J.; Park, C.; Jeong, J.; Chae, C. Comparison of the Pathogenesis of Single or Dual Infections with Type 1 and Type 2 Porcine Reproductive and Respiratory Syndrome Virus. J. Comp. Pathol. 2015, 152, 317–324. [Google Scholar] [CrossRef]
- Bøtner, A.; Strandbygaard, B.; Sorensen, K.J.; Have, P.; Madsen, K.G.; Madsen, E.S.; Alexandersen, S. Appearance of acute PRRS-like symptoms in sow herds after vaccination with a modified live PRRS vaccine. Vet. Rec. 1997, 141, 497–499. [Google Scholar] [CrossRef]
- Opriessnig, T.; Halbur, P.G.; Yoon, K.-J.; Pogranichniy, R.M.; Harmon, K.M.; Evans, R.; Key, K.F.; Pallares, F.J.; Thomas, P.; Meng, X.J. Comparison of Molecular and Biological Characteristics of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Vaccine (Ingelvac PRRS MLV), the Parent Strain of the Vaccine (ATCC VR2332), ATCC VR2385, and Two Recent Field Isolates of PRRSV. J. Virol. 2002, 76, 11837–11844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karniychuk, U.; Geldhof, M.; Vanhee, M.; Doorsselaere, J.V.; A Saveleva, T.; Nauwynck, H.J. Pathogenesis and antigenic characterization of a new East European subtype 3 porcine reproductive and respiratory syndrome virus isolate. BMC Vet. Res. 2010, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadejek, T.; Larsen, L.E.; Podgórska, K.; Bøtner, A.; Botti, S.; Dolka, I.; Fabisiak, M.; Heegaard, P.M.; Hjulsager, C.K.; Huc, T.; et al. Pathogenicity of three genetically diverse strains of PRRSV Type 1 in specific pathogen free pigs. Vet. Microbiol. 2017, 209, 13–19. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.T.-D.; Nguyen, T.T.; Le, H.T.-T.; Nguyen, S.G.; Vo, H.K.; Do, K.V.-A. Genetic Analysis of ORF5 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Isolated in Vietnam. Microbiol. Immunol. 2013, 57, 518–526. [Google Scholar] [CrossRef] [Green Version]
- Rowland, R.; Kervin, R.; Kuckleburg, C.; Sperlich, A.; Benfield, D.A. The localization of porcine reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of infected cells and identification of a potential nucleolar localization signal sequence. Virus Res. 1999, 64, 1–12. [Google Scholar] [CrossRef]
- Lee, C.; Hodgins, D.; Calvert, J.G.; Welch, S.-K.W.; Jolie, R.; Yoo, D. Mutations within the nuclear localization signal of the porcine reproductive and respiratory syndrome virus nucleocapsid protein attenuate virus replication. Virology 2006, 346, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Bautista, N.R.; Sciutto-Conde, E.; Cervantes-Torres, J.; Segura-Velazquez, R.; García, M.C.M.; Ramírez-Mendoza, H.; Ortega, M.E.T.; Alvarez, J.D.; Castillo-Juárez, H.; Sánchez-Betancourt, J.I. Phylogenetic analysis of ORF5 and ORF7 of porcine reproductive and respiratory syndrome (PRRS) virus and the frequency of wild-type PRRS virus in México. Transbound. Emerg. Dis. 2018, 65, 993–1008. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Animal # | Antibody Detection (ELISA) | Viremia Detection (PCR) | ||
|---|---|---|---|---|---|
| PRRSV1 | Pan-PRRSV | PRRSV1 | PRRSV2 | ||
| 3 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
| 7 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
| 9 | |||||
| 12 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
| 9 | |||||
| 10 | |||||
| 16 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
| 9 | |||||
| 10 | |||||
| 20 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
| 9 | |||||
| 10 | |||||
| 26 | 1 | ||||
| 2 | |||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
| 6 | |||||
| 7 | |||||
| 8 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raev, S.; Yuzhakov, A.; Bulgakov, A.; Kostina, L.; Gerasianinov, A.; Verkhovsky, O.; Zaberezhny, A.; Aliper, T. An Outbreak of a Respiratory Disorder at a Russian Swine Farm Associated with the Co-Circulation of PRRSV1 and PRRSV2. Viruses 2020, 12, 1169. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101169
Raev S, Yuzhakov A, Bulgakov A, Kostina L, Gerasianinov A, Verkhovsky O, Zaberezhny A, Aliper T. An Outbreak of a Respiratory Disorder at a Russian Swine Farm Associated with the Co-Circulation of PRRSV1 and PRRSV2. Viruses. 2020; 12(10):1169. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101169
Chicago/Turabian StyleRaev, Sergei, Anton Yuzhakov, Alexandr Bulgakov, Ludmila Kostina, Alexei Gerasianinov, Oleg Verkhovsky, Alexei Zaberezhny, and Taras Aliper. 2020. "An Outbreak of a Respiratory Disorder at a Russian Swine Farm Associated with the Co-Circulation of PRRSV1 and PRRSV2" Viruses 12, no. 10: 1169. https://0-doi-org.brum.beds.ac.uk/10.3390/v12101169