ART-Treated Patients Exhibit an Adaptive Immune Response against the HFVAC Peptides, a Potential HIV-1 Therapeutic Vaccine (Provir/Latitude45 Study)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. ELISPOT and Flow Cytometry
Flow Cytometry
2.3. Statistical Analysis
3. Results
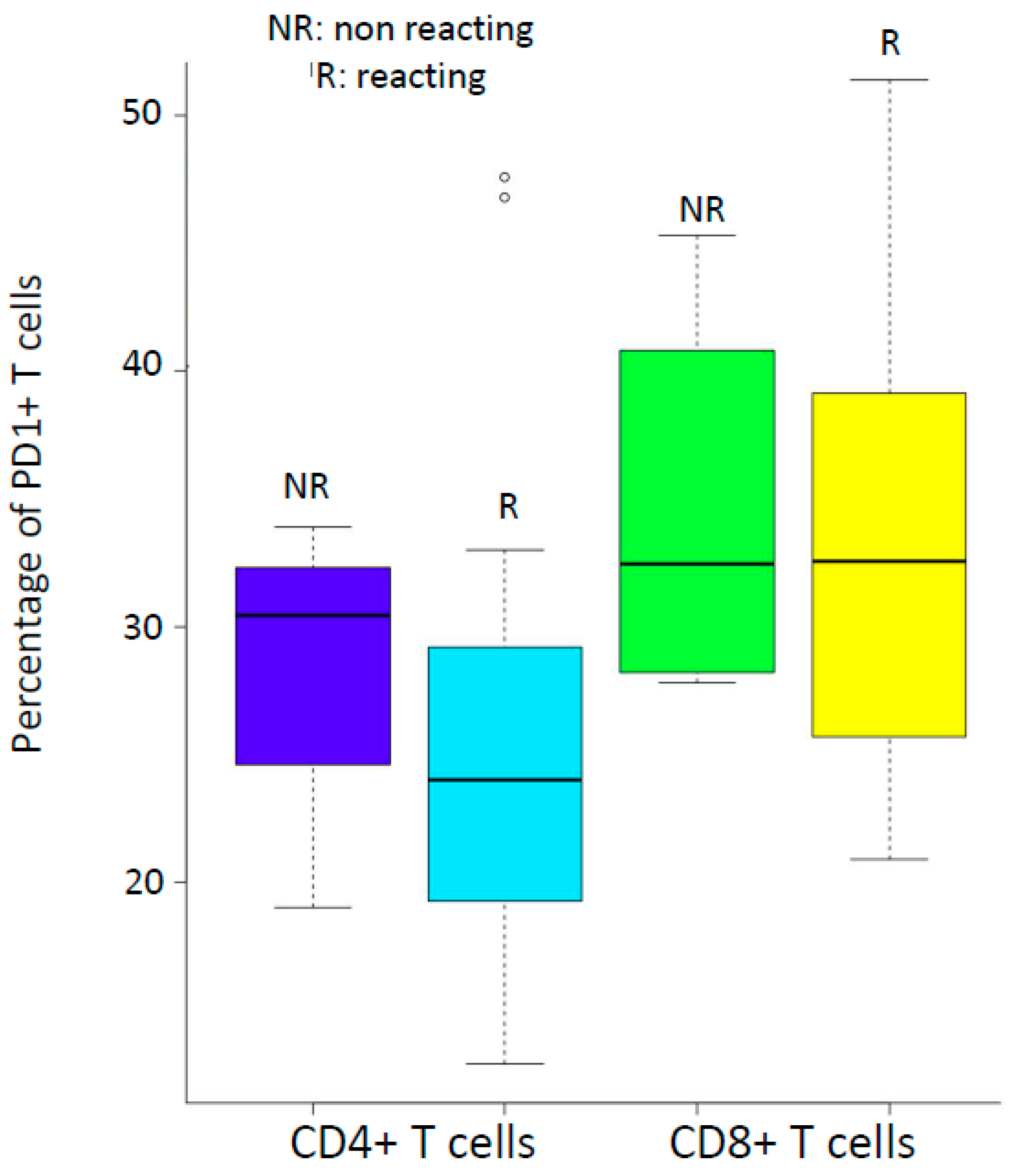
3.1. Patient Characteristics, Lymphocyte Immunophenotyping and PD-1 Expression
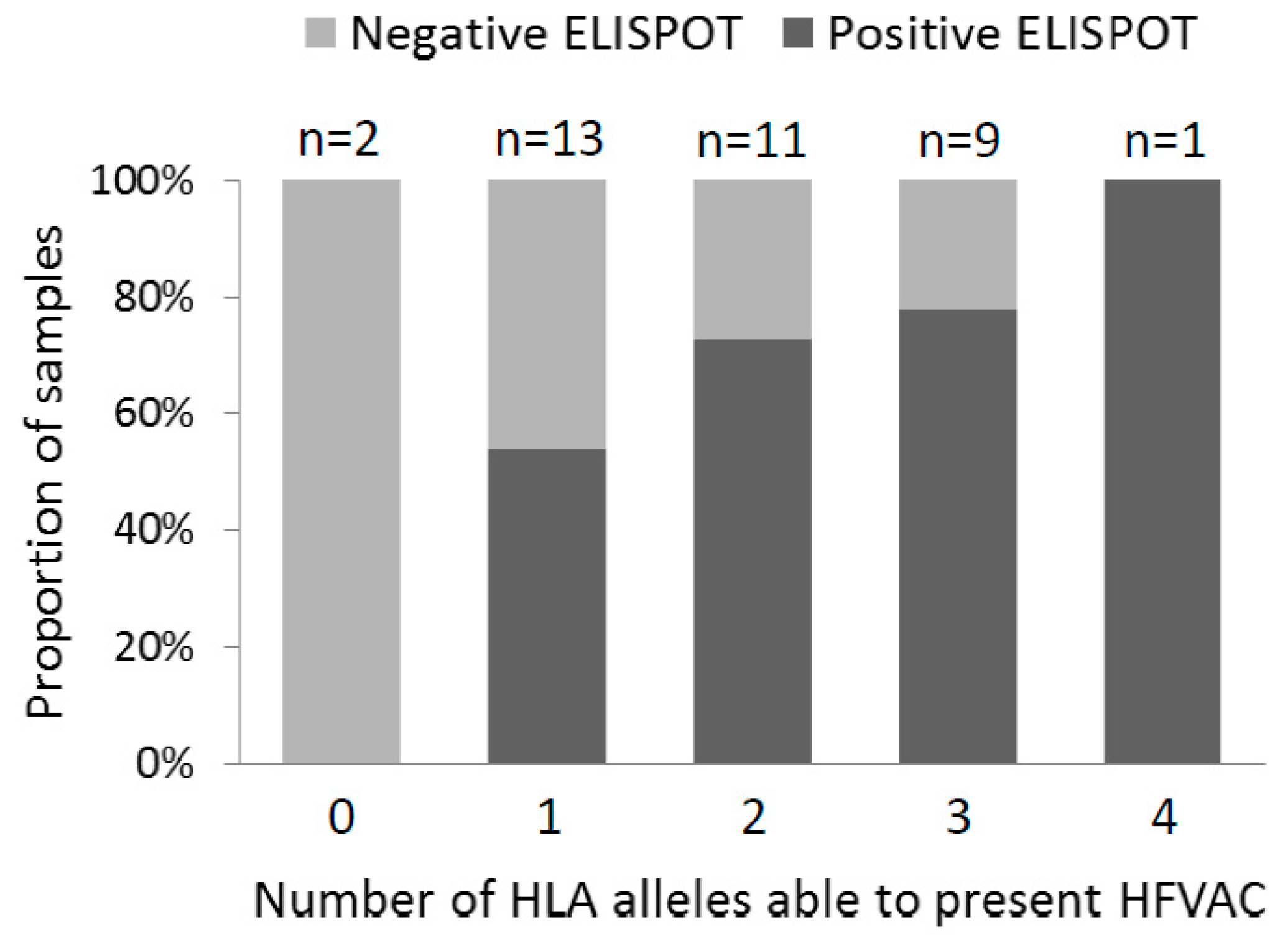
3.2. ELISPOT Reactivity According to HLA Alleles, Archived Viral Sequences and PD-1 Expression
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Salk, J.; Bretscher, P.A.; Salk, P.L.; Clerici, M.; Shearer, G.M. A strategy for prophylactic vaccination against HIV. Science 1993, 28, 1270–1272. [Google Scholar] [CrossRef] [PubMed]
- Fleury, H.; Tumiotto, C.; Bellecave, P.; Recordon-Pinson, P. Therapeutic Vaccine Against HIV, Viral Variability, Cytotoxic T Lymphocyte Epitopes, and Genetics of Patients. AIDS Res. Hum. Retrovir. 2018, 34, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Pollard, R.B.; Rockstroh, J.K.; Pantaleo, G.; Asmuth, D.M.; Peters, B.; Lazzarin, A.; Garcia, F.; Ellefsen, K.; Podzamczer, D.; Van Lunzen, J.; et al. Safety and efficacy of the peptide-based therapeutic vaccine for HIV-1, Vacc-4x: A phase 2 randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2014, 14, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Rockstroh, J.K.; Asmuth, D.; Pantaleo, P.; Clotet, C.; Podzamczer, D.; Van Lunzen, J.; Arastéh, K.; Mitsuyasu, R.; Peters, B.; Silvia, N.; et al. Re-boost immunizations with the peptide-based therapeutic HIV vaccine, Vacc-4x, restores geometric mean viral load set-point during treatment interruption. PLoS ONE 2019, 14, e0210965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gahéry-Ségard, H.; Pialoux, G.; Charmeteau, B.; Sermet, S.; Poncelet, H.; Raux, M.; Tartar, A.; Lévy, J.P.; Gras-Masse, H.; Guillet, J.G. Multiepitopic B- and T-cell responses induced in humans by a human immunodeficiency virus type 1 lipopeptide vaccine. J. Virol. 2000, 74, 1694–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, K.; Pertea, M.; Rongvaux, A.; Wang, L.; Durand, C.M.; Ghiaur, G.; Lai, J.; McHugh, H.L.; Hao, H.; Zhang, H.; et al. Broad CTL response is required to clear latent HIV-1 due to dominance of escape mutations. Nature 2015, 517, 381–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tumiotto, C.; Alves, B.M.; Recordon-Pinson, P.; Jourdain, M.; Bellecave, P.; Guidicelli, G.L.; Visentin, J.; Bonnet, F.; Hessamfar, M.; Neau, D.; et al. Provir/Latitude 45 study: A step towards a multi-epitopic CTL vaccine designed on archived HIV-1 DNA and according to dominant HLA I alleles. PLoS ONE 2019, 14, e0212347. [Google Scholar] [CrossRef] [PubMed]
- Russell, N.D.; Hudgens, M.G.; Ha, R.; Havenar-Daughton, C.; McElrath, M.J. Moving to human immunodeficiency virus type 1 vaccine efficacy trials: Defining T cell responses as potential correlates of immunity. J. Infect. Dis. 2003, 187, 226–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, L.; Guardo, A.C.; Morón-López, S.; Salgado, M.; Mothe, M.; Heirman, C.; Pannus, P.; Vanham, G.; van den Ham, H.J.; Gruters, R.; et al. Phase I clinical trial of an intranodally administered mRNA-based therapeutic vaccine against HIV-1 infection. AIDS 2018, 32, 2533–2545. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.; Leal, L.; Buyze, J.; Pannus, P.; Guardo, A.; Salgado, M.; Mothe, B.; Molto, J.; Moron-Lopez, S.; Galvez, C.; et al. Therapeutic Vaccine in Chronically HIV-1-Infected Patients: A Randomized, Double-Blind, Placebo-Controlled Phase IIa Trial with HTI-TriMix. Vaccines 2019, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Pantaleo, G.; Tapia, G.; Sanchez, B.; Zhang, L.; Trondsen, M.; Hovden, A.-O.; Pollard, R.; Rockstroh, J.; Ökvist, M.; et al. Cell-Mediated Immune Predictors of Vaccine Effect on Viral Load and CD4 Count in a Phase 2 Therapeutic HIV-1 Vaccine Clinical Trial. EBioMedicine 2017, 24, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Mothe, B.; Manzardo, C.; Sanchez-Bernabeu, A.; Coll, P.; Morón-López, S.; Puertas, M.C.; Rosas-Umbert, M.; Cobarsi, P.; Escrig, R.; Jose, M.; et al. Therapeutic Vaccination Refocuses T-cell Responses Towards Conserved Regions of HIV-1 in Early Treated Individuals (BCN 01 study). EClinicalMedicine 2019, 11, 65–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylvaganam, G.; Yanez, A.G.; Maus, M.; Walker, B.D. Toward T Cell-Mediated Control or Elimination of HIV Reservoirs: Lessons From Cancer Immunology. Front. Immunol. 2019, 10, 2109. [Google Scholar] [CrossRef] [PubMed]
- Day, C.L.; Kaufmann, D.E.; Kiepiela, P.; Brown, J.A.; Moodley, E.S.; Reddy, S.; Mackey, E.W.; Miller, J.D.; Leslie, A.J.; DePierres, C.; et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Booiman, T.; Wit, F.W.; Girigorie, A.F.; Maurer, M.; De Francesco, D.; Sabin, C.A.; Harskamp, A.M.; Prins, M.; Franceschi, C.; Deeks, S.G.; et al. Terminal differentiation of T cells is strongly associated with CMV infection and increased in HIV-positive individuals on ART and lifestyle matched controls. PLoS ONE 2017, 12, e0183357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer-Olson, D.; Brady, K.W.; Bartman, M.T.; O’Sullivan, K.M.; Simons, B.C.; Conrad, J.A.; Duncan, C.B.; Lorey, S.; Siddique, A.; Draenert, R.; et al. Fluctuations of functionally distinct CD8+ T-cell clonotypes demonstrate flexibility of the HIV-specific TCR repertoire. Blood 2006, 107, 2373–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Pool | Epitope Identification | Class I Allele(s) with High Affinity to Epitope |
|---|---|---|
| A | 15 peptides | |
| B1 | RT (181–189) | HLA-A*02:01(11.5 nM) |
| RT (158–166) | HLA-A*03:01(12.8 nM), A*11:01(8.3 nM), A*:68:01(176.2 nM) | |
| RT (110–118) | HLA-A*68:02(20.2 nM) | |
| RT (163–171) | HLA-B*15:01(24.8 nM) | |
| Gag (433–440) | HLA-A*02:01(12.5 nM) | |
| B2 | RT (227–234) | HLA-A*02:01(17.7 nM) |
| RT (73–82) | HLA-A*03:01(20.3 nM), A*11:01(40.8 nM) | |
| RT (156–164) | HLA-B*07:02(9 nM), B*35:01(43.5 nM) | |
| Gag (362–370) | HLA-A*02:01(5.9 nM) | |
| Nef (134–143) | HLA-A*24:02(558.3 nM), A*29:02(144.8 nM) | |
| B3 | RT (232–241) | HLA-A*02:01(43 nM) |
| RT (240–249) | HLA-A*11:01(17.1 nM), A*68:01(75.9 nM) | |
| RT (18–26) | HLA-B*08:01(41.5 nM) | |
| RT (107–115) | HLA-A*29:02(90.2), HLA-B*35:01(21.8 nM) | |
| Gag (148–156) | HLA-B*07:02(37.3 nM) |
| Median | Q1 | Q3 | |
|---|---|---|---|
| Lymphocytes | 1685/µL | 1314/µL | 2090/µL |
| CD4+ | 548/µL | 460/µL | 742/µL |
| CD8+ | 541/µL | 331/µL | 848/µL |
| CD4+/CD8+ | 1 | 0.8 | 1.7 |
| CD4+ PD-1+ among CD4+ T cells | 23.70% | 19.70% | 29.80% |
| CD8+ PD-1+ among CD8+ T cells | 32.90% | 28.33% | 38.98% |
| Patient ID | ELISPOT Results (SFUs/10 6 PBMC) | ELISPOT Results | Peptide Presentation | HLA-Typing | |||
|---|---|---|---|---|---|---|---|
| Pool A | Pool B1 | Pool B2 | Pool B3 | ||||
| SA0107 | <30 | <30 | <30 | <30 | Negative | Yes | A*29:02,*68:01; B*58:02,*81:01 |
| SA0108 | 460 | <30 | 380 | <30 | Positive | Yes | A*02:01,*24:02, B*07:02, |
| SA0109 | 60 | <30 | 65 | <30 | Positive | Yes | A*03:01,*23:01; B*07:02,*58:01 |
| SA0110 | <30 | <30 | <30 | <30 | Negative | No | A*02:05,*26:01; B*49:01,*55:01 |
| SA0111 | <30 | <30 | <30 | <30 | Negative | Yes | A*03:01,*26:01; B*38:01,*40:01 |
| SA0116 | <30 | <30 | <30 | <30 | Negative | Yes | A*02:01,*26:01; B*27:05,*39:01 |
| SA0117 | <30 | <30 | <30 | <30 | Negative | Yes | A*24:02,*29:02; B*15:01,*51:01 |
| SA0118 | <30 | <30 | <30 | <30 | Negative | Yes | A*02:01,*68:02; B*53:01,*58:01 |
| SA0119 | <30 | <30 | <30 | <30 | Negative | Yes | A*03:01,*68:01; B*58:02,- |
| SA0120 | <30 | <30 | <30 | <30 | Negative | Yes | A*02:05,*11:01; B*35:03,*50:01 |
| A0121 | <30 | <30 | <30 | <30 | Negative | Yes | A*01:01,*26:01; B*08:01,*14:01 |
| SA0122 | 55 | <30 | <30 | <30 | Positive | Yes | A*02:01,*24:02 |
| SA0123 | <30 | <30 | <30 | <30 | Negative | Yes | A*24:02,*32:01; B*13:02,*39:01 |
| SA0124 | <30 | 41 | <30 | <30 | Positive | Yes | A*02:01,*03:01; B*07:02,*39:06 |
| SA0125 | ND | ND | ND | ND | ND | Yes | A*03:01,*03:02; B*18:01,*49:01 |
| SA0126 | <30 | 130 | 100 | 65 | Positive | Yes | A*11:01,*24:02; B*08:01,*27:05 |
| SA0127 | < 30 | <30 | <30 | <30 | Negative | No | A*23:01,*34:02; B*15:03,*58:01 |
| SA0128 | < 30 | <30 | <30 | <30 | Negative | Yes | A*29:01,*68:02; B*49:01,*53:01 |
| SA0129 | < 30 | <30 | <30 | <30 | Negative | Yes | A*02:01; B*14:01,*15:01 |
| SA0130 | 310 | 110 | <30 | <30 | Positive | Yes | A*03:01,*24:02; B*08:01,*40:01 |
| SA0131 | 455 | 115 | <30 | <30 | Positive | Yes | A*02:01,*68:01; B*15:17,*51:01 |
| SA0132 | 30 | 440 | <30 | <30 | Positive | Yes | A*11:01,*29:02; B*40:01,*44:02 |
| SA0133 | 175 | 285 | <30 | <30 | Positive | Yes | A*11:01,-; B*35:01,*55:01 |
| SA0134 | 80 | 260 | <30 | <30 | Positive | Yes | A*01:01,*29:02; B*44:03,*57:01 |
| SA0135 | ND | ND | ND | ND | ND | Yes | A*01:01,*02:02; B*07:02,*18:01 |
| SA0136 | ND | ND | ND | ND | ND | Yes | A*03:01,*25:01; B*08:01,*35:01 |
| SA0137 | 115 | 105 | <30 | <30 | Positive | Yes | A*30:02,*68:02; B*15:17,*18:01 |
| SA0138 | 555 | 200 | <30 | <30 | Positive | Yes | A*02:01,*30:02; B*41:01,*44:02 |
| SA0139 | 215 | 170 | 50 | 30 | Positive | Yes | A*03:01,*30:01; B*07:02,*53:01 |
| SA0140 | 1025 | 475 | <30 | <30 | Positive | Yes | A*68:01,*74:03; B*51:01, |
| SA0141 | 935 | 375 | <30 | <30 | Positive | Yes | A*02:01,*30:01; B*35:01,*51:01 |
| SA0142 | 295 | 110 | <30 | <30 | Positive | Yes | A*03:01,*24:02; B*07:02,*56:01 |
| SA0143 | 800 | 275 | <30 | <30 | Positive | Yes | A*24:02,*29:02; B*07:02,*13:02 |
| SA0144 | 400 | 365 | <30 | <30 | Positive | Yes | A*01:01,*30:02; B*08:01,*18:01 |
| SA0145 | 810 | 640 | <30 | <30 | Positive | Yes | A*02:01,*68:02; B*35:03,*53:01 |
| SA0146 | 170 | 70 | <30 | <30 | Positive | Yes | A*03:01,*30:02; B*07:02,*15:10 |
| SA0147 | ND | ND | ND | ND | ND | Yes | A*68:02,*74:01; B*44:03,*53:01 |
| SA0148 | 1160 | 510 | <30 | <30 | Positive | Yes | A*24:02,*68:02; B*35:02,*50:01 |
| SA0149 | 365 | 110 | <30 | <30 | Positive | Yes | A*02:01,*74:01; B*15:10,*53:01 |
| SA0150 | 1695 | 1655 | 30 | 90 | Positive | Yes | A*03:01,*30:01; B*13:02,*27:05 |
| SA0151 | 550 | 445 | <30 | <30 | Positive | Yes | A*01:01,*03:01; B*07:02,*08:01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fleury, H.; Caldato, S.; Recordon-Pinson, P.; Thebault, P.; Guidicelli, G.-L.; Hessamfar, M.; Morlat, P.; Bonnet, F.; Visentin, J. ART-Treated Patients Exhibit an Adaptive Immune Response against the HFVAC Peptides, a Potential HIV-1 Therapeutic Vaccine (Provir/Latitude45 Study). Viruses 2020, 12, 1256. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111256
Fleury H, Caldato S, Recordon-Pinson P, Thebault P, Guidicelli G-L, Hessamfar M, Morlat P, Bonnet F, Visentin J. ART-Treated Patients Exhibit an Adaptive Immune Response against the HFVAC Peptides, a Potential HIV-1 Therapeutic Vaccine (Provir/Latitude45 Study). Viruses. 2020; 12(11):1256. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111256
Chicago/Turabian StyleFleury, Hervé, Sabrina Caldato, Patricia Recordon-Pinson, Patricia Thebault, Gwenda-Line Guidicelli, Mojgan Hessamfar, Philippe Morlat, Fabrice Bonnet, and Jonathan Visentin. 2020. "ART-Treated Patients Exhibit an Adaptive Immune Response against the HFVAC Peptides, a Potential HIV-1 Therapeutic Vaccine (Provir/Latitude45 Study)" Viruses 12, no. 11: 1256. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111256