JSRV Intragenic Enhancer Element Increases Expression from a Heterologous Promoter and Promotes High Level AAV-Mediated Transgene Expression in the Lung and Liver of Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
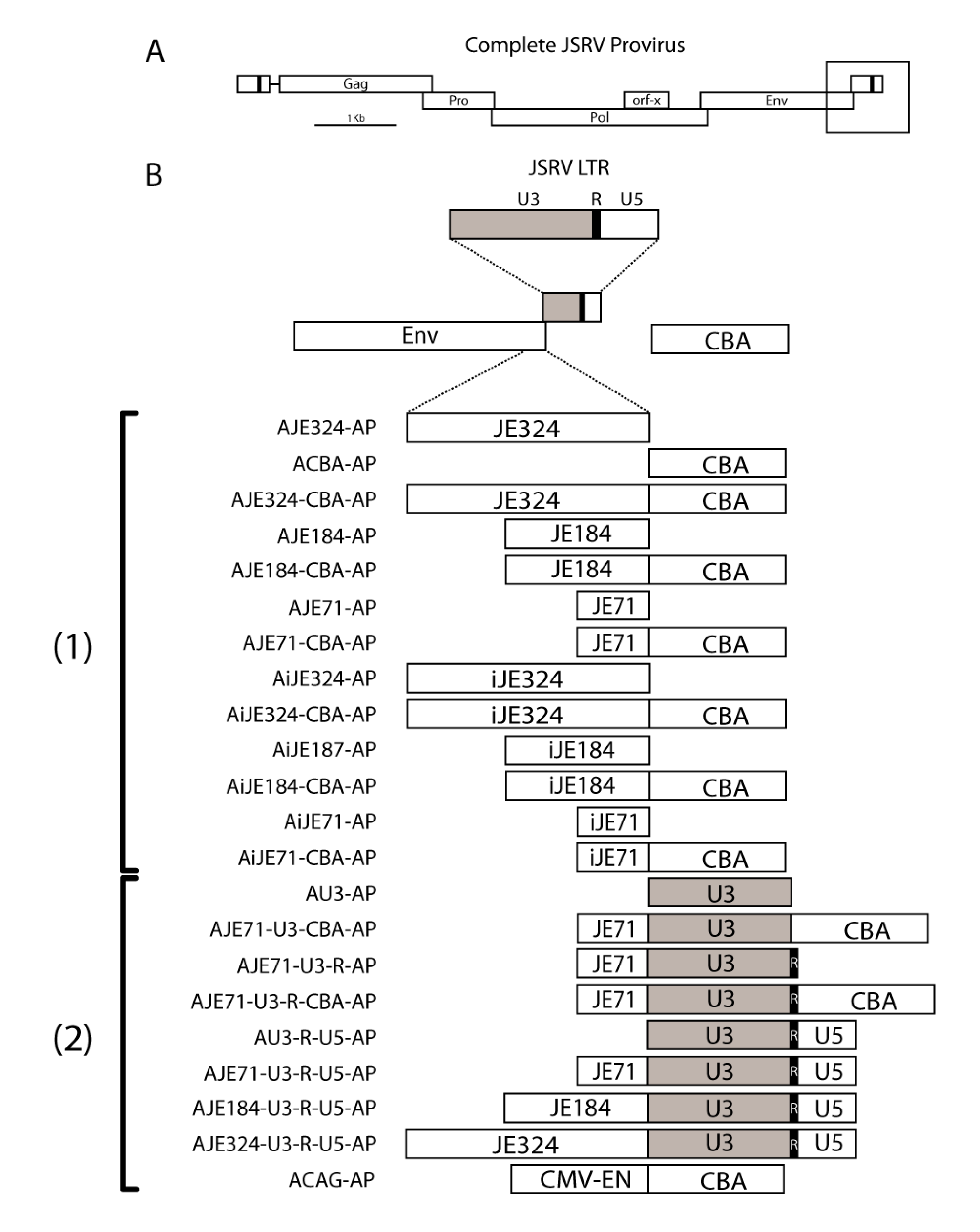
2.2. Molecular Cloning
2.3. Transfection of Mammalian Cells
2.4. Preparation of Cell Lysates
2.5. Beta-Galactosidase Assay
2.6. In Vivo Administration of AAV Vectors
2.7. Determination of Alkaline Phosphatase Activity
3. Results
3.1. JE on Its Own Functions Poorly as a Promoter
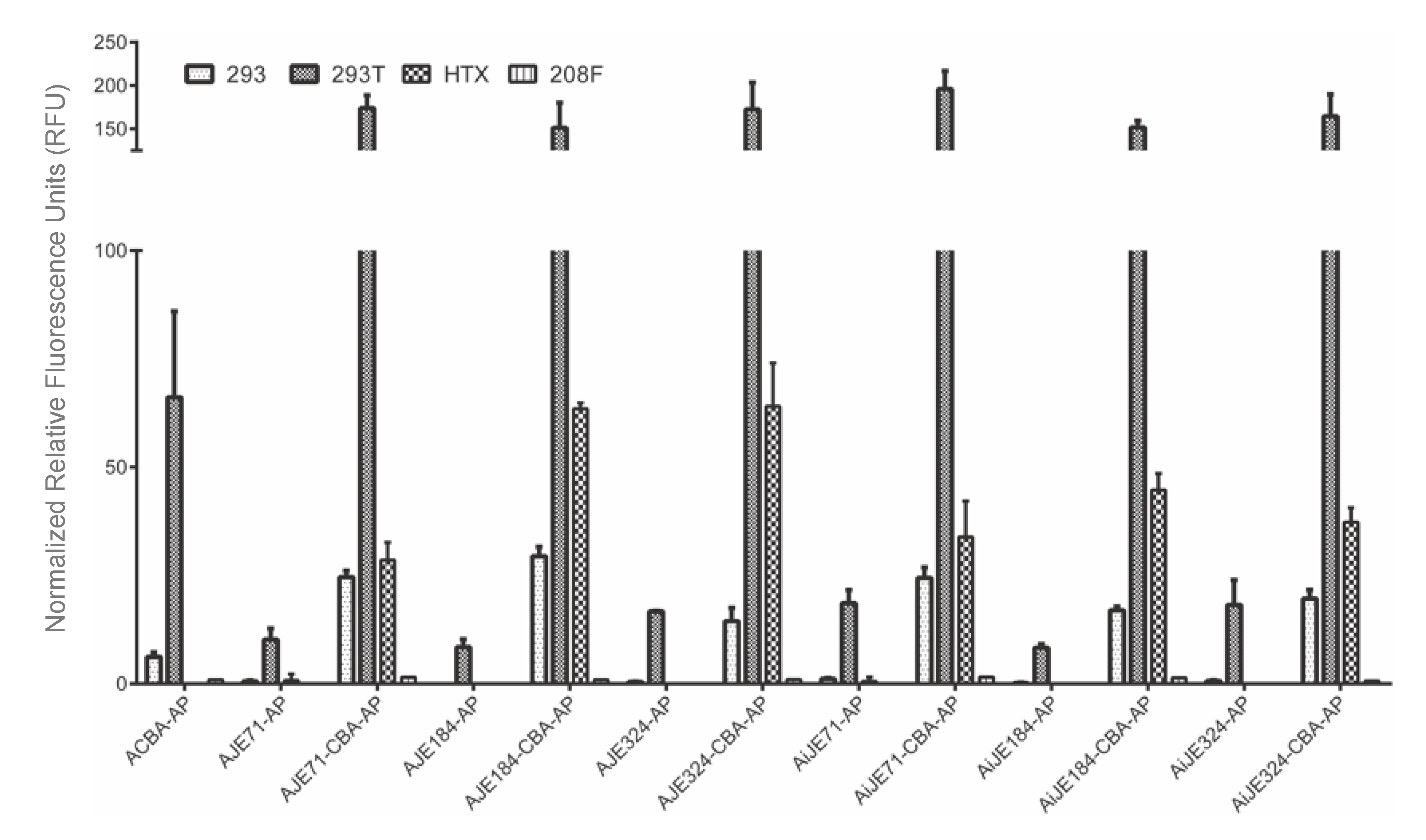
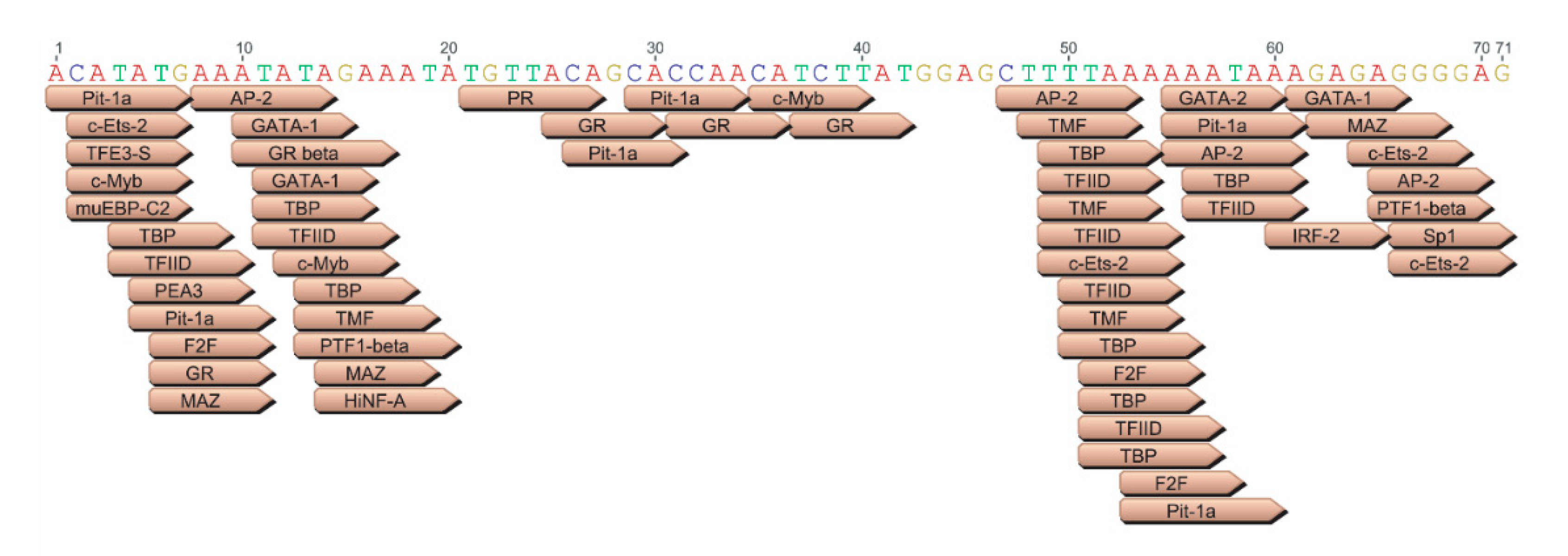
3.2. JE Enhances Expression from a Heterologous Promoter, in Both Forward and Reverse Orientations, and Extending JE from the 5’ End Does Not Appear to Further Increase Expression
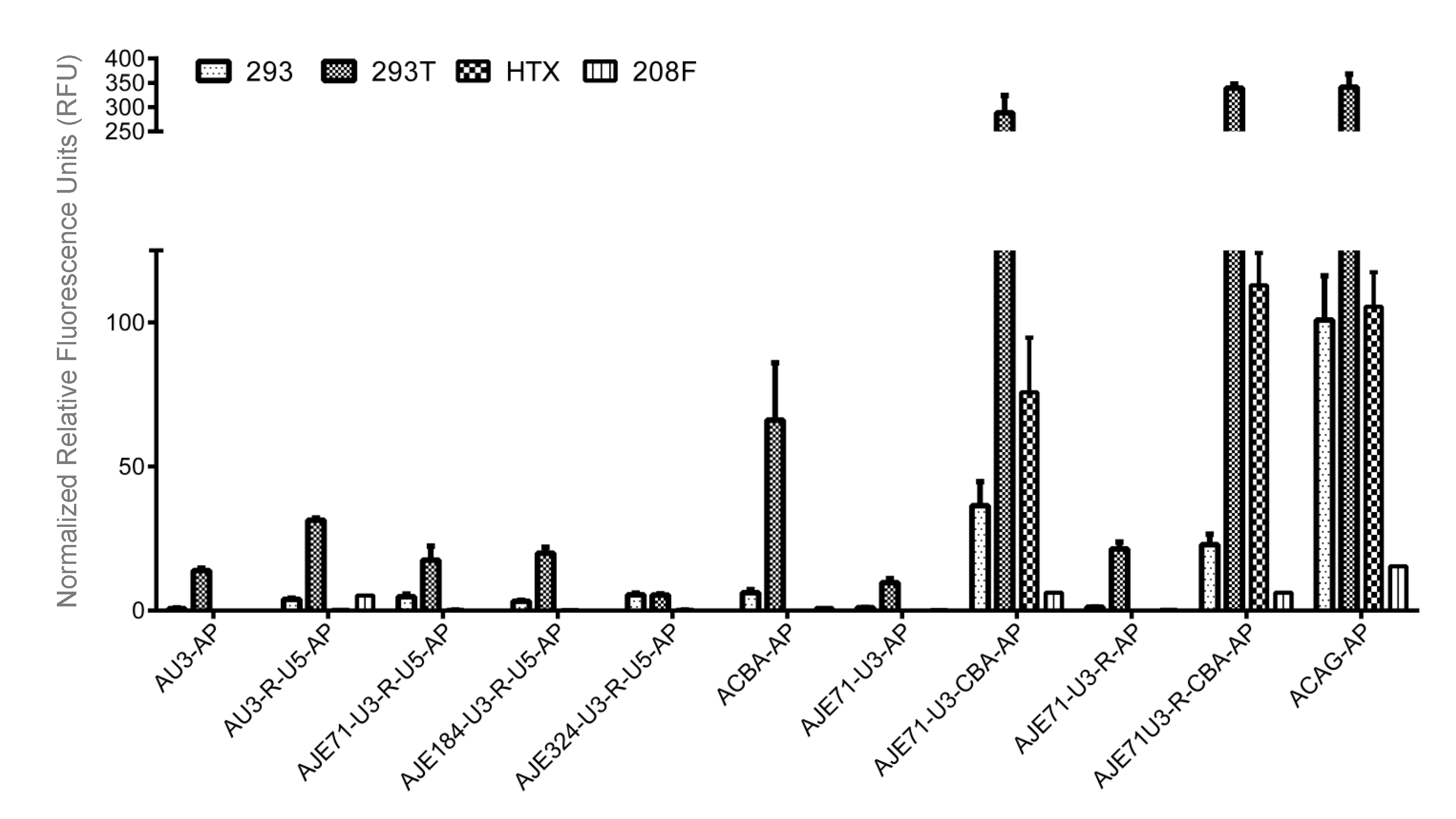
3.3. A Hybrid Promoter Consisting of JE, the U3 and R Regions of the LTR, and the Chicken Beta Actin Promoter Is Highly Active in a Variety of Cell Lines
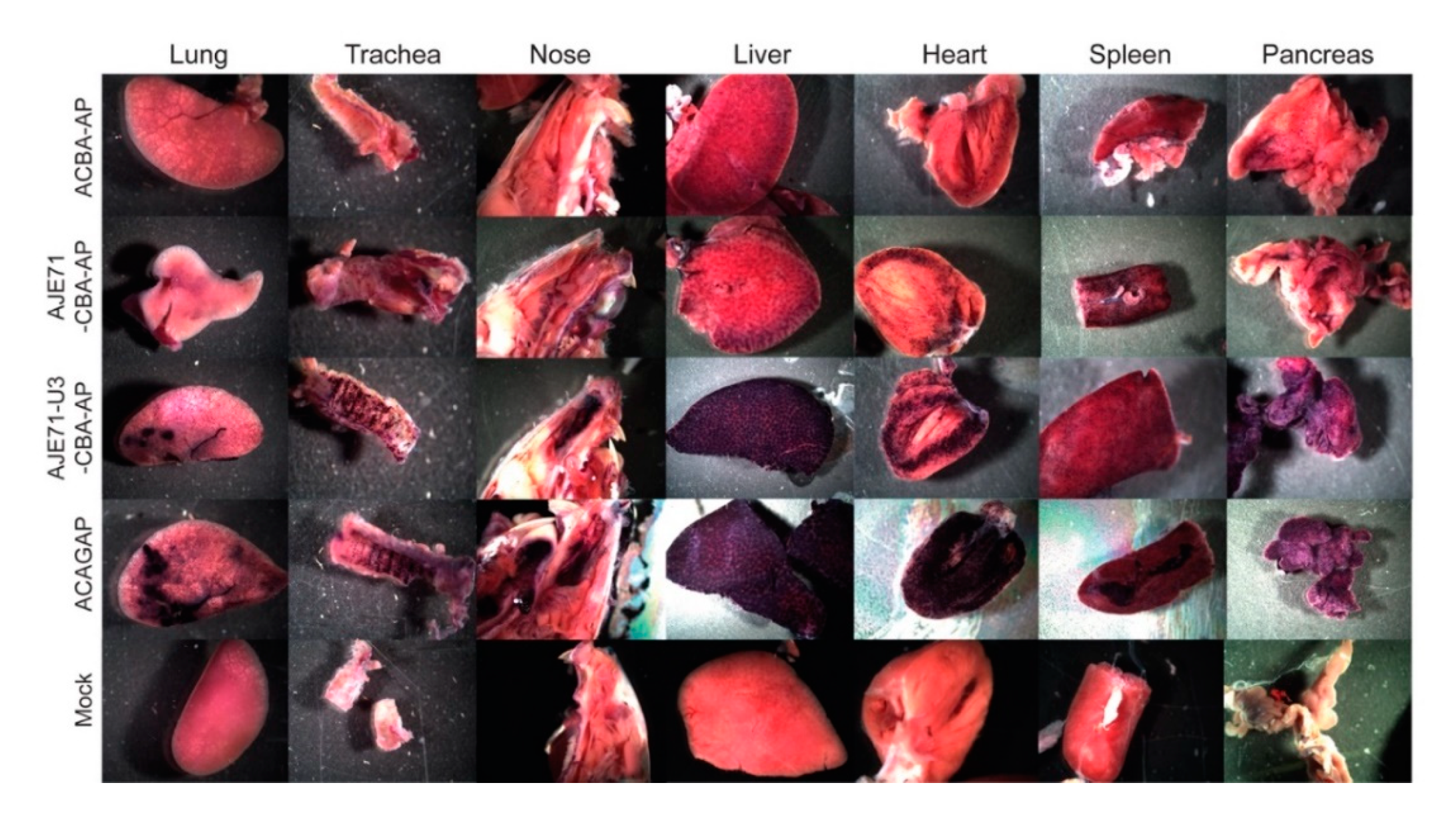
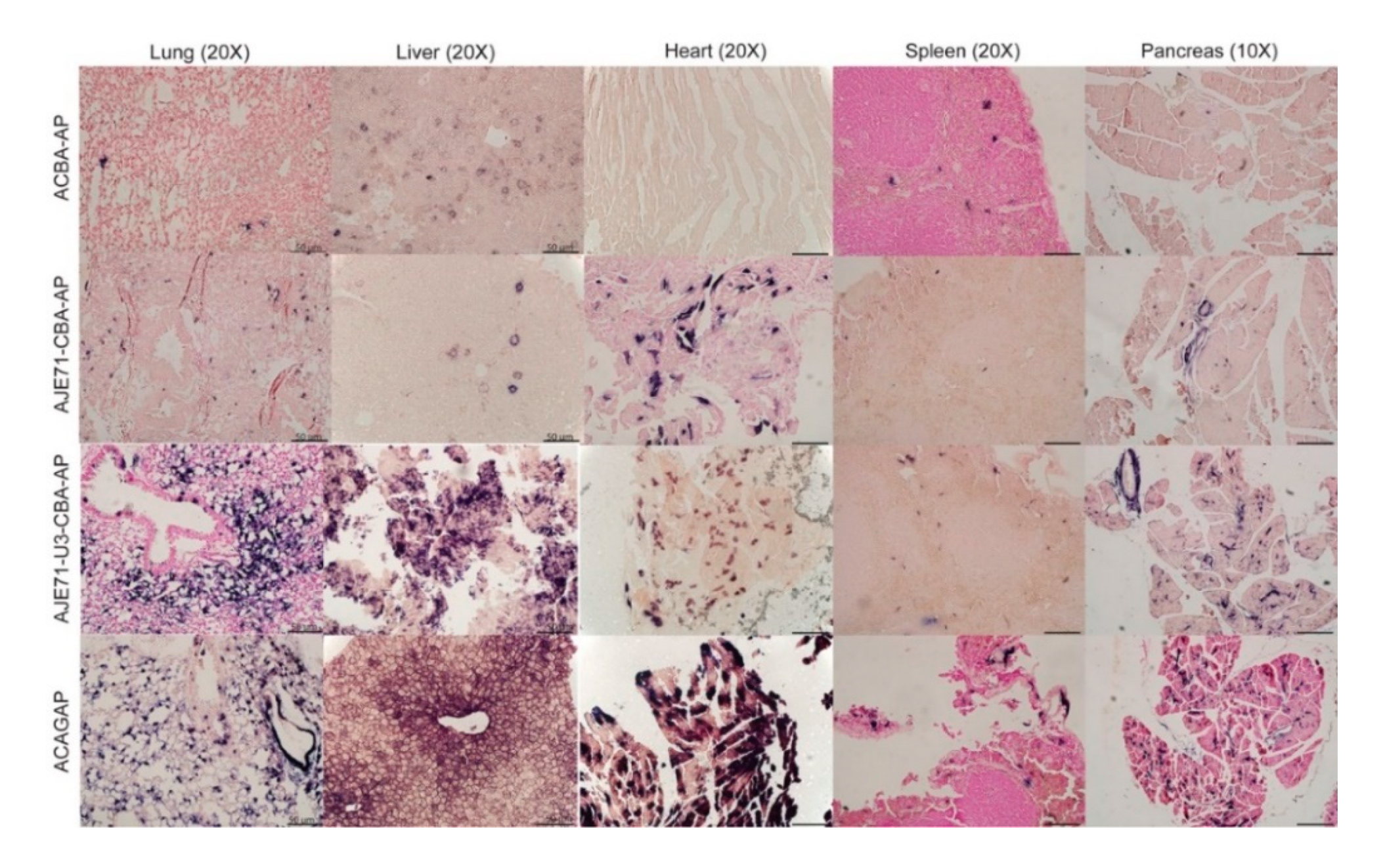
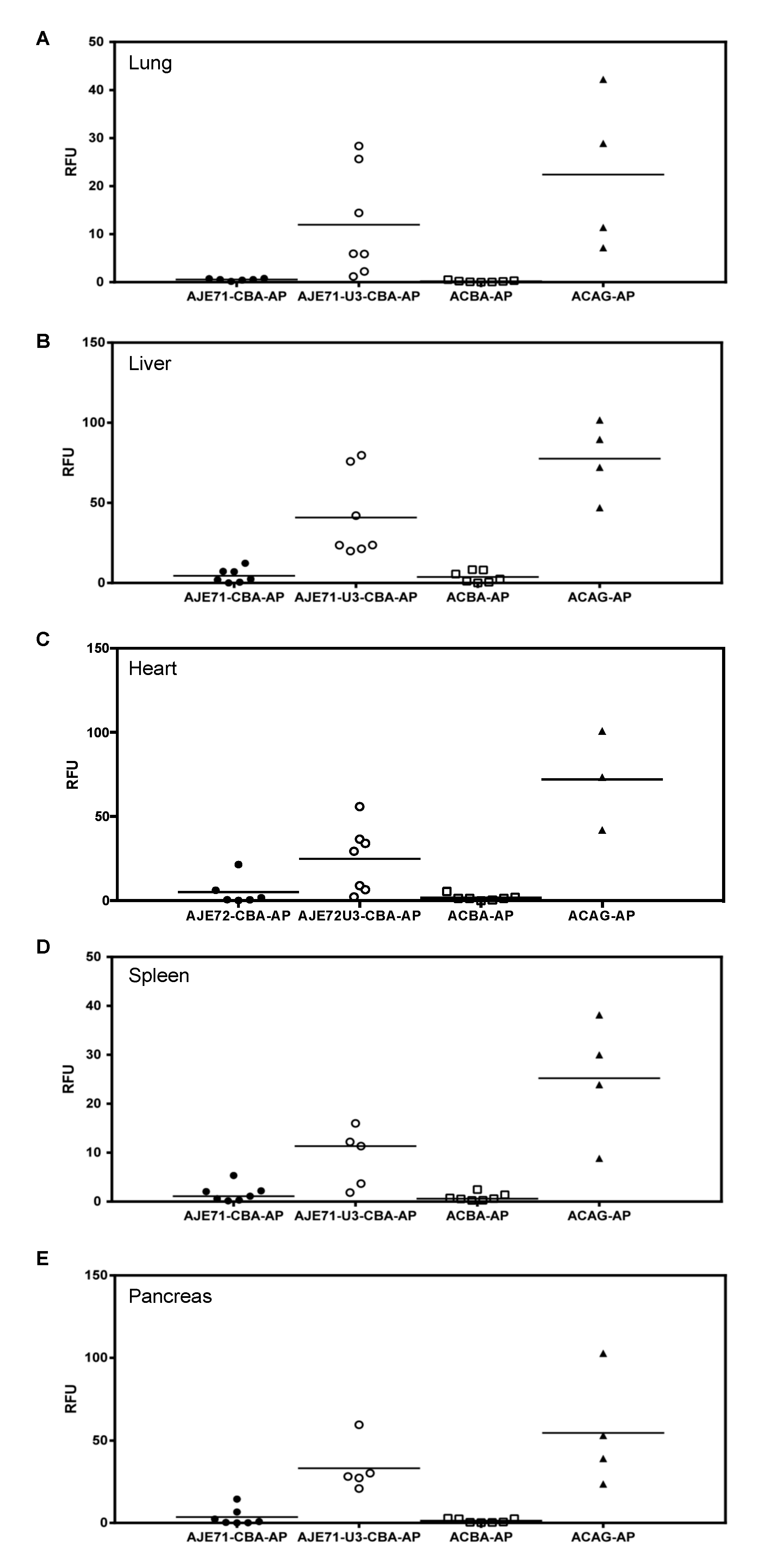
3.4. The JE/JSRV LTR/CBA Hybrid Promoter Is Active in the Lungs, Nose, Trachea, Liver, Heart, Spleen, and Pancreas of Mice
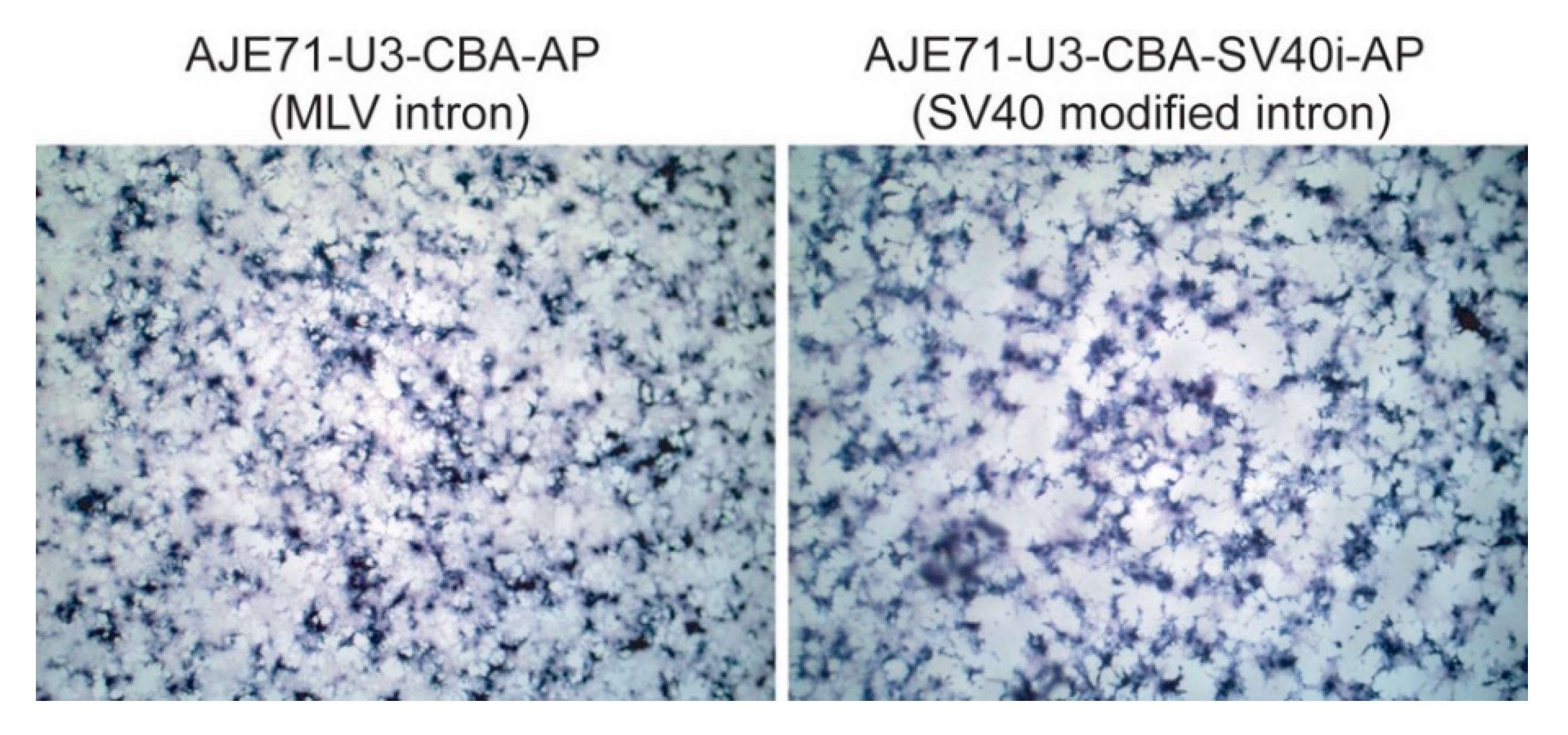
3.5. A Shorter Intron Can Be Used in Conjunction with AJE71-U3-CBA-AP without Reducing Transgene Expression
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gonçalves, M.A. Adeno-associated virus: From defective virus to effective vector. Virol. J. 2005, 2, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grieger, J.C.; Samulski, R.J. Adeno-associated virus vectorology, manufacturing, and clinical applications. Methods Enzymol. 2012, 507, 229–254. [Google Scholar] [CrossRef] [PubMed]
- Bainbridge, J.W.; Mehat, M.S.; Sundaram, V.; Robbie, S.J.; Barker, S.E.; Ripamonti, C.; Georgiadis, A.; Mowat, F.M.; Beattie, S.G.; Gardner, P.J.; et al. Long-term effect of gene therapy on Leber’s congenital amaurosis. N. Engl. J. Med. 2015, 372, 1887–1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L.; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. BioDrugs 2017, 31, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Palmarini, M.; Fan, H. Molecular biology of jaagsiekte sheep retrovirus. Curr. Top. Microbiol. Immunol. 2003, 275, 81–115. [Google Scholar]
- Rosales Gerpe, M.C.; van Lieshout, L.P.; Domm, J.M.; Ingrao, J.C.; Datu, J.; Walsh, S.R.; Yu, D.L.; Jong, J.; Krell, P.J.; Wootton, S.K. The U3 and Env Proteins of Jaagsiekte Sheep Retrovirus and Enzootic Nasal Tumor Virus Both Contribute to Tissue Tropism. Viruses 2019, 11, 1061. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.L.; Linnerth-Petrik, N.M.; Halbert, C.L.; Walsh, S.R.; Miller, A.D.; Wootton, S.K. Jaagsiekte sheep retrovirus and enzootic nasal tumor virus promoters drive gene expression in all airway epithelial cells of mice but only induce tumors in the alveolar region of the lungs. J. Virol. 2011, 85, 7535–7545. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, S.; Nelson-Rees, W.A.; Toth, E.M.; Arnstein, P.; Gardner, M.B. Characterization of a newly derived human sarcoma cell line (HT-1080). Cancer 1974, 33, 1027–1033. [Google Scholar] [CrossRef]
- Palmarini, M.; Sharp, J.M.; de las Heras, M.; Fan, H. Jaagsiekte sheep retrovirus is necessary and sufficient to induce a contagious lung cancer in sheep. J. Virol. 1999, 73, 6964–6972. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.D.; Chen, F. Retrovirus packaging cells based on 10A1 murine leukemia virus for production of vectors that use multiple receptors for cell entry. J. Virol. 1996, 70, 5564–5571. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Lab. Press: Plainview, NY, USA, 2001. [Google Scholar]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harb. Lab. Press: Plainview, NY, USA, 1972. [Google Scholar]
- van Lieshout, L.P.; Domm, J.M.; Wootton, S.K. AAV-Mediated Gene Delivery to the Lung. Methods Mol. Biol. 2019, 1950, 361–372. [Google Scholar] [CrossRef]
- Aurnhammer, C.; Haase, M.; Muether, N.; Hausl, M.; Rauschhuber, C.; Huber, I.; Nitschko, H.; Busch, U.; Sing, A.; Ehrhardt, A.; et al. Universal real-time PCR for the detection and quantification of adeno-associated virus serotype 2-derived inverted terminal repeat sequences. Hum. Gene Ther. Methods 2012, 23, 18–28. [Google Scholar] [CrossRef]
- Santry, L.A.; Ingrao, J.C.; Yu, D.L.; de Jong, J.G.; van Lieshout, L.P.; Wood, G.A.; Wootton, S.K. AAV vector distribution in the mouse respiratory tract following four different methods of administration. BMC Biotechnol. 2017, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Banerji, J.; Rusconi, S.; Schaffner, W. Expression of a beta-globin gene is enhanced by remote SV40 DNA sequences. Cell 1981, 27, 299–308. [Google Scholar] [CrossRef]
- Caporale, M.; Arnaud, F.; Mura, M.; Golder, M.; Murgia, C.; Palmarini, M. The signal peptide of a simple retrovirus envelope functions as a posttranscriptional regulator of viral gene expression. J. Virol. 2009, 83, 4591–4604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Faller, D. The long terminal repeats of a murine retrovirus encode a trans-activator for cellular genes. J. Biol. Chem. 1994, 269, 19691–19694. [Google Scholar] [PubMed]
- Choi, S.; Faller, D. A transcript from the long terminal repeats of a murine retrovirus associated with trans activation of cellular genes. J. Virol. 1995, 69, 7054–7060. [Google Scholar] [CrossRef] [Green Version]
- Forman, L.; Pal-Ghosh, R.; Spanjaard, R.; Faller, D.; Ghosh, S. Identification of LTR-specific small non-coding RNA in FeLV infected cells. FEBS Lett. 2009, 583, 1386–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinn, P.; Burnight, E.; Shen, H.; Fan, H.; McCray, P.J. Inclusion of Jaagsiekte sheep retrovirus proviral elements markedly increases lentivirus vector pseudotyping efficiency. Mol. Ther. 2005, 11, 460–469. [Google Scholar] [CrossRef]
- Luciw, P.; Bishop, J.; Varmus, H.; Capecchi, M. Location and function of retroviral and SV40 sequences that enhance biochemical transformation after microinjection of DNA. Cell 1983, 33, 705–716. [Google Scholar] [CrossRef]
- Laimins, L.; Tsichlis, P.; Khoury, G. Multiple enhancer domains in the 3’ terminus of the Prague strain of Rous sarcoma virus. Nucleic Acids Res. 1984, 12, 6427–6442. [Google Scholar] [CrossRef] [Green Version]
- Tsichlis, P.; Donehower, L.; Hager, G.; Zeller, N.; Malavarca, R.; Astrin, S.; Skalka, A. Sequence comparison in the crossover region of an oncogenic avian retrovirus recombinant and its nononcogenic parent: Genetic regions that control growth rate and oncogenic potential. Mol. Cell. Biol. 1982, 2, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Chesters, P.; Smith, L.; Nair, V. E (XSR) element contributes to the oncogenicity of Avian leukosis virus (subgroup J). J. Gen. Virol. 2006, 87, 2685–2692. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, D.L.; Chow, N.; Wootton, S.K. JSRV Intragenic Enhancer Element Increases Expression from a Heterologous Promoter and Promotes High Level AAV-Mediated Transgene Expression in the Lung and Liver of Mice. Viruses 2020, 12, 1266. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111266
Yu DL, Chow N, Wootton SK. JSRV Intragenic Enhancer Element Increases Expression from a Heterologous Promoter and Promotes High Level AAV-Mediated Transgene Expression in the Lung and Liver of Mice. Viruses. 2020; 12(11):1266. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111266
Chicago/Turabian StyleYu, Darrick L., Natalie Chow, and Sarah K. Wootton. 2020. "JSRV Intragenic Enhancer Element Increases Expression from a Heterologous Promoter and Promotes High Level AAV-Mediated Transgene Expression in the Lung and Liver of Mice" Viruses 12, no. 11: 1266. https://0-doi-org.brum.beds.ac.uk/10.3390/v12111266