Experimental Usutu Virus Infection in Domestic Canaries Serinus canaria


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Birds
2.2. USUV Challenge
2.3. Sample Collection
2.4. Histopathology and Immunohistochemistry
2.5. USUV Genome Detection
2.6. Detection of Antibodies to USUV
2.7. Statistical Analyses
3. Results
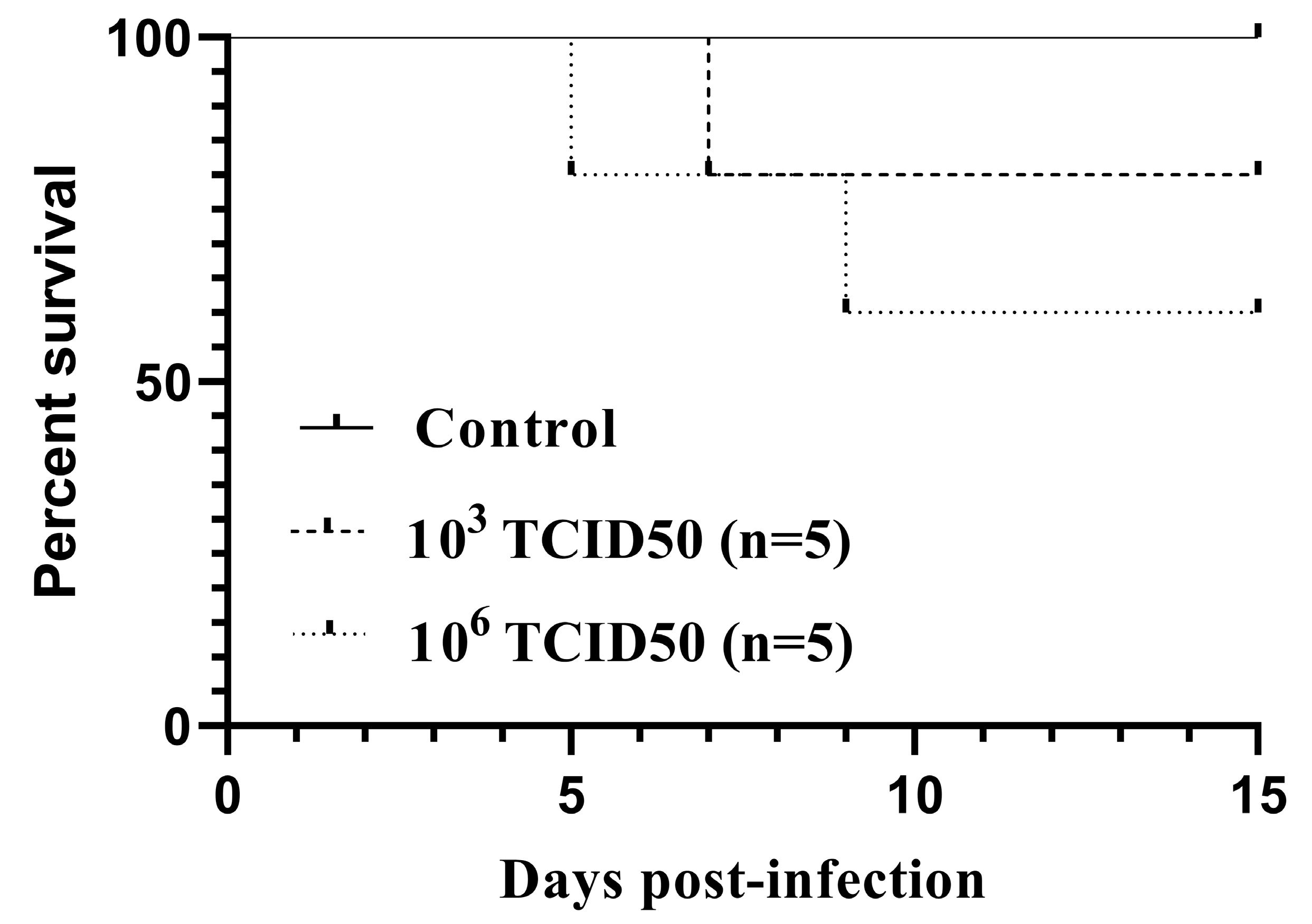
3.1. Survival and Body Weight Changes
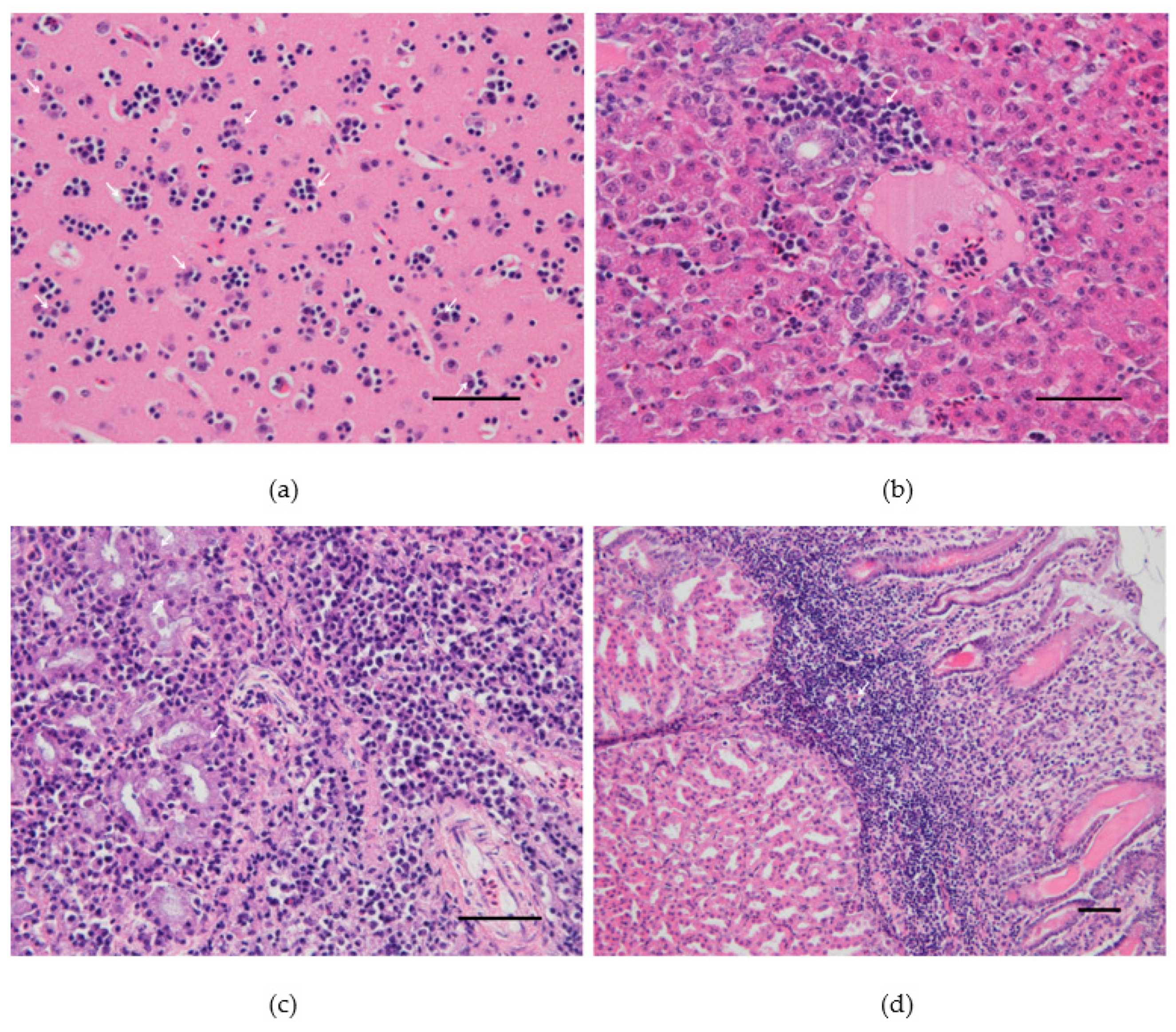
3.2. Necropsy and Histopathology Findings
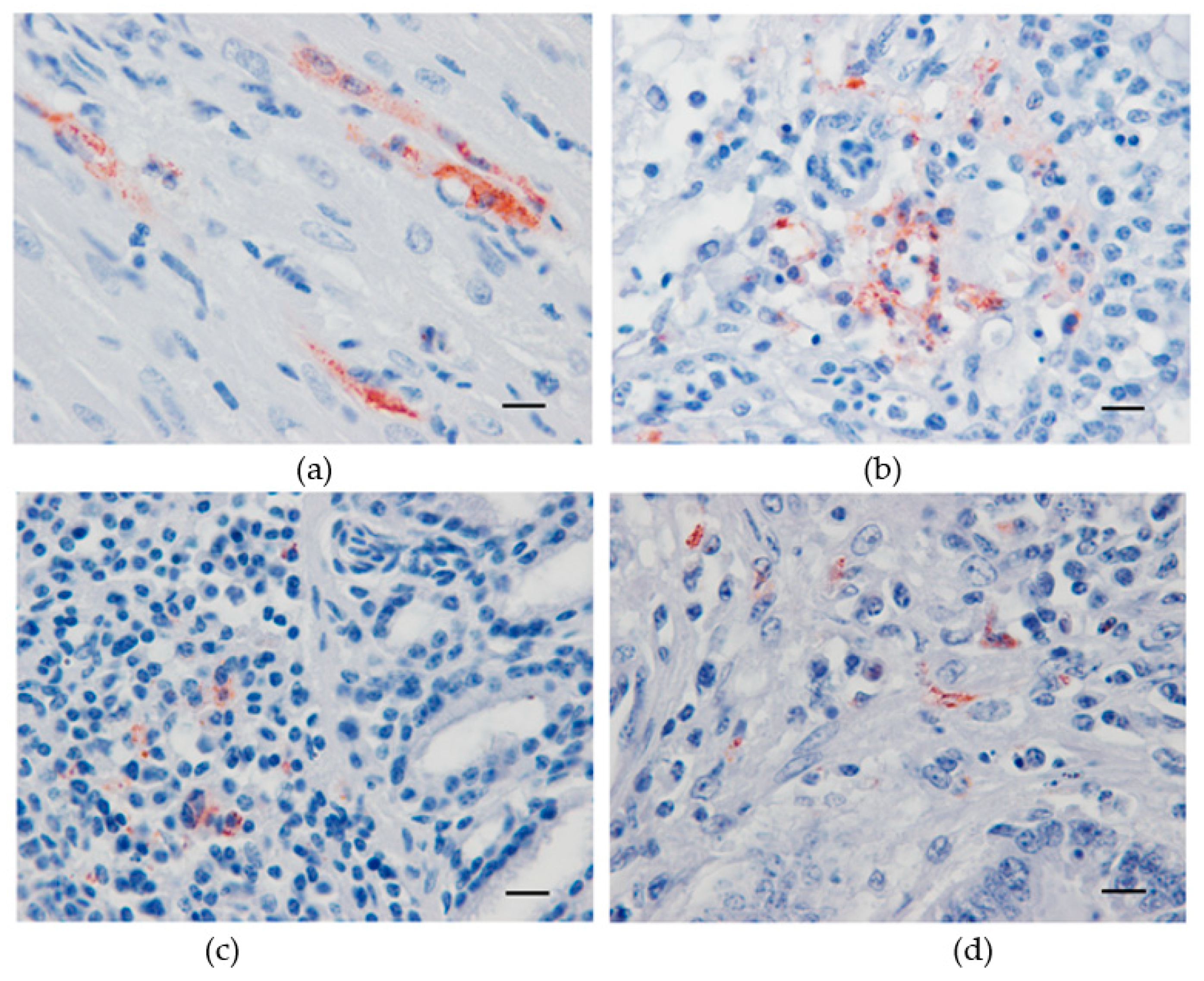
3.3. Virus Detection by Immunohistochemistry
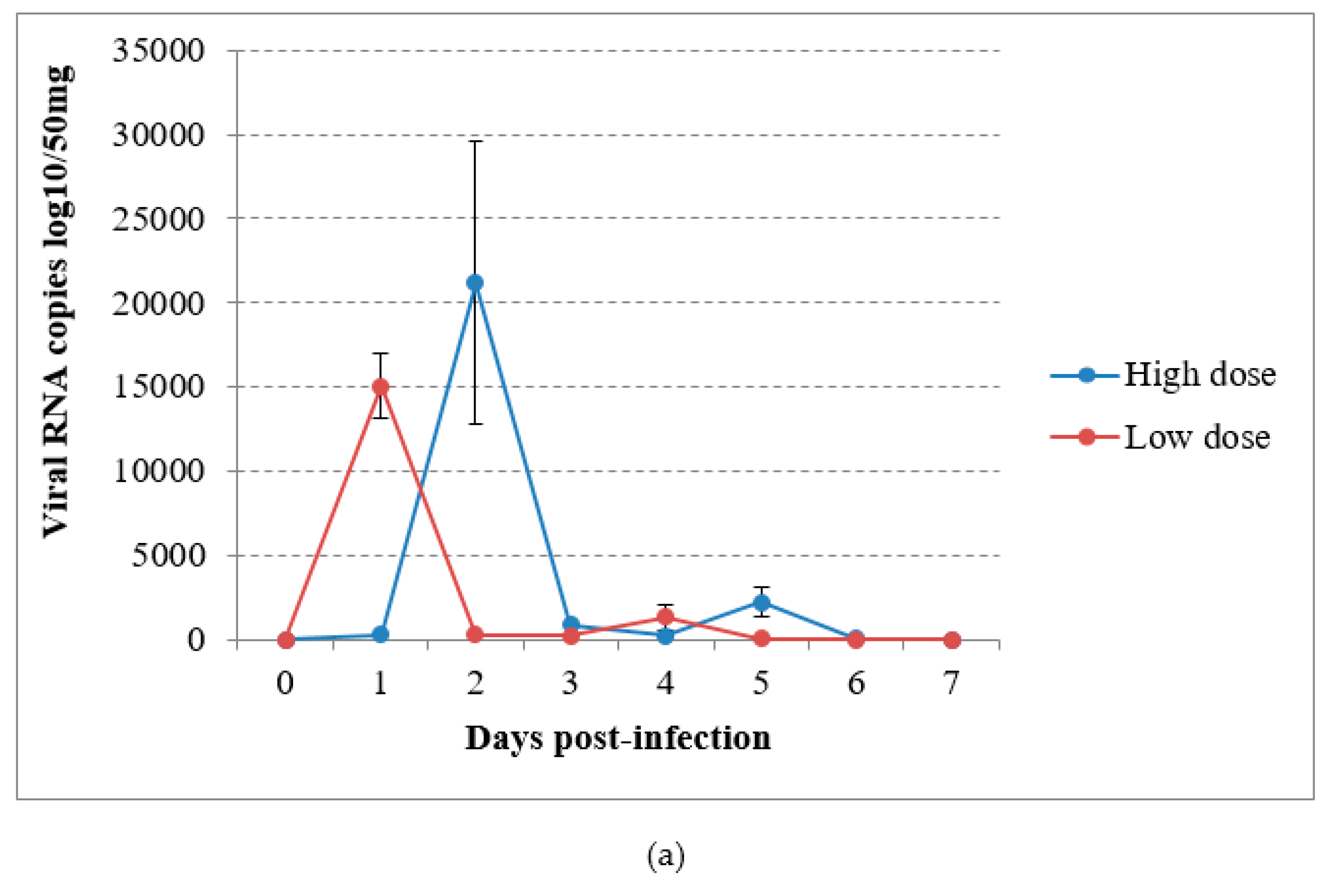
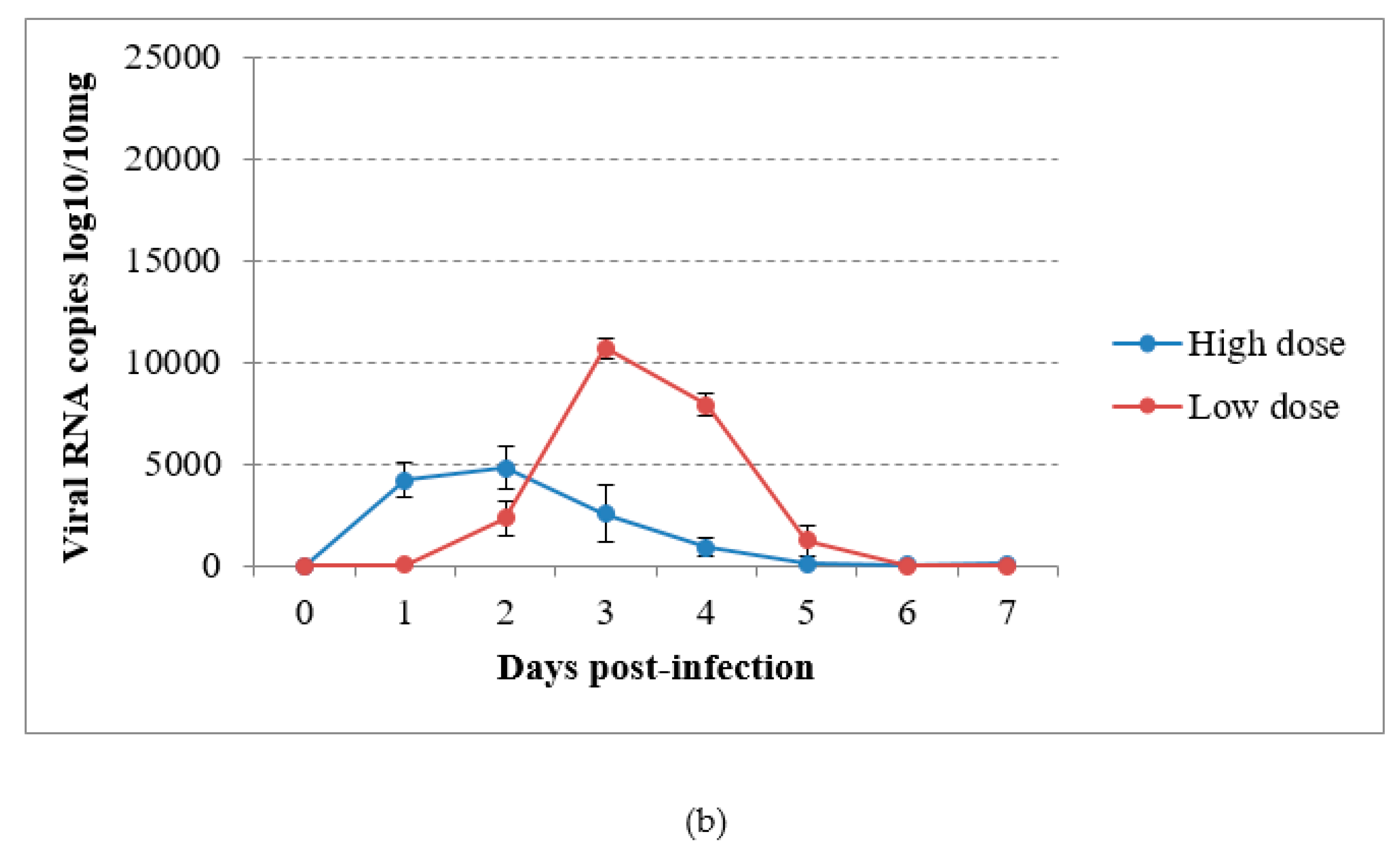
3.4. Virus Detection by RT-qPCR
3.5. Antibody Response to USUV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the Genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, A.; Mezei, E.; Nagy, O.; Bakonyi, T.; Csonka, N.; Kaposi, M.; Koroknai, A.; Szomor, K.; Rigó, Z.; Molnár, Z.; et al. Extraordinary increase in West Nile virus cases and first confirmed human Usutu virus infection in Hungary, 2018. Eurosurveillance 2019, 24, 1900038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Percivalle, E.; Sassera, D.; Rovida, F.; Isernia, P.; Fabbi, M. Usutu Virus Antibodies in Blood Donors. Vector Borne Zoonotic Dis. 2017, 17, 658–661. [Google Scholar] [CrossRef]
- Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A.; Rossini, G.; Bonilauri, P.; Gerunda, G.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; et al. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August–September 2009. Eurosurveillance 2009, 14, 19448. [Google Scholar]
- Kaic, B.; Barbic, L.; Lesnikar, V.; Listes, E.; Stevanovic, V.; Savini, G. First evidence of simultaneous occurrence of West Nile virus and Usutu virus neuroinvasive disease in humans in Croatia during the 2013 outbreak. Infection 2014, 42, 689–695. [Google Scholar]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.M.T.; Tagliazucchi, S. First human Case of Usutu Virus neuroinvasive Infection, Italy, August-September 2009. Eurosurveillance 2009, 14, 19446. [Google Scholar]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M.F.; Girard, G.; Van De Perre, P.; Salinas, S.; et al. Human Usutu Virus Infection with Atypical Neurologic Presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef] [Green Version]
- Grottola, A.; Marcacci, M.; Tagliazucchi, S.; Gennari, W.; Di Gennaro, A.; Orsini, M.; Marchegiano, P.; Marini, V.; Meacci, M.; Rumpianesi, F.; et al. Usutu virus infections in humans: a retrospective analysis in the municipality of Modena, Italy. Clin. Microbiol. Infect. 2017, 23, 33–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu Virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, E.; Linden, A.; Desmecht, D.; Garigliany, M. Mosquito-borne epornitic flaviviruses: An update and review. J. Gen. Virol. 2019, 100, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, E.; Garigliany, M.; Hauman, D.; Paternostre, J.; Linden, A.; Franssen, M.; Sarlet, M.; Cassart, D.; Desmecht, D. First evidence of fatal Usutu virus natural infections in an Anatidae, the common scoter (Melanitta nigra). Vector Borne Zoonotic Dis. 2019, 19, 777–780. [Google Scholar] [CrossRef] [PubMed]
- Benzarti, E.; Sarlet, M.; Franssen, M.; Cadar, D.; Schmidt-Chanasit, J.; Rivas, J.; Linden, A.; Desmecht, D.; Garigliany, M. Usutu Virus Epizootic in Belgium in 2017 and 2018: Evidence of Virus Endemization and Ongoing Introduction Events. Vector Borne Zoonotic Dis. 2019, 20, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.; Id, D.F.; Id, M.E.; Fast, C.; Reuschel, M.; Müller, K.; Id, M.R.; Urbaniak, S.; Brandes, F.; Schwehn, R.; et al. West Nile Virus and Usutu Virus Monitoring of Wild Birds in Germany. Int. J. Env. Res. Public Health 2018, 15, 171. [Google Scholar] [CrossRef] [Green Version]
- Weissenböck, H.; Kolodziejek, J.; Fragner, K.; Kuhn, R.; Pfeffer, M.; Nowotny, N. Usutu virus activity in Austria, 2001–2002. Microbes. Infect. 2003, 5, 1132–1136. [Google Scholar] [CrossRef]
- Chvala, S.; Kolodziejek, J.; Nowotny, N.; Weissenbo, H. Pathology and Viral Distribution in Fatal Usutu Virus Infections of Birds from the 2001 and 2002 Outbreaks in Austria. J. Comp. Path. 2004, 131, 176–185. [Google Scholar] [CrossRef]
- Becker, N.; Jost, H.; Ziegler, U.; Eiden, M.; Ho, D.; Emmerich, P.; Becker, N.; Jo, H.; Gabriel, M.; Fichet-calvet, E.; et al. Epizootic Emergence of Usutu Virus in Wild and Captive Birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csörgo, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; et al. Emergence of Usutu Virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef] [Green Version]
- Savini, G.; Monaco, F.; Terregino, C.; Di Gennaro, A.; Bano, L.; Pinoni, C.; De Nardi, R.; Bonilauri, P.; Pecorari, M.; Di Gialleonardo, L.; et al. Usutu virus in Italy: An emergence or a silent infection? Vet. Microbiol. 2011, 151, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmetz, H.W.; Bakonyi, T.; Weissenböck, H.; Hatt, J.M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland-Genomic and pathologic comparison to other central European outbreaks. Vet. Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garigliany, M.; Linden, A.; Gilliau, G.; Levy, E.; Sarlet, M.; Franssen, M.; Benzarti, E.; Derouaux, A.; Francis, F.; Desmecht, D. Usutu Virus, Belgium, 2016. Infect. Genet. Evol. 2017, 48, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenbo, H. Limited Pathogenicity of Usutu Virus for the Domestic Goose (Anser anser f. domestica) Following Experimental Inoculation. J. Vet. Med. 2006, 53, 171–175. [Google Scholar] [CrossRef]
- Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; et al. Limited pathogenicity of Usutu virus for the domestic chicken (Gallus domesticus). Avian. Pathol. 2005, 34, 392–395. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, S.; Tenorio, A. Evidence that Passerine Birds Act as Amplifying Hosts for Usutu Virus Circulation. Ecohealth 2019. Epub. [Google Scholar] [CrossRef]
- Hofmeister, E.K.; Lund, M.; Bochsler, V.S. West Nile Virus Infection in American Singer Canaries: An Experimental Model in a Highly Susceptible Avian Species. Vet. Pathol. 2018, 55, 531–538. [Google Scholar] [CrossRef]
- Ziegler, U.; Jost, H.; Muller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Klaus-Jurgen Danner, N.B.; Skuballa, J.; Hamann, H.-P.; Bosch, S.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Surasombatpattana, P.; Hamel, R.; Patramool, S.; Luplertlop, N.; Thomas, F.; Desprès, P.; Briant, L.; Yssel, H.; Missé, D. Dengue virus replication in infected human keratinocytes leads to activation of antiviral innate immune responses. Infect. Genet. Evol. 2011, 11, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- García-Bocanegra, I.; Busquets, N.; Napp, S.; Alba, A.; Zorrilla, I.; Villalba, R.; Arenas, A. Serosurvey of West Nile virus and other flaviviruses of the Japanese encephalitis antigenic complex in birds from Andalusia, southern Spain. Vector-Borne Zoonotic Dis. 2011, 11, 1107–1113. [Google Scholar] [CrossRef] [Green Version]
- Maquart, M.; Boyer, S.; Rakotoharinome, V.M.; Ravaomanana, J.; Tantely, M.L.; Heraud, J.M.; Cardinale, E. High Prevalence of West Nile Virus in Domestic Birds and Detection in 2 New Mosquito Species in Madagascar. PLoS ONE 2016, 11, e0147589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roehrig, J.T. Antigenic structure of flavivirus proteins. Adv. Virus Res. 2003, 59, 141–175. [Google Scholar] [PubMed]
- Di Gennaro, A.; Lorusso, A.; Casaccia, C.; Conte, A.; Savini, G. Serum Neutralization Assay Can Efficiently Replace Plaque Reduction Neutralization Test for Detection and Quantitation of West Nile Virus Antibodies in Human and Animal Serum Samples. Clin. Vaccine Immunol. 2014, 21, 1460–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, D.M.; Newman, C.M.; Lalli, J.; Stewart, L.M.; Koenig, M.R.; Weiler, A.M.; Semler, M.R.; Barry, G.L.; Zarbock, K.R.; Mohns, M.S.; et al. Infection via mosquito bite alters Zika virus tissue tropism and replication kinetics in rhesus macaques. Nat. Commun. 2017, 8, 2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, B.S.; Higgs, S. The enhancement of arbovirus transmission and disease by mosquito saliva is associated with modulation of the host immune response. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macneil, T.; Marin-lopez, A.; Hwang, J.; Wang, Y.; Grover, J.R. Crossm Aedes aegypti NeSt1 Protein Enhances Zika Virus Pathogenesis. J. Virol. 2019, 93, e00395-19. [Google Scholar]
- Conway, M.J.; Watson, A.M.; Colpitts, T.M.; Dragovic, S.M.; Li, Z.; Wang, P.; Feitosa, F.; Shepherd, D.T.; Ryman, K.D.; Klimstra, W.B.; et al. Mosquito Saliva Serine Protease Enhances Dissemination of Dengue Virus into the Mammalian Host. J. Virol. 2014, 88, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Welte, T.; Reagan, K.; Fang, H.; Machain-Williams, C.; Zheng, X.; Mendell, N.; Chang, G.J.J.; Wu, P.; Blair, C.D.; Wang, T. Toll-like receptor 7-induced immune response to cutaneous West Nile virus infection. J. Gen. Virol. 2009, 90, 2660–2668. [Google Scholar] [CrossRef]
- Styer, L.M.; Kent, K.A.; Albright, R.G.; Bennett, C.J.; Kramer, L.D.; Bernard, K.A. Mosquitoes inoculate high doses of West Nile virus as they probe and feed on live hosts. PLoS Pathog. 2007, 3, e132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanDalen, K.K.; Hall, J.S.; Clark, L.; McLean, R.G.; Smeraski, C. West Nile Virus Infection in American Robins: New Insights on Dose Response. PLoS ONE 2013, 8, e68537. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Avian Host and Mosquito (Diptera: Culicidae) Vector Competence Determine the Efficiency of West Nile and St. Louis Encephalitis Virus Transmission. J. Med. Entomol. 2006, 42, 367–375. [Google Scholar] [CrossRef]
- Llorente, F.; Pérez-ramírez, E.; Fernández-pinero, J.; Elizalde, M.; Figuerola, J.; Soriguer, R.C.; Jiménez-clavero, M.Á. Bagaza virus is pathogenic and transmitted by direct contact in experimentally infected partridges, but is not infectious in house sparrows and adult mice. Vet. Res. 2015, 46, 93. [Google Scholar] [CrossRef] [Green Version]
- Höfle, U.; Gamino, V.; de Mera, I.G.F.; Mangold, A.J.; Ortíz, J.-A.; de la Fuente, J. Usutu Virus in Migratory Song Thrushes, Spain. Emerg. Infect. Dis. 2013, 19, 1173–1175. [Google Scholar]
- Banet-Noach, C.; Simanov, L.; Malkinson, M. Direct (non-vector) transmission of West Nile virus in geese. Avian. Pathol. 2003, 32, 489–494. [Google Scholar] [CrossRef]
- Langevin, S.A.; Bunning, M.; Davis, B.; Komar, N. Experimental Infection of Chickens as Candidate Sentinels for West Nile Virus. Emerg. Infect. Dis. 2001, 7, 726–729. [Google Scholar] [CrossRef]
- Komar, N.; Langevin, S.; Hinten, S.; Nemeth, N.; Edwards, E.; Hettler, D.; Davis, B.; Bowen, R.; Bunning, M. Experimental Infection of North American Birds with the New York 1999 Strain of West Nile Virus. Emerg. Infect. Dis. 2003, 9, 311–322. [Google Scholar] [CrossRef]
- Nemeth, N.; Gould, D.; Bowen, R.; Komar, N. Natural and experimental West Nile virus Infection in five Raptor Species. J. Wildl. Dis. 2006, 42, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, N.M.; Hahn, D.C.; Gould, D.H.; Bowen, R.A. Experimental West Nile Virus Infection in Eastern Screech Owls (Megascops asio). Avian. Dis. 2006, 50, 252–258. [Google Scholar] [CrossRef]
- Bertran, K.; Costa, T.P.; Rivas, R.; Solanes, D.; Bensaid, A.; Page, N. Experimental West Nile Virus Infection in Gyr-Saker Hybrid Falcons. Vector Borne Zoonotic Dis. 2012, 12, 482–489. [Google Scholar]
- Gómez, C.C.; Llorente, F.; Ramírez, E.P.; Soriguer, R.C.; Sarasa, M.; Ángel, M.; Clavero, J. Experimental infection of grey partridges with Bagaza virus: Pathogenicity evaluation and potential role as a competent host. Vet. Res. 2018, 49, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, I.; Natour-altory, A.; Raibstein, I.; Kin, E.; Dahan, Y.; Krispin, H. Monitoring the uptake of live avian vaccines by their detection in feathers. Vaccine 2018, 36, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Thomsen, B.V.; Spraker, T.R.; Benson, J.M.; Oesterle, P.T.; Bright, J.M.; Muth, J.P. Clinical and Pathologic Responses of American Crows (Corvus brachyrhynchos) and Fish Crows (C. ossifragus) to Experimental West Nile Virus Infection. Vet. Pathol. 2011, 48, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirafuji, H.; Kanehira, K.; Kubo, M.; Shibahara, T.; Kamio, T. Experimental West Nile Virus Infection in Jungle Crows (Corvus macrorhynchos). Am. J. Trop. Med. Hyg 2008, 78, 838–842. [Google Scholar] [CrossRef] [Green Version]
- Pauli, A.M.; Cruz-martinez, L.A.; Ponder, J.B.; Redig, P.T.; Glaser, A.L.; Klauss, G.; Schoster, J.V. Ophthalmologic and oculopathologic finding in red-tailed hawks and Cooper’s hawks with naturally acquired West Nile virus infection. J. Am. Vet. Med. Assoc. 2007, 231, 1240–1248. [Google Scholar] [CrossRef]
- Gamino, V.; Escribano-Romero, E.; Gutierrez-Guzman, A.V.; Blazquez, A.B.; Saiz, J.C.; Hofle, U. Oculopathologic Findings in Flavivirus-Infected Gallinaceous Birds. Vet. Pathol. 2014, 51, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Klećkowska-Nawrot, J.; Goździewska-Harłajczuk, K.; Kowalczyk, A.; Łukaszewicz, E.; Nowaczyk, R. Histological, histochemical and ultrastructural studies on Harderian and lacrimal glands of the Capercaillie (Tetrao urogallus major L.). Acta. Biol. Hung. 2016, 67, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Gamino, V.; Höfle, U. Pathology and Tissue Tropism of natural West Nile Virus Infection in Birds: A Review. Vet. Res. 2013, 44, 39. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Liang, Y.; Wang, N.; Cui, L.; Chen, Z.; Wu, H.; Zhu, C.; Wang, Z.; Liu, S.; Li, H. Avian Flavivirus Infection of Monocytes Macrophages by Extensive Subversion of Host Antiviral Innate Immune Responses. J. Virol. 2019, 93, e00978-19. [Google Scholar] [CrossRef]
- Pérez-Ramírez, E.; Llorente, F.; Jiménez-Clavero, M.Á. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blázquez, A.; Escribano-romero, E.; Martín-acebes, M.A.; Petrovic, T.; Saiz, J. Limited susceptibility of mice to Usutu virus (USUV) infection and induction of flavivirus cross-protective immunity. Virol 2015, 482, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Reisen, W.K. Previous infection with West Nile or St. Louis encephalitis viruses provides cross protection during reinfection in house finches. Am. J. Trop. Med. Hyg. 2006, 75, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, N.M.; Bosco-Lauth, A.M.; Bowen, R.A. Cross-Protection Between West Nile and Japanese Encephalitis Viruses in Red-Winged Blackbirds (Agelaius phoeniceus). Avian Dis. Dig. 2009, 53, 421–425. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days Post-Infection. | |||||||
|---|---|---|---|---|---|---|---|
| Canary | 1 | 3 | 5 | 7 | 9 | 15 | |
| Group A (106TCID50) | 1 | 5.36 ± 0.01 | 5.21 ± 0.11 | 4.73 ± 0.1 | 0.37 ± 0.01 | ||
| 2 | 7.01 ± 0.02 | 6.34 ± 0.03 | 7.01 ± 0.3 | ||||
| 3 | 5.99 ± 0.06 | 5.23 ± 0.04 | Insuff. | 3.71 ± 0.24 | |||
| 4 | 6.34 ± 0.03 | 6.73 ± 0.01 | 7.95 ± 0.08 | ||||
| 5 | 5.50 ± 0.12 | 5.69 ± 0.01 | 3.83 ± 0.02 | 1.64 ± 0.21 | |||
| Group B (103TCID50) | 1 | 7.13 ± 0.03 | 6.34 ± 0.03 | 2.81 ± 0.25 | 1.76 ± 0.23 | ||
| 2 | 4.38 ± 0.01 | 5.80 ± 0.01 | Insuff. | 2.09 ± 0.29 | |||
| 3 | 6.01 ± 0.02 | 5.83 ± 0.04 | 2.37 ± 0.4 | 1.6 ± 1.38 | |||
| 4 | 7.13 ± 0.08 | 7.33 ± 0.01 | 8.20 ± 0.21 | ||||
| 5 | 4.38 ± 0.03 | 5.96 ± 0.07 | 1.95 ± 0.52 | 2.00 ± 0.24 |
 Dead canary; Insuff. = Insufficient volume.
Dead canary; Insuff. = Insufficient volume.| Samples | Blood | Brain | Liver | Eye | Feathers | Lung | Kidney | Intestine | |
|---|---|---|---|---|---|---|---|---|---|
| Bird | |||||||||
| Group A | |||||||||
| Canary 4 | 5.39 ± 0.31 | 7.15 ± 0.09 | 9.05 ± 0.19 | 7.18 ± 0.15 | 4.32 ± 0.09 | 9.12 ± 0.21 | 6.22 ± 0.14 | 6.62 ± 0.02 | |
| Canary 2 | 4.60 ± 0.81 | 6.19 ± 0.03 | 7.40 ± 0.50 | 3.73 ± 0.02 | 3.05 ± 0.62 | 7.26 ± 0.1 | 7.48 ± 0.03 | 3.86 ± 0.04 | |
| Group B | |||||||||
| Canary 4 | 7.30 ± 0.41 | 7.77 ± 0.01 | 5.05 ± 0.41 | 3.91 ± 0.3 | 3.32 ± 0.11 | 3.41 ± 0.07 | 4.53 ± 0.13 | 7.54 ± 0.10 | |
| Surviving Birds | ELISA | Neutralizing Antibodies | |
|---|---|---|---|
| Group A (106TCID50) | 1 | + | 1:80 |
| 2 | + | 1:20 | |
| 3 | + | 1:40 | |
| Group B (103TCID50) | 1 | + | 1:20 |
| 2 | D | <1:5 | |
| 3 | + | 1:40 | |
| 4 | + | 1:20 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Desmecht, D.; Schmidt-Chanasit, J.; Savini, G.; Lorusso, A.; Van Laere, A.-S.; Garigliany, M.-M. Experimental Usutu Virus Infection in Domestic Canaries Serinus canaria. Viruses 2020, 12, 164. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020164
Benzarti E, Rivas J, Sarlet M, Franssen M, Desmecht D, Schmidt-Chanasit J, Savini G, Lorusso A, Van Laere A-S, Garigliany M-M. Experimental Usutu Virus Infection in Domestic Canaries Serinus canaria. Viruses. 2020; 12(2):164. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020164
Chicago/Turabian StyleBenzarti, Emna, José Rivas, Michaël Sarlet, Mathieu Franssen, Daniel Desmecht, Jonas Schmidt-Chanasit, Giovanni Savini, Alessio Lorusso, Anne-Sophie Van Laere, and Mutien-Marie Garigliany. 2020. "Experimental Usutu Virus Infection in Domestic Canaries Serinus canaria" Viruses 12, no. 2: 164. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020164