Characterization of Experimental Oro-Nasal Inoculation of Seba’s Short-Tailed Bats (Carollia perspicillata) with Bat Influenza A Virus H18N11

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus
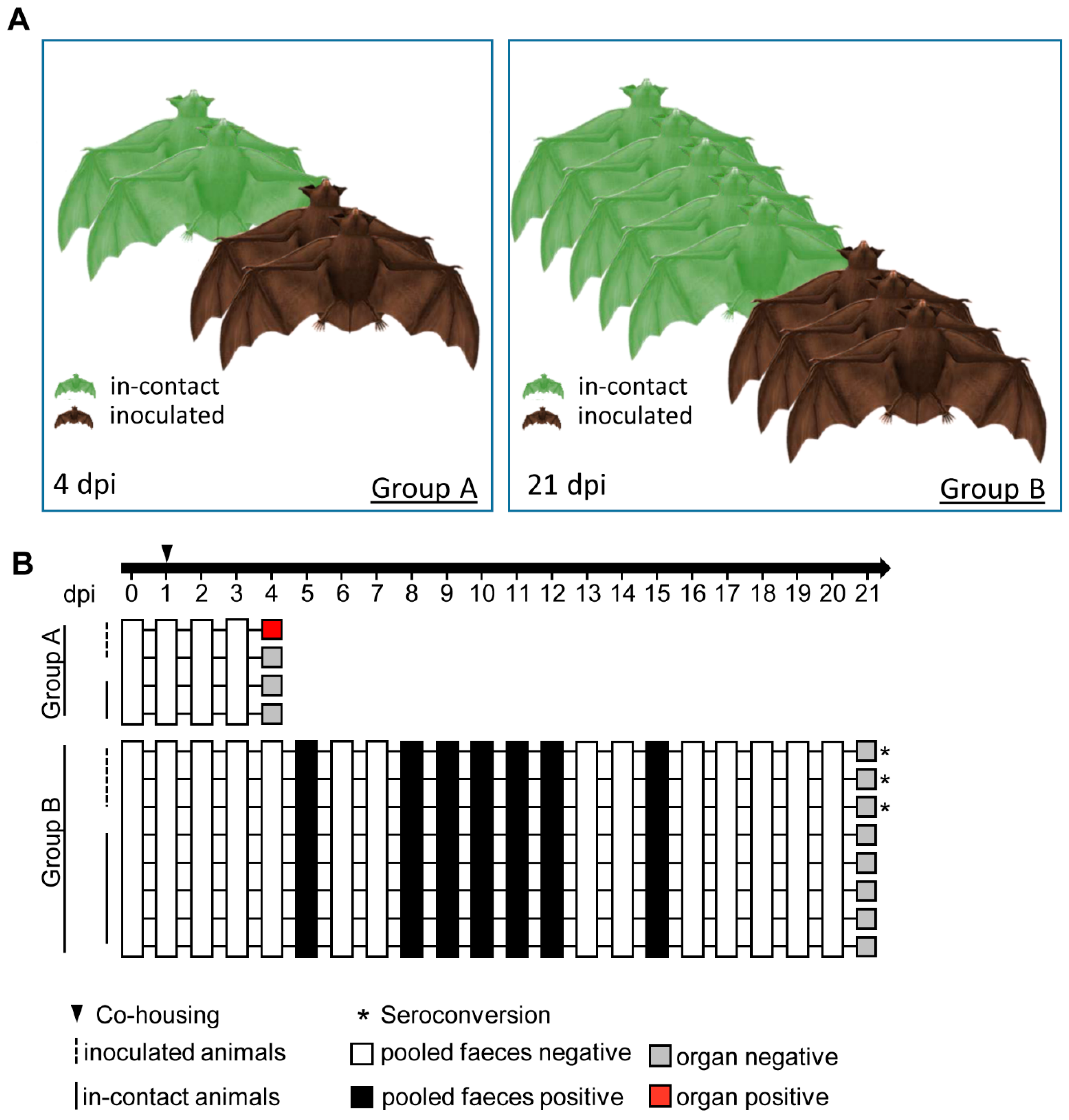
2.3. Bat Experiment and Sampling
2.4. Organ Homogenization
2.5. RNA Isolation
2.6. RT-qPCR
2.7. Virus Isolation
2.8. Serology
2.9. Necropsy and Histologic Examination
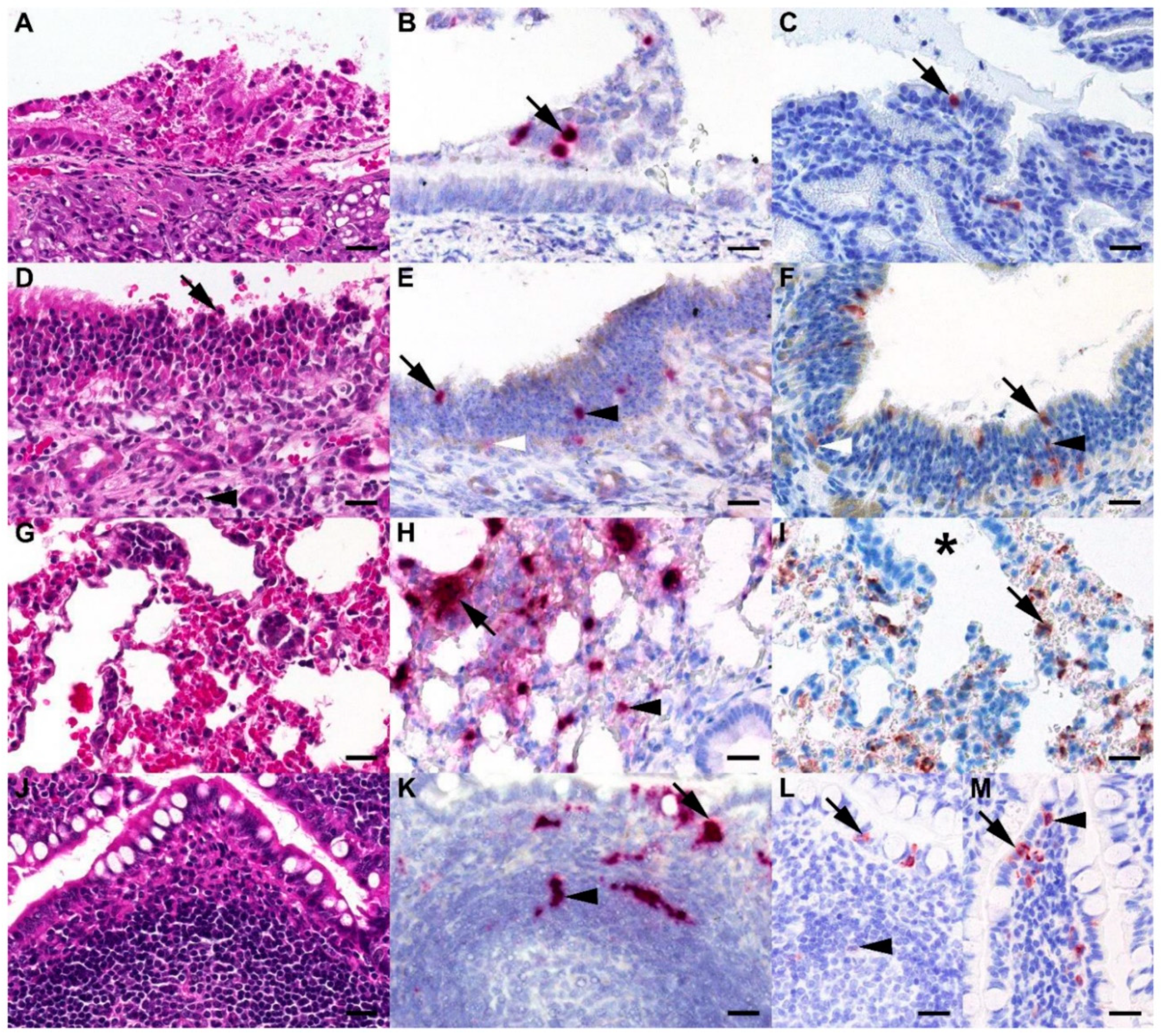
2.10. Immunohistochemistry
2.11. In Situ Hybridization
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, W.M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and Genetic Characteristics of Swine-Origin 2009 A(H1N1) Influenza Viruses Circulating in Humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Imai, M.; Watanabe, T.; Hatta, M.; Das, S.C.; Ozawa, M.; Shinya, K.; Zhong, G.; Hanson, A.; Katsura, H.; Watanabe, S.; et al. Experimental adaptation of an influenza H5 HA confers respiratory droplet transmission to a reassortant H5 HA/H1N1 virus in ferrets. Nature 2012, 486, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Desselberger, U.; Nakajima, K.; Alfino, P.; Pedersen, F.S.; Haseltine, W.A.; Hannoun, C.; Palese, P. Biochemical evidence that “new” influenza virus strains in nature may arise by recombination (reassortment). Proc. Natl. Acad. Sci. USA 1978, 75, 3341–3345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herfst, S.; Schrauwen, E.J.A.; Linster, M.; Chutinimitkul, S.; De Wit, E.; Munster, V.J.; Sorrell, E.M.; Bestebroer, T.M.; Burke, D.; Smith, D.J.; et al. Airborne Transmission of Influenza A/H5N1 Virus Between Ferrets. Science 2012, 336, 1534–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important Reservoir Hosts of Emerging Viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Wynne, J.W.; Wang, L.F. Bats and viruses: Friend or foe? PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.A.; Chen, L.-M.; Recuenco-Cabrera, S.; Ellison, J.A.; Davis, C.T.; York, I.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.; Zhu, X.; Li, Y.; Shi, M.; Zhang, J.; Bourgeois, M.; Yang, H.; Chen, X.; Recuenco-Cabrera, S.; Gómez, J.; et al. New World Bats Harbor Diverse Influenza A Viruses. PLOS Pathog. 2013, 9. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Shi, Y.; Lu, X.; He, J.; Gao, G.F.; Yan, J.; Qi, J.; Gao, G.F. Bat-Derived Influenza Hemagglutinin H17 Does Not Bind Canonical Avian or Human Receptors and Most Likely Uses a Unique Entry Mechanism. Cell Rep. 2013, 3, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sun, X.; Li, Z.; Liu, Y.; Vavricka, C.J.; Qi, J.; Gao, G.F. Structural and functional characterization of neuraminidase-like molecule n10 derived from bat influenza a virus. Proc. Natl. Acad. Sci. USA 2012, 109, 18897–18902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Yu, W.; McBride, R.; Li, Y.; Chen, L.-M.; Donis, R.O.; Tong, S.; Paulson, J.C.; Wilson, I.A. Hemagglutinin homologue from H17N10 bat influenza virus exhibits divergent receptor-binding and pH-dependent fusion activities. Proc. Natl. Acad. Sci. USA 2013, 110, 1458–1463. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Yang, H.; Guo, Z.; Yu, W.; Carney, P.J.; Li, Y.; Chen, L.-M.; Paulson, J.C.; Donis, R.O.; Tong, S.; et al. Crystal structures of two subtype N10 neuraminidase-like proteins from bat influenza A viruses reveal a diverged putative active site. Proc. Natl. Acad. Sci. USA 2012, 109, 18903–18908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambaryan, A.; Tuzikov, A.; Piskarev, V.; Yamnikova, S.S.; Lvov, D.K.; Robertson, J.; Bovin, N.V.; Matrosovich, M. Specification of Receptor-Binding Phenotypes of Influenza Virus Isolates from Different Hosts Using Synthetic Sialylglycopolymers: Non-Egg-Adapted Human H1 and H3 Influenza A and Influenza B Viruses Share a Common High Binding Affinity for 6?-Sialyl(N-acetyllactosamine). Virology 1997, 232, 345–350. [Google Scholar] [PubMed]
- Sauter, N.K.; Bednarski, M.D.; Wurzburg, B.A.; Hanson, J.E.; Whitesides, G.M.; Skehel, J.J.; Wiley, N.C. Hemagglutinins from two influenza virus variants bind to sialic acid derivatives with millimolar dissociation constants: A 500-MHz proton nuclear magnetic resonance study. Biochemistry 1989, 28, 8388–8396. [Google Scholar] [CrossRef] [PubMed]
- Karakus, U.; Thamamongood, T.; Ciminski, K.; Ran, W.; Günther, S.C.; Pohl, M.; Eletto, D.; Jeney, C.; Hoffmann, D.; Reiche, S.; et al. MHC class II proteins mediate cross-species entry of bat influenza viruses. Nature 2019, 567, 109–112. [Google Scholar] [CrossRef]
- Juozapaitis, M.; Moreira, É.A.; Mena, I.; Giese, S.; Riegger, D.; Pohlmann, A.; Höper, D.; Zimmer, G.; Beer, M.; García-Sastre, A.; et al. An infectious bat-derived chimeric influenza virus harbouring the entry machinery of an influenza A virus. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Ciminski, K.; Ran, W.; Gorka, M.; Lee, J.; Malmlov, A.; Schinköthe, J.; Eckley, M.; Murrieta, R.A.; Aboellail, T.A.; Campbell, C.L.; et al. Bat influenza viruses transmit among bats but are poorly adapted to non-bat species. Nat. Microbiol. 2019, 4, 2298–2309. [Google Scholar] [CrossRef]
- Rasweiler, J.; Badwaik, N.K. Improved procedures for maintaining and breeding the short-tailed fruit bat (Carollia perspicillata) in a laboratory setting. Lab. Anim. 1996, 30, 171–181. [Google Scholar] [CrossRef]
- Rasweiler, J.J.; Cretekos, C.J.; Behringer, R.R. The Short-Tailed Fruit Bat Carollia perspicillata: A Model for Studies in Reproduction and Development. Cold Spring Harb. Protoc. 2009, 2009. [Google Scholar] [CrossRef]
- Rasweiler, J.J.; Cretekos, C.J.; Behringer, R.R. Feeding Short-Tailed Fruit Bats (Carollia perspicillata). Cold Spring Harb. Protoc. 2009, 2009. [Google Scholar] [CrossRef] [PubMed]
- Grund, C.; Hoffmann, D.; Ulrich, R.; Naguib, M.; Schinköthe, J.; Hoffmann, B.; Harder, T.; Saenger, S.; Zscheppang, K.; Tönnies, M.; et al. A novel European H5N8 influenza A virus has increased virulence in ducks but low zoonotic potential. Emerg. Microbes Infect. 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue virus detection by two real-time rt-qpcrs targeting two different genomic segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef]
- Moreira, É.A.; Locher, S.; Kolesnikova, L.; Bolte, H.; Aydillo, T.; García-Sastre, A.; Schwemmle, M.; Zimmer, G. Synthetically derived bat influenza A-like viruses reveal a cell type- but not species-specific tropism. Proc. Natl. Acad. Sci. USA 2016, 113, 12797–12802. [Google Scholar] [CrossRef] [Green Version]
- Yewdell, J.W.; Frank, E.; Gerhard, W. Expression of influenza A virus internal antigens on the surface of infected P815 cells. J. Immunol. 1981, 126, 1814–1819. [Google Scholar]
- Brandtzaeg, P.; Kiyono, H.; Pabst, R.; Russell, M.W. Terminology: Nomenclature of mucosa-associated lymphoid tissue. Mucosal Immunol. 2008, 1, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Schuh, A.J.; Amman, B.R.; Sealy, T.K.; Spengler, J.R.; Nichol, S.T.; Towner, J.S. Egyptian rousette bats maintain long-term protective immunity against Marburg virus infection despite diminished antibody levels. Sci. Rep. 2017, 7, 8763. [Google Scholar] [CrossRef]
- Baker, M.L.; Schountz, T.; Wang, L.F. Antiviral immune responses of bats: A review. Zoonoses Public Health 2013, 60, 104–116. [Google Scholar] [CrossRef]
- Schountz, T.; Baker, M.; Butler, J.; Munster, V.J. Immunological Control of Viral Infections in Bats and the Emergence of Viruses Highly Pathogenic to Humans. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef]
- Campos, A.C.A.; Góes, L.G.B.; Moreira-Soto, A.; De Carvalho, C.; Ambar, G.; Sander, A.-L.; Fischer, C.; Da Rosa, A.R.; De Oliveira, D.C.; Kataoka, A.P.G.; et al. Bat Influenza A(HL18NL11) Virus in Fruit Bats, Brazil. Emerg. Infect. Dis. 2019, 25, 333–337. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Animal No. | 1 | 2 | ||||
|---|---|---|---|---|---|---|
| Method | PCR | ISH | IHC | PCR | ISH | IHC |
| Conchae | 33.6 | 1 # | 1 | - | 0 | 0 |
| Trachea | - | 0 | 0 | - | 0 | 0 |
| Lung | - | 2 | 2 | - | 0 | 0 |
| Heart | - | 0 | 0 | - | 0 | 0 |
| Kidney | - | 0 | 0 | - | 0 | 0 |
| Liver | - | 0 | 0 | - | 0 | 0 |
| Intestine | 34.7 † | 2 * | 2 * | - | 2 * | 1 * |
| Olfactory bulb | - | 0 | 0 | - | 0 | 0 |
| Cerebrum | - | 0 | 0 | - | 0 | 0 |
| Cerebellum | - | 0 | 0 | - | 0 | 0 |
| Muscle | - | nd | nd | - | nd | nd |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorka, M.; Schinköthe, J.; Ulrich, R.; Ciminski, K.; Schwemmle, M.; Beer, M.; Hoffmann, D. Characterization of Experimental Oro-Nasal Inoculation of Seba’s Short-Tailed Bats (Carollia perspicillata) with Bat Influenza A Virus H18N11. Viruses 2020, 12, 232. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020232
Gorka M, Schinköthe J, Ulrich R, Ciminski K, Schwemmle M, Beer M, Hoffmann D. Characterization of Experimental Oro-Nasal Inoculation of Seba’s Short-Tailed Bats (Carollia perspicillata) with Bat Influenza A Virus H18N11. Viruses. 2020; 12(2):232. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020232
Chicago/Turabian StyleGorka, Marco, Jan Schinköthe, Reiner Ulrich, Kevin Ciminski, Martin Schwemmle, Martin Beer, and Donata Hoffmann. 2020. "Characterization of Experimental Oro-Nasal Inoculation of Seba’s Short-Tailed Bats (Carollia perspicillata) with Bat Influenza A Virus H18N11" Viruses 12, no. 2: 232. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020232