TCR Repertoire Characterization for T Cells Expanded in Response to hRSV Infection in Mice Immunized with a Recombinant BCG Vaccine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Animals
2.3. hRSV Infection and rBCG-N-hRSV Immunization
2.4. Flow Cytometry Analyses
2.5. Statistical Analyses
3. Results
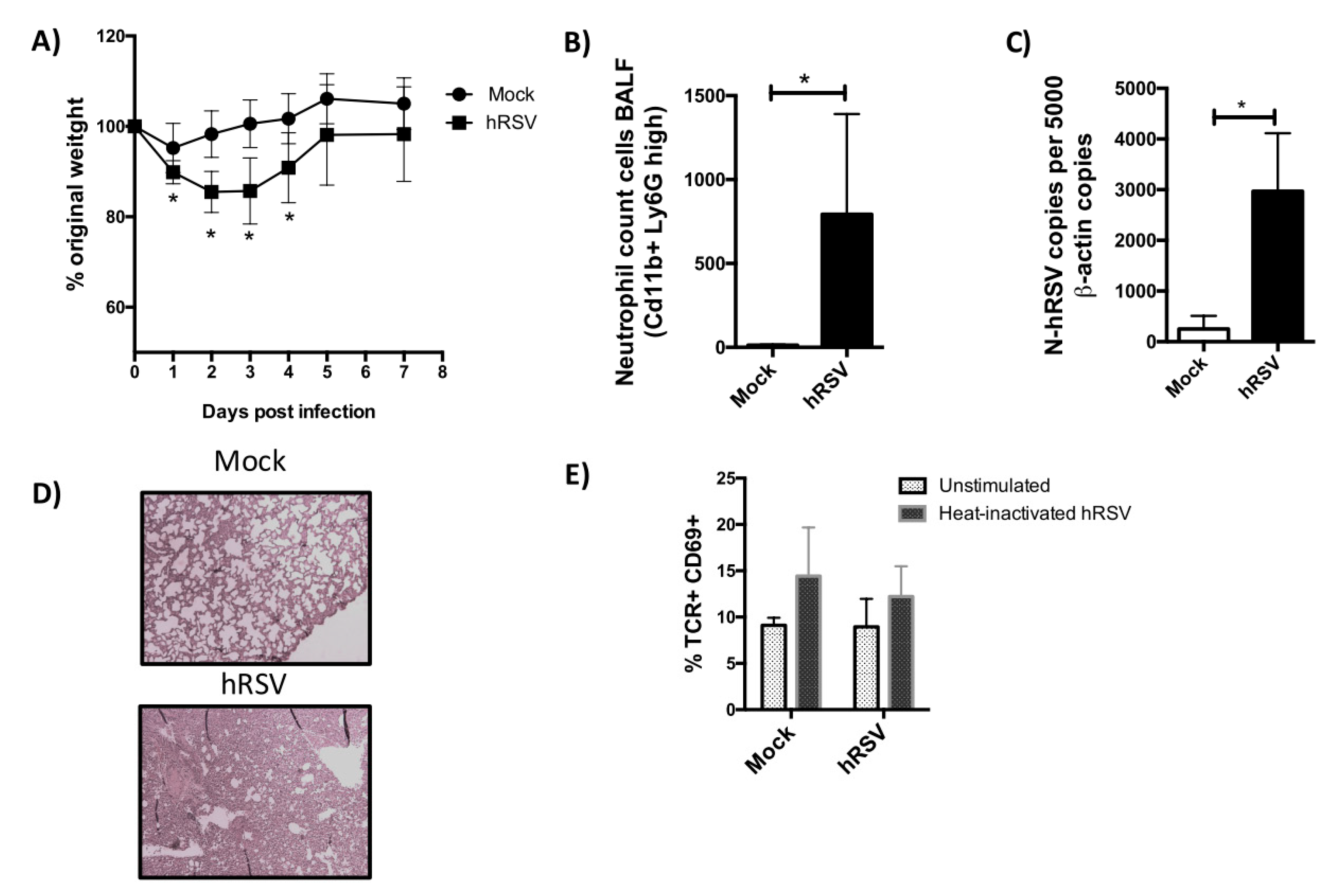
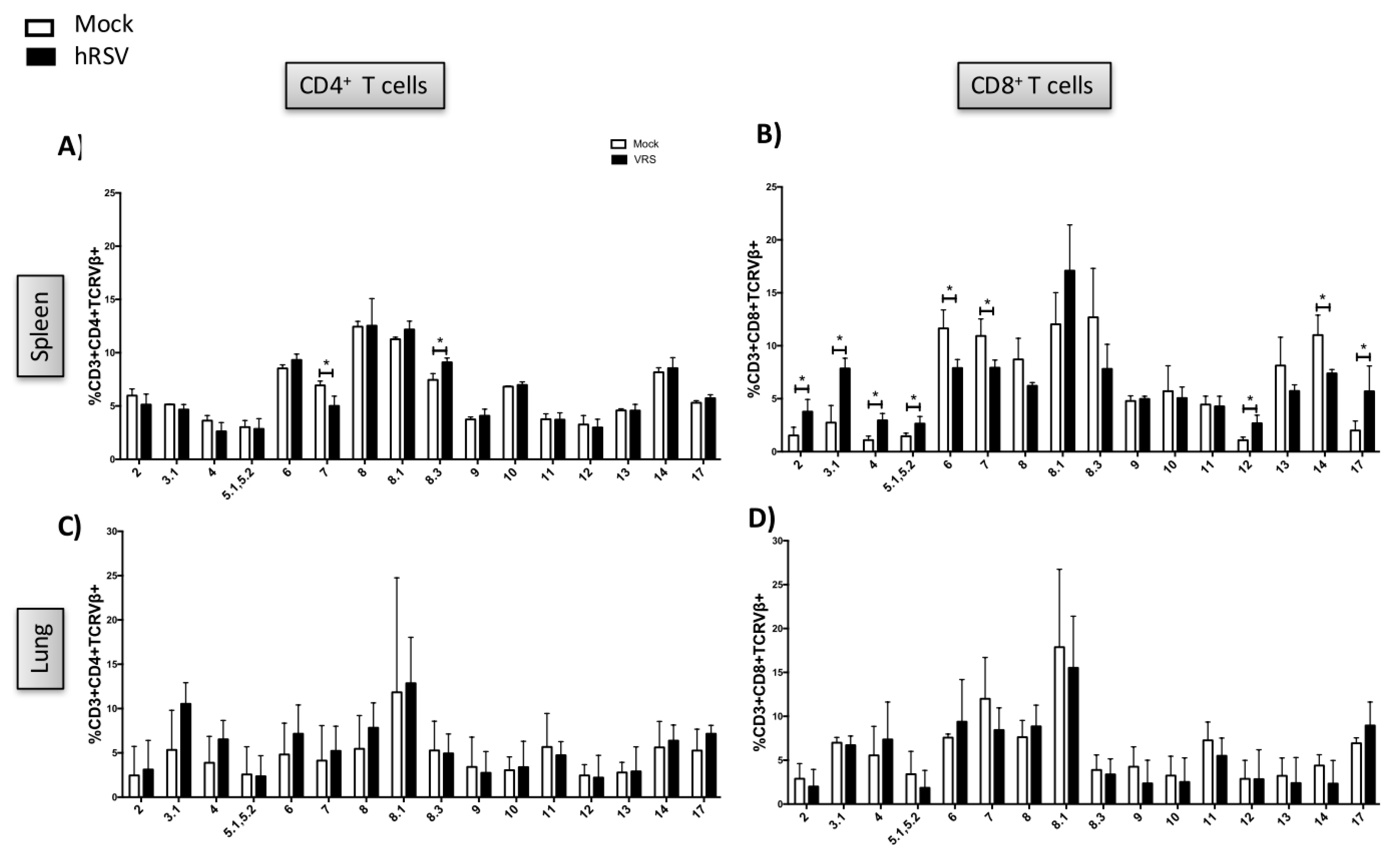
3.1. TCR Repertoire of T Cells from hRSV-Infected Mice
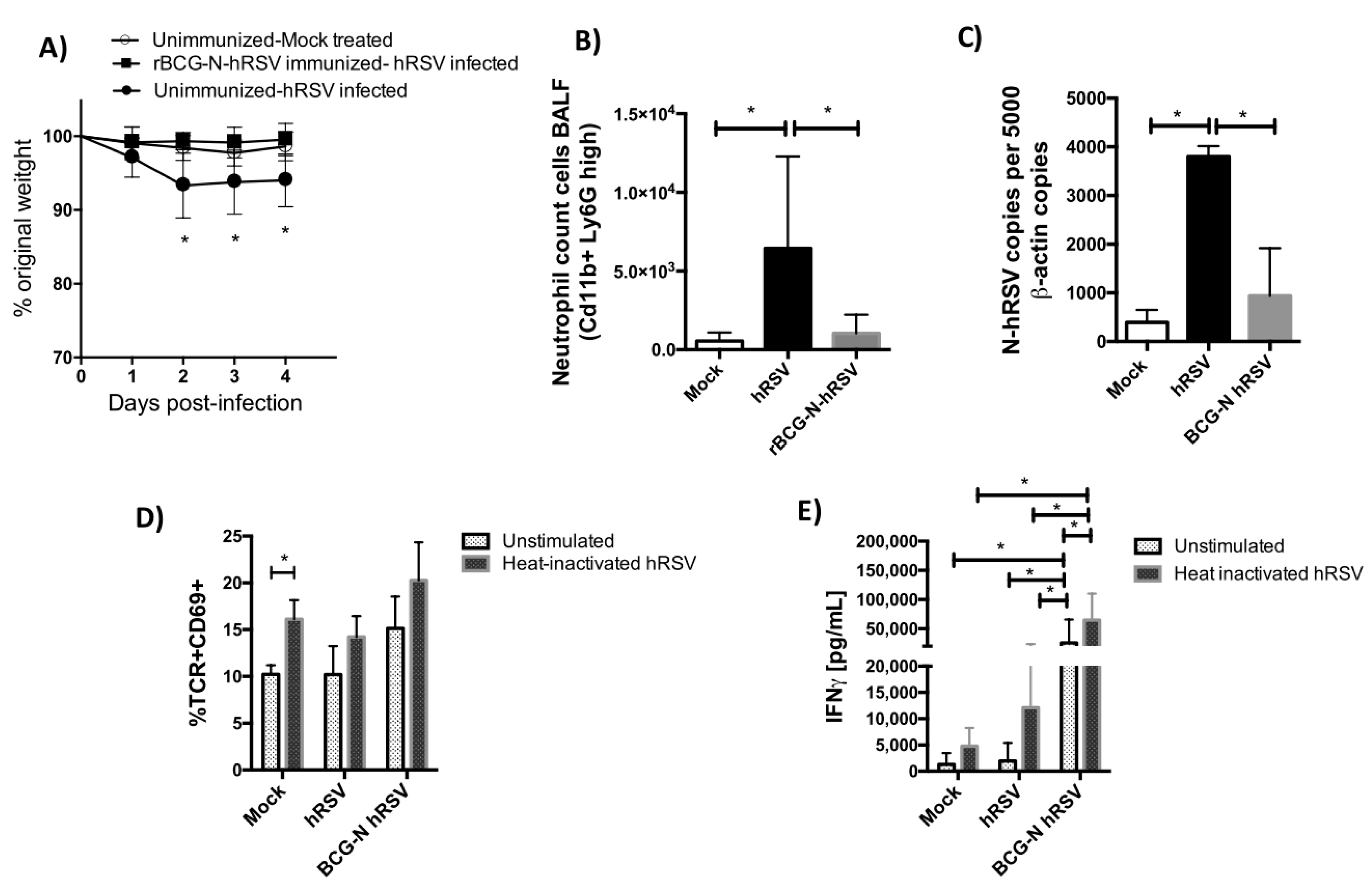
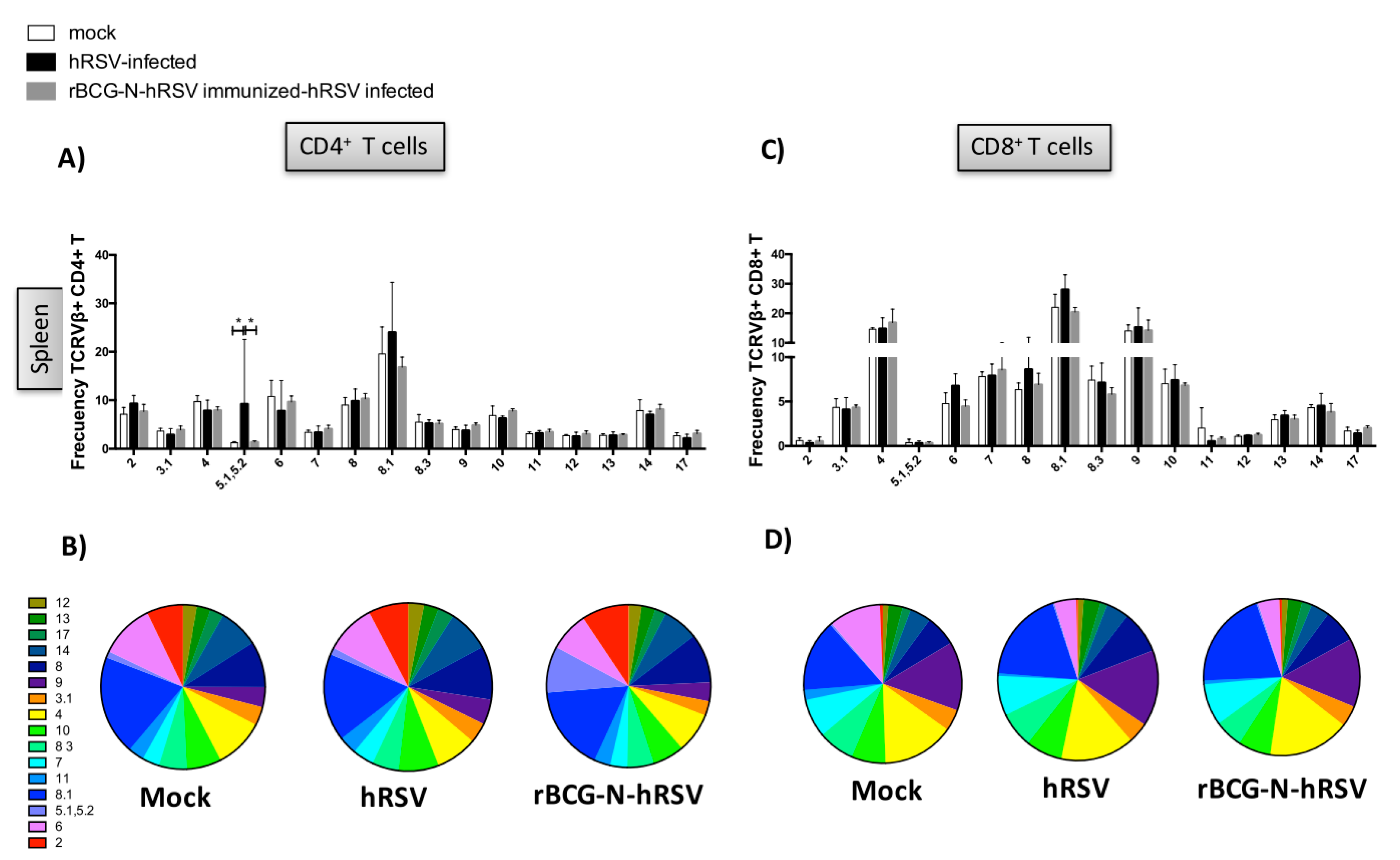
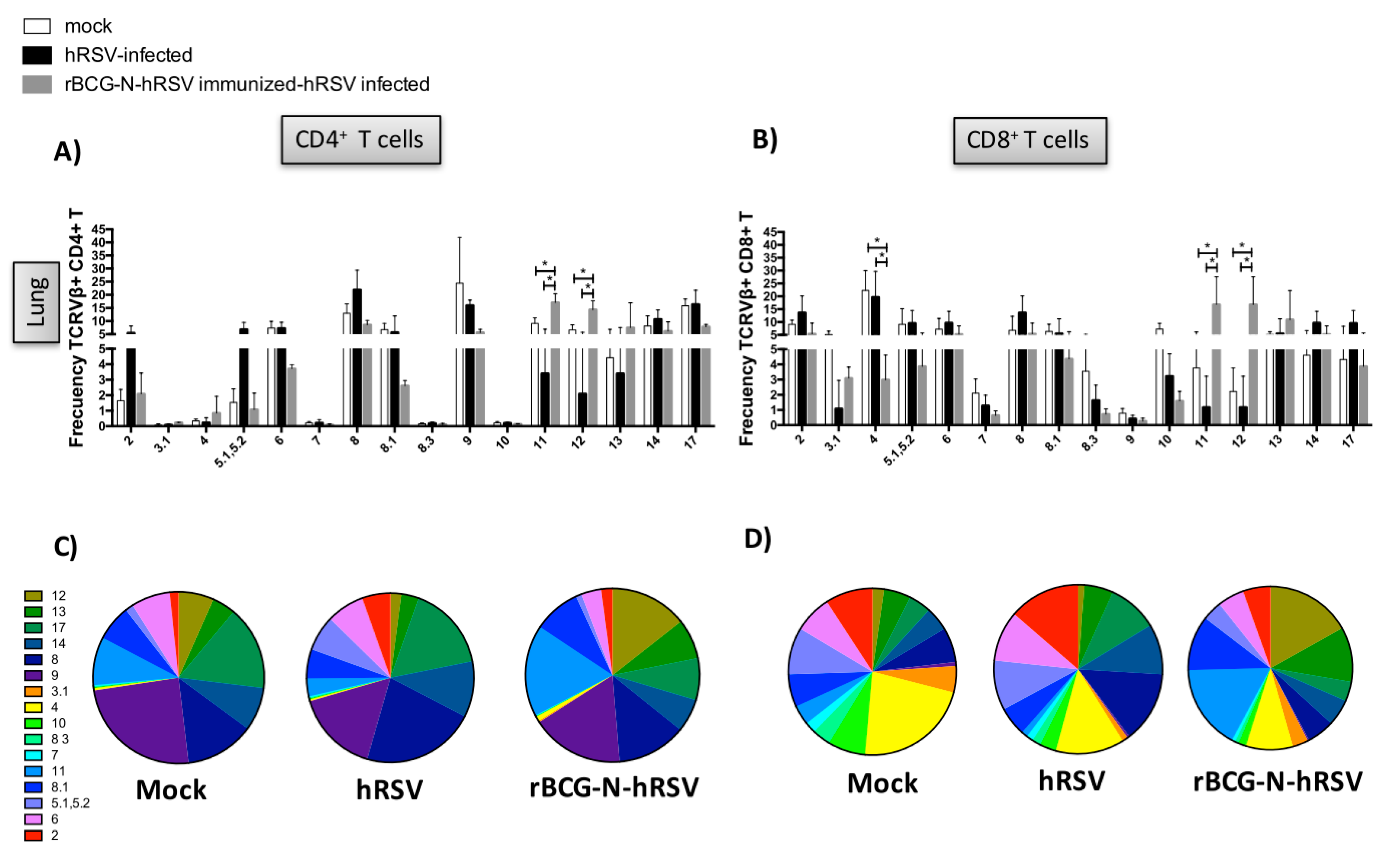
3.2. TCR Repertoire of T Cells from hRSV-Infected Mice Previously Immunized with rBCG-N-hRSV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Bendelja, K.; Gagro, A.; Bace, A.; Lokar-Kolbas, R.; Krsulovic-Hresic, V.; Drazenovic, V.; Mlinaric-Galinovic, G.; Rabatic, S. Predominant type-2 response in infants with respiratory syncytial virus (rsv) infection demonstrated by cytokine flow cytometry. Clin. Exp. Immunol. 2000, 121, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, P.; Lay, M.K.; Piedimonte, G.; Brockmann, P.E.; Palavecino, C.E.; Hernandez, J.; Leon, M.A.; Kalergis, A.M.; Bueno, S.M. Elevated il-3 and il-12p40 levels in the lower airway of infants with rsv-induced bronchiolitis correlate with recurrent wheezing. Cytokine 2015, 76, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Graham, B.S.; Perkins, M.D.; Wright, P.F.; Karzon, D.T. Primary respiratory syncytial virus infection in mice. J. Med. Virol. 1988, 26, 153–162. [Google Scholar] [CrossRef]
- Kong, X.; Hellermann, G.R.; Patton, G.; Kumar, M.; Behera, A.; Randall, T.S.; Zhang, J.; Lockey, R.F.; Mohapatra, S.S. An immunocompromised balb/c mouse model for respiratory syncytial virus infection. Virol. J. 2005, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Bueno, S.M.; Gonzalez, P.A.; Cautivo, K.M.; Mora, J.E.; Leiva, E.D.; Tobar, H.E. Protective t cell immunity against respiratory syncytial virus is efficiently induced by recombinant bcg. Proc. Natl. Acad. Sci. USA 2008, 105, 20822–20827. [Google Scholar] [CrossRef] [Green Version]
- Graham, B.S.; Bunton, L.A.; Wright, P.F.; Karzon, D.T. Role of t lymphocyte subsets in the pathogenesis of primary infection and rechallenge with respiratory syncytial virus in mice. J. Clin. Investig. 1991, 88, 1026–1033. [Google Scholar] [CrossRef]
- Cautivo, K.M.; Bueno, S.M.; Cortes, C.M.; Wozniak, A.; Riedel, C.A.; Kalergis, A.M. Efficient lung recruitment of respiratory syncytial virusspecific th1 cells induced by recombinant bacillus calmette-guerin promotes virus clearance and protects from infection. J. Immunol. 2010, 185, 7633–7645. [Google Scholar] [CrossRef] [Green Version]
- Céspedes, P.C.; Rey-Jurado, E.; Espinoza, J.A.; Rivera, C.A.; Canedo-Marroquín, G.; Bueno, S.M.; Kalergis, A. A single, low dose of a cgmp recombinant bcg vaccine elicits protective t cell immunity against the human respiratory syncytial virus infection and prevents lung pathology in mice. Vaccine 2017, 35, 757–766. [Google Scholar]
- Hodges, E.; Krishna, M.T.; Pickard, C.; Smith, J.L. Diagnostic role of tests for t cell receptor (tcr) genes. J. Clin. Pathol. 2003, 56, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Arstila, T.P.; Casrouge, A.; Baron, V.; Even, J.; Kanellopoulos, J.; Kourilsky, P. A direct estimate of the human alphabeta t cell receptor diversity. Science 1999, 286, 958–961. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Summer, R.; Wilson, A.A.; Kotton, D.N.; Fine, A. The prolonged life-span of alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 2008, 38, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, J.S.; Wearsch, P.A.; Cresswell, P. Pathways of antigen processing. Annu. Rev. Immunol. 2013, 31, 443–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.; Srikiatkhachorn, A.; Braciale, T.J. Visualization and characterization of respiratory syncytial virus f-specific cd8(+) t cells during experimental virus infection. J. Immunol. 2001, 167, 4254–4260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Borthwick, N.J.; Morrison, P.; Gao, G.F.; Steward, M.W. Virus-specific ctl responses induced by an h-2k(d)-restricted, motif-negative 15-mer peptide from the fusion protein of respiratory syncytial virus. J. Gen. Virol. 2002, 83, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnstone, C.; de León, P.; Medina, F.; Melero, J.A.; García-Barreno, B.; Del Val, M. Shifting immunodominance pattern of two cytotoxic t-lymphocyte epitopes in the f glycoprotein of the long strain of respiratory syncytial virus. J. Gen. Virol. 2004, 85, 3229–3238. [Google Scholar] [CrossRef]
- Kulkarni, A.B.; Collins, P.L.; Bacik, I.; Yewdell, J.W.; Bennink, J.R.; Crowe, J.E.; Murphy, B.R. Cytotoxic t cells specific for a single peptide on the m2 protein of respiratory syncytial virus are the sole mediators of resistance induced by immunization with m2 encoded by a recombinant vaccinia virus. J. Virol. 1995, 69, 1261–1264. [Google Scholar] [CrossRef] [Green Version]
- Gil, A.; Yassai, M.B.; Naumov, Y.N.; Selin, L.K. Narrowing of human influenza a virus-specific t cell receptor α and β repertoires with increasing age. J. Virol. 2015, 89, 4102–4116. [Google Scholar] [CrossRef] [Green Version]
- Miles, J.J.; Thammanichanond, D.; Moneer, S.; Nivarthi, U.K.; Kjer-Nielsen, L.; Tracy, S.L.; Aitken, C.K.; Brennan, R.M.; Zeng, W.; Marquart, L.; et al. Antigen-driven patterns of tcr bias are shared across diverse outcomes of human hepatitis c virus infection. J. Immunol. 2011, 186, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, D.M.; Valderrama, S.; Gualtero, S.; Hernandez, C.; Lopez, M.; Herrera, M.V.; Solano, J.; Fiorentino, S.; Quijano, S. Loss of t-cell multifunctionality and tcr-vbeta repertoire against epstein-barr virus is associated with worse prognosis and clinical parameters in hiv(+) patients. Front. Immunol. 2018, 9, 2291. [Google Scholar] [CrossRef] [Green Version]
- de Vos van Steenwijk, P.J.; Heusinkveld, M.; Ramwadhdoebe, T.H.; Lowik, M.J.; van der Hulst, J.M.; Goedemans, R.; Piersma, S.J.; Kenter, G.G.; van der Burg, S.H. An unexpectedly large polyclonal repertoire of hpv-specific t cells is poised for action in patients with cervical cancer. Cancer Res. 2010, 70, 2707–2717. [Google Scholar] [CrossRef] [Green Version]
- Varga, S.M.; Wang, X.; Welsh, R.M.; Braciale, T.J. Immunopathology in rsv infection is mediated by a discrete oligoclonal subset of antigen-specific cd4(+) t cells. Immunity 2001, 15, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Bar-Haim, E.; Erez, N.; Malloy, A.M.; Graham, B.S.; Ruckwardt, T.J. Cd8+ tcr transgenic strains expressing public versus private tcr targeting the respiratory syncytial virus k(d)m2(82-90) epitope demonstrate similar functional profiles. PLoS ONE 2014, 9, e99249. [Google Scholar] [CrossRef] [PubMed]
- Vallbracht, S.; Jessen, B.; Mrusek, S.; Enders, A.; Collins, P.L.; Ehl, S.; Krempl, C.D. Influence of a single viral epitope on t cell response and disease after infection of mice with respiratory syncytial virus. J. Immunol. 2007, 179, 8264–8273. [Google Scholar] [CrossRef]
- Cornberg, M.; Chen, A.T.; Wilkinson, L.A.; Brehm, M.A.; Kim, S.K.; Calcagno, C.; Ghersi, D.; Puzone, R.; Celada, F.; Welsh, R.M.; et al. Narrowed tcr repertoire and viral escape as a consequence of heterologous immunity. J. Clin. Investig. 2006, 116, 1443–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, J.A.; Galvez, N.; Rivera, C.A.; Palavecino, C.E.; Céspedes, P.F.; Rey-Jurado, E.; Bueno, S.M.; Kalergis, A.M. Recombinant bcg vaccines reduce pneumovirus-caused airway pathology by inducing protective cellular and humoral immunity rbcg induces protection against pneumoviruses. Submitt. Front. Immunol. 2018, 9, 2875. [Google Scholar] [CrossRef] [Green Version]
- Herrada, A.A.; Contreras, F.J.; Tobar, J.A.; Pacheco, R.; Kalergis, A.M. Immune complex-induced enhancement of bacterial antigen presentation requires fcγ receptor iii expression on dendritic cells. Proc. Natl. Acad. Sci. USA 2007, 104, 13402–13407. [Google Scholar] [CrossRef] [Green Version]
- Marchant, A.; Goetghebuer, T.; Ota, M.O.; Wolfe, I.; Ceesay, S.J.; De Groote, D.; Corrah, T.; Bennett, S.; Wheeler, J.; Huygen, K.; et al. Newborns develop a th1-type immune response to mycobacterium bovis bacillus calmette-guerin vaccination. J. Immunol. 1999, 163, 2249–2255. [Google Scholar]
- Nakatsugawa, M.; Rahman, M.A.; Yamashita, Y.; Ochi, T.; Wnuk, P.; Tanaka, S.; Chamoto, K.; Kagoya, Y.; Saso, K.; Guo, T.; et al. Cd4(+) and cd8(+) tcrβ repertoires possess different potentials to generate extraordinarily high-avidity t cells. Sci. Rep. 2016, 6, 23821. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Miller, S.A.; Wright, D.W.; Rock, M.T.; Crowe, J.E., Jr. Tissue-specific regulation of cd8+ t-lymphocyte immunodominance in respiratory syncytial virus infection. J. Virol. 2007, 81, 2349–2358. [Google Scholar] [CrossRef] [Green Version]
- Fozza, C.; Corda, G.; Virdis, P.; Contini, S.; Barraqueddu, F.; Galleu, A.; Isoni, A.; Cossu, A.; Dore, F.; Careddu, M.G.; et al. Derangement of the t-cell repertoire in patients with b-cell non-hodgkin’s lymphoma. Eur. J. Haematol. 2015, 94, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Salameire, D.; Solly, F.; Fabre, B.; Lefebvre, C.; Chauvet, M.; Gressin, R.; Corront, B.; Ciapa, A.; Pernollet, M.; Plumas, J.; et al. Accurate detection of the tumor clone in peripheral t-cell lymphoma biopsies by flow cytometric analysis of tcr-vβ repertoire. Mod. Pathol. 2012, 25, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.C.; Dash, P.; McCullers, J.A.; Doherty, P.C.; Thomas, P.G. T cell receptor αβ diversity inversely correlates with pathogen-specific antibody levels in human cytomegalovirus infection. Sci. Transl. Med. 2012, 4, 128ra142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer-Olson, D.; Shoukry, N.H.; Brady, K.W.; Kim, H.; Olson, D.P.; Hartman, K.; Shintani, A.K.; Walker, C.M.; Kalams, S.A. Limited t cell receptor diversity of hcv-specific t cell responses is associated with ctl escape. J. Exp. Med. 2004, 200, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Koning, D.; Ladell, K.; McLaren, J.E.; Grady, B.P.; Schellens, I.M.; Ham, P.v.; Nijhuis, M.; Borghans, J.A.; Keşmir, C.; et al. Complex t-cell receptor repertoire dynamics underlie the cd8+ t-cell response to hiv-1. J. Virol. 2015, 89, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, W.; Reinherz, E.L. In vivo selection of a tcr vβ repertoire directed against an immunodominant influenza virus ctl epitope. Int. Immunol. 2004, 16, 1549–1559. [Google Scholar] [CrossRef]
- Pizzolla, A.; Nguyen, T.H.; Sant, S.; Jaffar, J.; Loudovaris, T.; Mannering, S.I.; Thomas, P.G.; Westall, G.P.; Kedzierska, K.; Wakim, L.M. Influenza-specific lung-resident memory t cells are proliferative and polyfunctional and maintain diverse tcr profiles. J. Clin. Investig. 2018, 128, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Retamal-Diaz, A.; Covian, C.; Pacheco, G.A.; Castiglione-Matamala, A.T.; Bueno, S.M.; Gonzalez, P.A.; Kalergis, A.M. Contribution of resident memory cd8(+) t cells to protective immunity against respiratory syncytial virus and their impact on vaccine design. Pathogens 2019, 8, 147. [Google Scholar] [CrossRef] [Green Version]
- Hogan, L.H.; Macvilay, K.; Barger, B.; Co, D.; Malkovska, I.; Fennelly, G.; Sandor, M. Mycobacterium bovis strain bacillus calmette-guerin-induced liver granulomas contain a diverse tcr repertoire, but a monoclonal t cell population is sufficient for protective granuloma formation. J. Immunol. 2001, 166, 6367–6375. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rey-Jurado, E.; Bohmwald, K.; Correa, H.G.; Kalergis, A.M. TCR Repertoire Characterization for T Cells Expanded in Response to hRSV Infection in Mice Immunized with a Recombinant BCG Vaccine. Viruses 2020, 12, 233. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020233
Rey-Jurado E, Bohmwald K, Correa HG, Kalergis AM. TCR Repertoire Characterization for T Cells Expanded in Response to hRSV Infection in Mice Immunized with a Recombinant BCG Vaccine. Viruses. 2020; 12(2):233. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020233
Chicago/Turabian StyleRey-Jurado, Emma, Karen Bohmwald, Hernán G. Correa, and Alexis M. Kalergis. 2020. "TCR Repertoire Characterization for T Cells Expanded in Response to hRSV Infection in Mice Immunized with a Recombinant BCG Vaccine" Viruses 12, no. 2: 233. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020233