Field Trial Vaccination against Cowpox in Two Alpaca Herds

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Alpaca Herds
2.2. Herd History
2.3. MVA Vaccination
2.4. Sampling
2.5. Diagnostic Methods
2.6. Statistical Analysis
3. Results
3.1. Safety
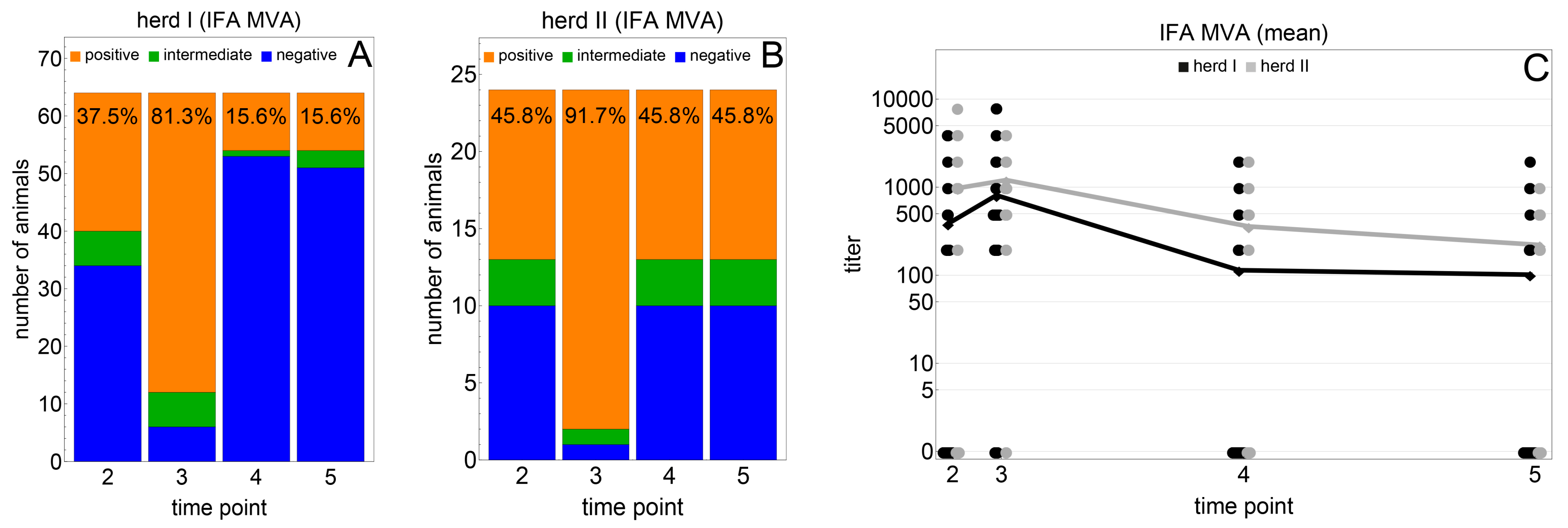
3.2. Immunogenicity
3.3. Results of Crias
3.4. Results of Swab Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Essbauer, S.; Pfeffer, M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef]
- Eulenberger, K.; Bernhard, A.; Nieper, H.; Hoffmann, K.; Scheller, R.; Meyer, H.; Zimmermann, P.; Kiessling, J. An outbreak of cowpox virus infection in a black rhino (diceros bicornis) at Leipzig Zoo. Verh ber Erkrg Zootiere 2005, 42, 77–85. [Google Scholar]
- Knieriem, A. Pockenerkrankung eines Asiatischen Elefantenbullen—Fallbericht einer generalisierten Infektion. Tagungsbericht Arbeitstagung der Zootierärzte im Deutschsprachigen Raum 2002, 21, 9–14. [Google Scholar]
- Pilaski, J.; Jacoby, F. Die Kuhpocken-Erkrankungen der Zootiere. Verh ber Erkrg Zootiere 1993, 35, 39–50. [Google Scholar]
- Pilaski, J.; Von Witzendorff, P.; Brandt, H.-P.; Höhr, D. Ein Pockenausbruch bei Elefanten (Elephas maximus, Loxodonta africans) in einem Wanderzirkus während des Aufenthaltes im Winterquartier. Verh ber Erkrg Zootiere 1995, 37, 357–363. [Google Scholar]
- Schaller, K.; Pilaski, J. Pocken bei Breitmaulnashörnern (Ceratotherium s. simum) im Zoologischen Garten Münster. Zool. Garten N. F. 1979, 49, 169–184. [Google Scholar]
- Kalthoff, D.; Bock, W.-I.; Hühn, F.; Beer, M.; Hoffmann, B. Fatal cowpox virus infection in cotton-top tamarins (Saguinus oedipus) in Germany. Vector Borne Zoonotic Dis. (Larchmt. N. Y.) 2014, 14, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Kik, M.J.L.; Liu, P.L.; Van Asten, J.A.M. Cowpoxvirus infection in the Patagonian cavy (Dolichotis patagonum) emerging disease in an educational animal park the first reported case. Vet. Q. 2006, 28, 42–44. [Google Scholar] [CrossRef]
- Kurth, A.; Straube, M.; Kuczka, A.; Dunsche, A.J.; Meyer, H.; Nitsche, A.; Art, F.Y.P. Cowpox Virus Outbreak in Banded Mongooses (Mungos mungo) and Jaguarundis (Herpailurus yagouaroundi) with a Time-Delayed Infection to Humans. PLoS ONE 2009, 4, e6883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marennikova, S.S.; Maltseva, N.N.; Korneeva, V.I.; Garanina, N.M. Outbreak of Pox Disease among Carnivora (Felidae) and Edentata. J. Infect. Dis. 1977, 135, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Mätz-Rensing, K.; Ellerbrok, H.; Ehlers, B.; Pauli, G.; Floto, A.; Alex, M.; Czerny, C.-P.; Kaup, F.-J. Fatal poxvirus outbreak in a colony of New World monkeys. Vet. Pathol. 2006, 43, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Schüppel, K.-F.; Menger, S.; Eulenberger, K.; Bernhard, A.; Pilaski, J. Kuhpockeninfektion bei Alpakas (Lama Glama pacos). Verh ber Erkrg Zootiere 1997, 38, 259–265. [Google Scholar]
- Cardeti, G.; Brozzi, A.; Eleni, C.; Polici, N.; D’Alterio, G.; Carletti, F.; Scicluna, M.T.; Castiletti, C.; Capobianchi, M.R.; Di Caro, A.; et al. Cowpox Virus in Llama, Italy. Emerg. Infect. Dis. 2011, 17, 1513–1515. [Google Scholar] [CrossRef] [PubMed]
- Goerigk, D.; Theuß, T.; Pfeffer, M.; Konrath, A.; Kalthoff, D.; Woll, D.; Vahlenkamp, T.W.; Beer, M.; Starke, A. Kuhpockenvirusinfektion bei einem Alpaka (Vicugna pacos)-klinische Symptomatik, Diagnostik und pathologische Befunde. Tierärztl Prax 2014, 42, 169–177. [Google Scholar]
- Franke, A.; Pfaff, F.; Jenckel, M.; Hoffmann, B.; Höper, D.; Antwerpen, M.; Meyer, H.; Beer, M.; Hoffmann, D. Classification of Cowpox Viruses into Several Distinct Clades and Identification of a Novel Lineage. Viruses 2017, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Prkno, A.; Hoffmann, D.; Goerigk, D.; Kaiser, M.; Van Maanen, A.C.F.; Jeske, K.; Jenckel, M.; Pfaff, F.; Vahlenkamp, T.W.; Beer, M.; et al. Epidemiological Investigations of Four Cowpox Virus Outbreaks in Alpaca Herds, Germany. Viruses 2017, 9, 344. [Google Scholar] [CrossRef] [Green Version]
- Prkno, A.; Kaiser, M.; Goerigk, D.; Pfeffer, M.; Vahlenkamp, T.W.; Hoffmann, D.; Beer, M.; Starke, A. Klinisches Erscheinungsbild der Kuhpockenvirus-infektion bei Neuweltkameliden. Tierärztl Prax 2018, 46, 50–56. [Google Scholar] [CrossRef]
- Foster, S.A.; Parker, S.; Lanier, R. The Role of Brincidofovir in Preparation for a Potential Smallpox Outbreak. Viruses 2017, 9, 320. [Google Scholar] [CrossRef] [Green Version]
- Mucker, E.M.; Goff, A.J.; Shamblin, J.D.; Grosenbach, D.W.; Damon, I.K.; Mehal, J.M.; Holman, R.C.; Carroll, D.; Gallardo, N.; Olson, V.A.; et al. Efficacy of tecovirimat (ST-246) in nonhuman primates infected with variola virus (Smallpox). Antimicrob. Agents Chemother. 2013, 57, 6246–6253. [Google Scholar] [CrossRef] [Green Version]
- Quenelle, D.C.; Kern, E.R. Treatment of Vaccinia and Cowpox Virus Infections in Mice with CMX001 and ST-246. Viruses 2010, 2, 2681–2695. [Google Scholar] [CrossRef] [Green Version]
- Smee, D.F. Orthopoxvirus inhibitors that are active in animal models: An update from 2008 to 2012. Future Virol. 2013, 8, 891–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Pevear, D.C.; Davies, M.H.; Collett, M.S.; Bailey, T.; Rippen, S.; Barone, L.; Burns, C.; Rhodes, G.; Tohan, S.; et al. An orally bioavailable antipoxvirus compound (ST-246) inhibits extracellular virus formation and protects mice from lethal orthopoxvirus Challenge. J. Virol. 2005, 79, 13139–13149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, H. Summary Report on First, Second and Third Generation Smallpox Vaccines. 2013. Available online: http://www.who.int/immunization/sage/meetings/2013/november/2_Smallpox_vaccine_review_updated_11_10_13.pdf (accessed on 28 April 2018).
- Mayr, A.; Hochstein-Mintzel, V.; Stickl, H. Abstammung, Eigenschaften und Verwendung des attenuierten Vaccinia-Stammes MVA. Infection 1975, 3, 6–14. [Google Scholar] [CrossRef]
- Mahnel, H.; Mayr, A. Erfahrungen bei der Schutzimpfung gegen Orthopocken von Mensch und Tier mit dem Impfstamm MVA. Berl. Münchener Tierärztliche Wochenschr. 1994, 107, 253–256. [Google Scholar]
- Meyer, H.; Sutter, G.; Mayr, A. Mapping of deletions in the genome of the highly attenuated vaccinia virus MVA and their influence on virulence. J. Gen. Virol. 1991, 72(Pt. 5), 1031–1038. [Google Scholar] [CrossRef]
- Mayr, A.; Stickl, H.; Müller, H.K.; Danner, K.; Singer, H. Der Pockenimpfstamm MVA: Marker, genetische Struktur, Erfahrungen mit der parenteralen Schutzimpfung und Verhalten im abwehrgeschwächten Organismus. Zentralblatt fur Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. Erste Abteilung Originale. Reihe B Hyg. Betr. Prav. Med. 1978, 167, 375–390. [Google Scholar]
- Greenberg, R.N.; Hurley, M.Y.; Hurley, Y.; Dinh, D.V.; Mraz, S.; Vera, J.G.; Von Bredow, D.; Von Krempelhuber, A.; Roesch, S.; Virgin, G.; et al. A Multicenter, Open-Label, Controlled Phase II Study to Evaluate Safety and Immunogenicity of MVA Smallpox Vaccine (IMVAMUNE) in 18-40 Year Old Subjects with Diagnosed Atopic Dermatitis. PLoS ONE 2015, 10, e0138348. [Google Scholar]
- European Medicines Agency. European Public Assessment Report—IMVANEX, Common Name–Modified Vaccinia Ankara Virus, Anhang I—Zusammenfassung der Merkmale des Arzneimittels. Available online: http://www.ema.europa.eu/docs/de_DE/document_library/EPAR_-_Product_Information/human/002596/WC500147896.pdf (accessed on 5 June 2018).
- Vaughan, A.; Aarons, E.; Astbury, J.; Balasegaram, S.; Beadsworth, M.; Beck, C.R.; Chand, M.; O’Connor, C.; Dunning, J.; Ghebrehewet, S.; et al. Two cases of monkeypox imported to the United Kingdom, September 2018. Eurosurveillance 2018, 23, 1800509. [Google Scholar] [CrossRef]
- Pilaski, J.; Zhou, X. Die Pockenimpfung der Elefanten. Verh ber Erkrg Zootiere 1991, 33, 203–211. [Google Scholar]
- Cardeti, G.; Gruber, C.E.M.; Eleni, C.; Carletti, F.; Castilletti, C.; Manna, G.; Rosone, F.; Giombini, E.; Selleri, M.; Lapa, D.; et al. Fatal Outbreak in Tonkean Macaques Caused by Possibly Novel Orthopoxvirus, Italy, January 2015 1. Emerg. Infect. Dis. 2017, 23, 1941–1949. [Google Scholar] [CrossRef] [Green Version]
- Lauer, J. Untersuchungen über die Wirksamkeit und Unschädlichkeit einer Schutzimpfung von Katzen gegen Pocken mit MVA-Lebendimpfstoff. Ph.D. Thesis, Ludwig Maximilian University of Munich, Munich, Bavaria, Germany, 1993. [Google Scholar]
- Munz, E.; Linckh, S.; Renner-Müller, I.C.; Reimann, M. Die Wirksamkeit einer Immunisierung mit Vaccinia-Virus Stamm “MVA” gegen eine Infektion mit Kuhpocken-Virus Stamm “OPV 85” beim Kaninchen. Zent. Vet. B 1993, 40, 131–140. [Google Scholar] [CrossRef]
- Härdi-Landerer, M.C.; Leu, M.; Steiner, A. Der polyvalente Moderhinke-Impfstoff im Praxistest. Tierärztl Prax 2012, 40, 294–300. [Google Scholar]
- Hoffmann, D.; Franke, A.; Jenckel, M.; Tamošiūnaitė, A.; Schluckebier, J.; Granzow, H.; Hoffmann, B.; Fischer, S.; Ulrich, R.G.; Höper, D.; et al. Out of the Reservoir: Phenotypic and Genotypic Characterization of a Novel Cowpox Virus Isolated from a Common Vole. J. Virol. 2015, 89, 10959–10969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksyutov, R.A.; Gavrilova, E.V.; Meyer, H.; Shchelkunov, S.N. Real-time PCR assay for specific detection of cowpox virus. J. Virol. Methods 2015, 211, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Earl, P.L.; Americo, J.L.; Wyatt, L.S.; Eller, L.A.; Whitbeck, J.C.; Cohen, G.H.; Eisenberg, R.J.; Hartmann, C.J.; Jackson, D.L.; Kulesh, D.A.; et al. Immunogenicity of a highly attenuated MVA smallpox vaccine and protection against monkeypox. Nature 2004, 428, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Grandpre, L.E.; Duke-Cohan, J.S.; Ewald, B.A.; Devoy, C.; Barouch, D.H.; Letvin, N.L.; Reinherz, E.L.; Baden, L.R.; Dolin, R.; Seaman, M.S. Immunogenicity of recombinant Modified Vaccinia Ankara following a single or multi-dose vaccine regimen in rhesus monkeys. Vaccine 2009, 27, 1549–1556. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, R.N.; Hay, C.M.; Stapleton, J.T.; Marbury, T.C.; Wagner, E.; Kreitmeir, E.; Röesch, S.; Von Krempelhuber, A.; Young, P.; Nichols, R.; et al. A Randomized, Double-Blind, Placebo-Controlled Phase II Trial Investigating the Safety and Immunogenicity of Modified Vaccinia Ankara Smallpox Vaccine (MVA-BN®) in 56-80-Year-Old Subjects. PLoS ONE 2016, 11, e0157335. [Google Scholar] [CrossRef]
- Zanolari, P. Neuweltkameliden—Von der Geburtsvorbereitung bis zur Versorgung der Neugeborenen. Forum 2006, 12, 6–12. [Google Scholar]
- Van Maanen, C.; Bruin, G.; De Boer-Luijtze, E.; Smolders, G.; De Boer, G.F. Interference of maternal antibodies with the immune response of foals after vaccination against equine influenza. Vet. Q. 1992, 14, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Price, P.J.R.; Torres-Domínguez, L.E.; Brandmüller, C.; Sutter, G.; Lehmann, M.H. Modified Vaccinia virus Ankara: Innate immune activation and induction of cellular signalling. Vaccine 2013, 31, 4231–4234. [Google Scholar] [CrossRef]


{kind=link}
| Time Points | 1 (Day Zero) | 2 (+1 Mo) | 3 (+1 Mo) | 4 (+5 Mo) | 5 (+6 Mo) |
|---|---|---|---|---|---|
| Vaccination | × | × | − | − | − |
| Adverse reaction | − | × | × | − | − |
| Clinical examination | × | × | × | × | × |
| Serum sample (antibody testing) | − | × | × | × | × |
| Swab sample (viral DNA) | − | × | × | × | × |
| Gr | 1 | 2 | 3 | 4 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TP | P | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | total |
| 2 | IFA (MVA) | + | + | − | + | − | ? | − | ? | − | + | ? | − | − | + | − | − | + | ? | |
| 3 | + | + | + | + | + | + | + | + | + | − | ? | ? | − | + | + | + | + | + | ||
| 4 | + | + | + | − | + | ? | ? | − | − | − | − | − | − | − | − | − | − | − | ||
| 5 | + | ? | + | − | − | − | − | − | − | − | − | − | − | + | + | ? | ? | ? | ||
| n herd I | 8 | 1 | 13 | 1 | 1 | 4 | 20 | 1 | 1 | 5 | 5 | 1 | 1 | 1 | 1 | 64 | ||||
| n herd II | 9 | 1 | 1 | 2 | 1 | 1 | 4 | 1 | 1 | 1 | 1 | 1 | 24 | |||||||
| n total | 17 | 2 | 1 | 13 | 1 | 2 | 2 | 5 | 24 | 1 | 1 | 6 | 6 | 1 | 2 | 2 | 1 | 1 | 88 | |
| Gr | 3 | 4 | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TP | P | 11 | 12 | 13 | 14 | 15 | 15 | 16 | 16 | 17 | 18 | ||||||||||
| 2 | IFA (MVA) | ? | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − | − | − | + | ? |
| 3 | ? | ? | ? | ? | ? | ? | ? | − | − | − | − | − | − | + | + | + | + | + | + | + | |
| 4 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | |
| 5 | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + | ? | ? | ? | ? | |
| ID | 149 | 24 | 104 | 115 | 121 | 140 | 153 | 23 | 126 | 127 | 143 | 151 | 163 | 155 | 10 | 152 | 11 | 137 | 6 | 144 | |
| 2 | IFA (CPXV) | − | − | + | − | ? | − | − | + | − | + | − | − | − | + | + | − | + | ? | + | ? |
| 3 | + | + | + | + | + | ? | − | + | + | + | − | − | − | + | + | + | + | + | + | + | |
| 4 | − | + | + | ? | + | ? | − | − | − | + | + | − | − | + | ? | + | ? | + | + | + | |
| 5 | − | ? | − | ? | + | − | ? | − | ? | ? | − | − | − | + | + | + | − | + | + | + | |
| ID | HERD | SEX | AGE * | IFA Initial Sampling | MVA-VAC | IFA TP 5 | Titre MARE ** | MARE INTERVAL *** |
|---|---|---|---|---|---|---|---|---|
| 4 | II | m | 3.3 | 1:500 | + | − | + | <6 mo |
| 5 | II | f | 3.7 | 1:500 | + | − | + | <6 mo |
| 28 | II | f | 5.1 | 1:200 | + | − | + | <6 mo |
| 29 | II | f | 3 | <1:200 | + | − | − | <6 mo |
| 30 | II | m | 2.9 | 1:1,000 | + | − | + | <6 mo |
| 31 | II | f | 6.6 | >0<1:200 | + | − | + | <6 mo |
| 38° | II | m | 6.6 | 1:200 | − | + | + | >6 mo |
| 39° | II | m | 12.3 | <1:200 | − | − | + | >6 mo |
| 40° | II | f | 7.3 | >0<1:200 | − | ? | + | >6 mo |
| 168 | I | m | 7.6 | <1:200 | − | − | − | >6 mo |
| 169 | I | m | 3.7 | <1:200 | − | − | − | >6 mo |
| 170 | I | f | 5.1 | <1:200 | − | − | − | >6 mo |
| 171 | I | f | 3.6 | 1:500 | − | − | + | >6 mo |
| 172 | I | f | 4.3 | 1:1000 | − | + | + | >6 mo |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prkno, A.; Hoffmann, D.; Kaiser, M.; Goerigk, D.; Pfeffer, M.; Winter, K.; Vahlenkamp, T.W.; Beer, M.; Starke, A. Field Trial Vaccination against Cowpox in Two Alpaca Herds. Viruses 2020, 12, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020234
Prkno A, Hoffmann D, Kaiser M, Goerigk D, Pfeffer M, Winter K, Vahlenkamp TW, Beer M, Starke A. Field Trial Vaccination against Cowpox in Two Alpaca Herds. Viruses. 2020; 12(2):234. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020234
Chicago/Turabian StylePrkno, Almut, Donata Hoffmann, Matthias Kaiser, Daniela Goerigk, Martin Pfeffer, Karsten Winter, Thomas W. Vahlenkamp, Martin Beer, and Alexander Starke. 2020. "Field Trial Vaccination against Cowpox in Two Alpaca Herds" Viruses 12, no. 2: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/v12020234