Serological Survey of Lyssaviruses in Polish Bats in the Frame of Passive Rabies Surveillance Using an Enzyme-Linked Immunosorbent Assay

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
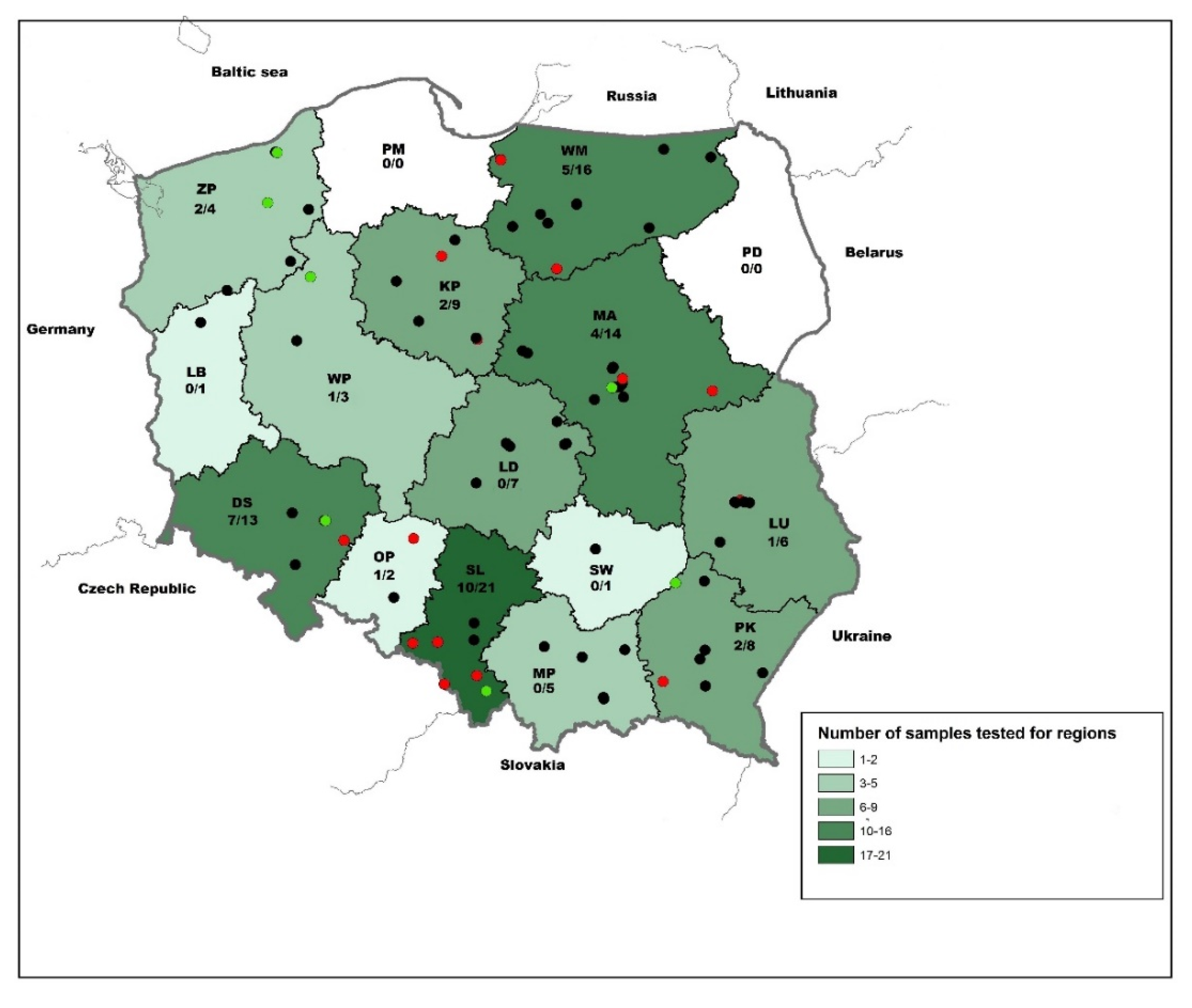
2.1. Samples
2.2. Molecular Classification of Polish Bats Based on Genetic Markers
2.3. Detection of Lyssavirus Antibodies with Bio-Pro Rabies ELISA Ab Kit
2.4. Modified Rapid Fluorescent Foci Inhibition Test (RFFIT)
3. Results
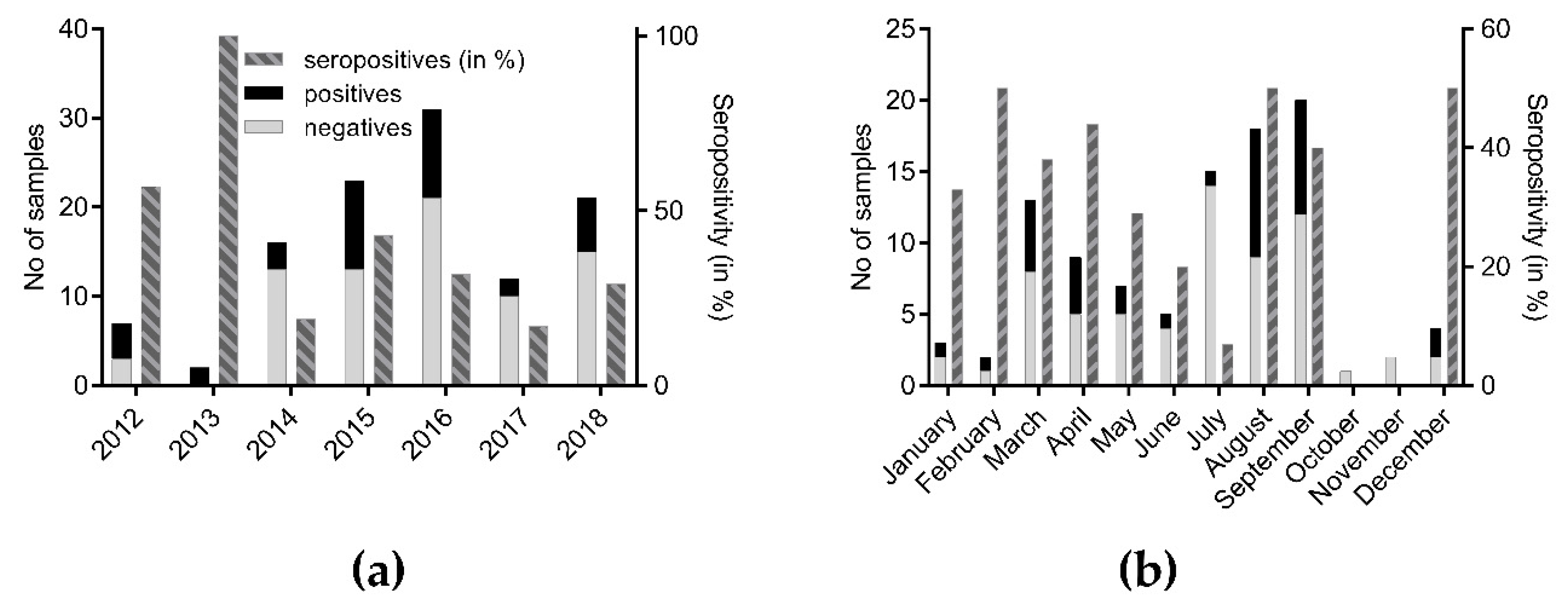
3.1. Serological Survey of Polish Bats
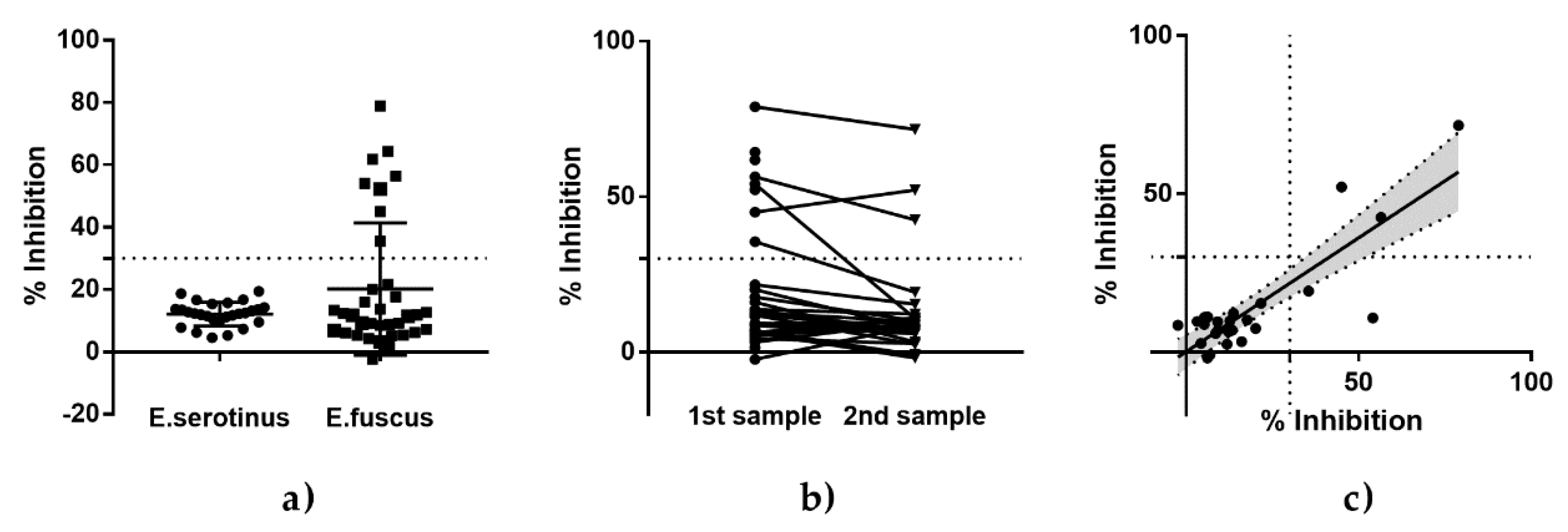
3.2. Serological Testing of Eptesicus Bats (Eptesicus Fuscus and Eptesicus Serotinus) from Experimental Studies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayman, D.T. Bats as Viral Reservoirs. Annu. Rev. Virol. 2016, 3, 77–99. [Google Scholar] [CrossRef]
- Fooks, A.R.; Cliquet, F.; Finke, S.; Freuling, C.; Hemachudha, T.; Mani, R.S.; Müller, T.; Nadin-Davis, S.; Picard-Meyer, E.; Wilde, H.; et al. Rabies. Nat. Rev. Dis. Primers. 2017, 3, 1–19. [Google Scholar] [CrossRef]
- Amarasinghe, G.K.; Arechiga Ceballos, N.G.; Banyard, A.C.; Basler, C.F.; Bavari, S.; Bennett, A.J.; Blasdell, K.R.; Briese, T.; Bukreyev, A.; Cai, Y.; et al. Taxonomy of the order Mononegavirales: Update 2018. Arch. Virol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Banyard, A.C.; Hayman, D.; Freuling, C.M.; Müller, T.; Fooks, A.R.; Johnson, N. Bat rabies. In Rabies: Scientific Basis of the Disease and Its Management, 3rd ed.; Jackson, A.C., Ed.; Academic Press: New York, NY, USA, 2013; Volume 3, pp. 215–267. [Google Scholar]
- Nokireki, T.; Tammiranta, N.; Kokkonen, U.-M.; Kantala, T.; Gadd, T. Tentative novel lyssavirus in a bat in Finland. Transbound. Emerg. Dis. 2018, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.C.; Hsu, C.L.; Lee, M.S.; Tu, Y.C.; Chang, J.C.; Wu, C.H.; Lee, S.H.; Ting, L.J.; Tsai, K.R.; Cheng, M.C.; et al. Lyssavirus in Japanese Pipistrelle, Taiwan. Emerg. Infect. Dis. 2018, 24, 782–785. [Google Scholar] [CrossRef] [Green Version]
- McElhinney, L.M.; Marston, D.A.; Wise, E.L.; Freuling, C.M.; Bourhy, H.; Zanoni, R.; Moldal, T.; Kooi, E.A.; Neubauer-Juric, A.; Nokireki, T.; et al. Molecular Epidemiology and Evolution of European Bat Lyssavirus 2. Int. J. Mol. Sci. 2018, 19, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smreczak, M.; Orłowska, A.; Marzec, A.; Trębas, P.; Müller, T.; Freuling, C.M.; Żmudziński, J.F. Bokeloh bat lyssavirus isolation in a Natterer’s bat, Poland. Zoonoses Public Health 2018, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Eggerbauer, E.; Troupin, C.; Passior, K.; Pfaff, F.; Höper, D.; Neubauer-Juric, A.; Haberl, S.; Bouchier, C.; Mettenleiter, T.C.; Bourhy, H.; et al. The Recently Discovered Bokeloh Bat Lyssavirus: Insights Into Its Genetic Heterogeneity and Spatial Distribution in Europe and the Population Genetics of Its Primary Host. Adv. Virus Res. 2017, 99, 199–232. [Google Scholar] [CrossRef] [PubMed]
- Banyard, A.C.; Selden, D.; Wu, G.; Thorne, L.; Jennings, D.; Marston, D.; Finke, S.; Freuling, C.M.; Müller, T.; Echevarria, J.E.; et al. Isolation, antigenicity and immunogenicity of Lleida bat lyssavirus. J. Gen. Virol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Beven, V.; Hirchaud, E.; Guillaume, C.; Larcher, G.; Robardet, E.; Servat, A.; Blanchard, Y.; Cliquet, F. Lleida Bat Lyssavirus isolation in Miniopterus schreibersii in France. Zoonoses Public Health 2018. [Google Scholar] [CrossRef] [PubMed]
- Arechiga Ceballos, N.; Moron, S.V.; Berciano, J.M.; Nicolas, O.; Lopez, C.A.; Juste, J.; Nevado, C.R.; Setien, A.A.; Echevarria, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botvinkin, A.D.; Poleschuk, E.M.; Kuzmin, I.; Borisova, T.I.; Gazaryan, S.V.; Yager, P.; Rupprecht, C.E. Novel lyssaviruses isolated from bats in Russia. Emerg. Infect. Dis. 2003, 9, 1623–1625. [Google Scholar] [CrossRef] [PubMed]
- Smreczak, M.; Trebas, P.; Orlowska, A.; Zmudzinski, J.F. Rabies surveillance in Poland (1992–2006). Dev. Biol. (Basel) 2008, 131, 249–256. [Google Scholar] [PubMed]
- WHO Rabies Bulletin Europe database. Available online: https://www.who-rabies-bulletin.org/ (accessed on 10 January 2019).
- Schatz, J.; Fooks, A.R.; McElhinney, L.; Horton, D.; Echevarria, J.; Vazquez-Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat Rabies Surveillance in Europe. Zoonoses Public Health 2013, 60, 22–34. [Google Scholar] [CrossRef]
- Med Vet Net Working Group. Passive and active surveillance of bat lyssavirus infections. Rabies Bull. Eur. 2005, 29, 5. [Google Scholar]
- Freuling, C.; Vos, A.; Johnson, N.; Kaipf, I.; Denzinger, A.; Neubert, L.; Mansfield, K.L.; Hicks, D.; Nunez, A.; Tordo, N.; et al. Experimental infection of serotine bats (Eptesicus serotinus) with European bat lyssavirus type 1a. J. Gen. Virol. 2009, 90, 2493–2502. [Google Scholar] [CrossRef]
- Franka, R.; Johnson, N.; Müller, T.; Vos, A.; Neubert, L.; Freuling, C.; Rupprecht, C.E.; Fooks, A.R. Susceptibility of North American big brown bats (Eptesicus fuscus) to infection with European bat lyssavirus type 1. J. Gen. Virol. 2008, 89, 1998–2010. [Google Scholar] [CrossRef]
- Johnson, N.; Vos, A.; Neubert, L.; Freuling, C.; Mansfield, K.L.; Kaipf, I.; Denzinger, A.; Hicks, D.; Nunez, A.; Franka, R.; et al. Experimental study of European bat lyssavirus type-2 infection in Daubenton’s bats (Myotis daubentonii). J. Gen. Virol. 2008, 89, 2662–2672. [Google Scholar] [CrossRef]
- Schatz, J.; Ohlendorf, B.; Busse, P.; Pelz, G.; Dolch, D.; Teubner, J.; Encarnacao, J.A.; Muhle, R.U.; Fischer, M.; Hoffmann, B.; et al. Twenty years of active bat rabies surveillance in Germany: A detailed analysis and future perspectives. Epidemiol. Infect. 2013, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Picard-Meyer, E.; Dubourg-Savage, M.-J.; Arthur, L.; Barataud, M.; Bécu, D.; Bracco, S.; Borel, C.; Larcher, G.; Meme-Lafond, B.; Moinet, M.; et al. Active surveillance of bat rabies in France: A 5-year study (2004–2009). Vet. Microbiol. 2011, 151, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Hammarin, A.-L.; Berndtsson, L.T.; Falk, K.; Nedinge, M.; Olsson, G.; Lundkvist, Å. Lyssavirus-reactive antibodies in Swedish bats. Infect. Ecol. Epidemiol. 2016, 6, 31262. [Google Scholar] [CrossRef]
- Simic, I.; Lojkic, I.; Kresic, N.; Cliquet, F.; Picard-Meyer, E.; Wasniewski, M.; Cukusic, A.; Zrncic, V.; Bedekovic, T. Molecular and serological survey of lyssaviruses in Croatian bat populations. BMC Vet. Res. 2018, 14, 274. [Google Scholar] [CrossRef]
- Robardet, E.; Borel, C.; Moinet, M.; Jouan, D.; Wasniewski, M.; Barrat, J.; Boue, F.; Montchatre-Leroy, E.; Servat, A.; Gimenez, O.; et al. Longitudinal survey of two serotine bat (Eptesicus serotinus) maternity colonies exposed to EBLV-1 (European Bat Lyssavirus type 1): Assessment of survival and serological status variations using capture-recapture models. PLOS Negl. Trop. Dis. 2017, 11, e0006048. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, S.; Ibanez, C.; Juste, J.; Echevarria, J.E. EBLV1 circulation in natural bat colonies of Eptesicus serotinus: A six year survey. Dev. Biol. (Basel) 2006, 125, 257–261. [Google Scholar]
- Colombi, D.; Serra-Cobo, J.; Métras, R.; Apolloni, A.; Poletto, C.; López-Roig, M.; Bourhy, H.; Colizza, V. Mechanisms for lyssavirus persistence in non-synanthropic bats in Europe: Insights from a modeling study. Sci. Rep. 2019, 9, 537. [Google Scholar] [CrossRef]
- Leopardi, S.; Priori, P.; Zecchin, B.; Poglayen, G.; Trevisiol, K.; Lelli, D.; Zoppi, S.; Scicluna, M.T.; D’Avino, N.; Schiavon, E.; et al. Active and passive surveillance for bat lyssaviruses in Italy revealed serological evidence for their circulation in three bat species. Epidemiol. Infect. 2018, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Megali, A.; Yannic, G.; Zahno, M.L.; Brugger, D.; Bertoni, G.; Christe, P.; Zanoni, R. Surveillance for European bat lyssavirus in Swiss bats. Arch. Virol. 2010, 155, 1655–1662. [Google Scholar] [CrossRef]
- Serra-Cobo, J.; Amengual, B.; Abellan, C.; Bourhy, H. European bat Lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef]
- Cliquet, F.; McElhinney, L.M.; Servat, A.; Boucher, J.M.; Lowings, J.P.; Goddard, T.; Mansfield, K.L.; Fooks, A.R. Development of a qualitative indirect ELISA for the measurement of rabies virus-specific antibodies from vaccinated dogs and cats. J. Virol. Methods 2004, 117, 1–8. [Google Scholar] [CrossRef]
- Servat, A.; Feyssaguet, M.; Blanchard, I.; Morize, J.L.; Schereffer, J.L.; Boue, F.; Cliquet, F. A quantitative indirect ELISA to monitor the effectiveness of rabies vaccination in domestic and wild carnivores. J. Immunol. Methods 2007, 318, 1–10. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, Y.; Zhang, F.; Hu, R. Competitive ELISA using a rabies glycoprotein-transformed cell line to semi-quantify rabies neutralizing-related antibodies in dogs. Vaccine 2009, 27, 2108–2113. [Google Scholar] [CrossRef] [PubMed]
- Wasniewski, M.; Guiot, A.L.; Schereffer, J.L.; Tribout, L.; Mähar, K.; Cliquet, F. Evaluation of an ELISA to detect rabies antibodies in orally vaccinated foxes and raccoon dogs sampled in the field. J. Virol. Methods 2013, 187, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.J.; Abelseth, M.K.; Athanasiu, P. The fluorescence antibody test In Laboratory techniques in rabies, 4th ed.; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 88–93. [Google Scholar]
- Heaton, P.R.; Johnstone, P.; McElhinney, L.M.; Cowley, R.; O’Sullivan, E.; Whitby, J.E. Heminested PCR assay for detection of six genotypes of rabies and rabies-related viruses. J. Clin. Microbiol. 1997, 35, 2762–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, S.L.; Johnson, N.; Brookes, S.M.; Hutson, A.M.; Fooks, A.R.; Jones, G. The application of genetic markers for EBLV surveillance in European bat species. Dev. Biol. (Basel) 2008, 131, 347–363. [Google Scholar] [PubMed]
- Wasniewski, M.; Cliquet, F. Evaluation of ELISA for detection of rabies antibodies in domestic carnivores. J. Virol. Methods 2012, 179, 166–175. [Google Scholar] [CrossRef]
- Wasniewski, M.; Almeida, I.; Baur, A.; Bedekovic, T.; Boncea, D.; Chaves, L.B.; David, D.; De Benedictis, P.; Dobrostana, M.; Giraud, P.; et al. First international collaborative study to evaluate rabies antibody detection method for use in monitoring the effectiveness of oral vaccination programmes in fox and raccoon dog in Europe. J. Virol. Methods 2016, 238, 77–85. [Google Scholar] [CrossRef]
- Tyem, D.; Dogonyaro, B.; Woma, T.; Ngoepe, E.; Sabeta, C. Sero-Surveillance of Lyssavirus Specific Antibodies in Nigerian Fruit Bats (Eidolon helvum). Trop. Med. Infect. Dis. 2017, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Malerczyk, C.; Selhorst, T.; Tordo, N.; Moore, S.A.; Müller, T. Antibodies induced by vaccination with purified chick embryo cell culture vaccine (PCECV) cross-neutralize non-classical bat lyssavirus strains. Vaccine 2009, 27, 5320–5325. [Google Scholar] [CrossRef]
- Calisher, C.H.; Child, J.E.; Field, H.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Anon. Agreement on the Conservation of Populations of Bats in Europe (EUROBATS), Annex 5: Bat rabies. Available online: http://www.eurobats.org/sites/default/files/documents/pdf/Meeting_of_Parties/MoP5_Record_Annex5_Res5_2_bat_rabies.pdf (accessed on 27 February 2019).
- George, D.B.; Webb, C.T.; Farnsworth, M.L.; O’Shea, T.J.; Bowen, R.A.; Smith, D.L.; Stanley, T.R.; Ellison, L.E.; Rupprecht, C.E. Host and viral ecology determine bat rabies seasonality and maintenance. Proc. Natl. Acad. Sci. USA 2011, 108, 10208–10213. [Google Scholar] [CrossRef] [Green Version]
- Serra-Cobo, J.; Lopez-Roig, M.; Segui, M.; Sanchez, L.P.; Nadal, J.; Borras, M.; Lavenir, R.; Bourhy, H. Ecological factors associated with European bat lyssavirus seroprevalence in spanish bats. PLoS ONE 2013, 8, e64467. [Google Scholar] [CrossRef]
- Picard-Meyer, E.; Robardet, E.; Arthur, L.; Larcher, G.; Harbusch, C.; Servat, A.; Cliquet, F. Bat Rabies in France: A 24-Year Retrospective Epidemiological Study. PLoS ONE 2014, 9, e98622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.; Gilbert, A.; Vos, A.; Freuling, C.M.; Ellis, C.; Kliemt, J.; Müller, T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Trop. Med. Infect. Dis. 2017, 2, 31. [Google Scholar] [CrossRef]
- Müller, T.; Selhorst, T.; Burow, J.; Schameitat, A.; Vos, A. Cross reactive antigenicity in orally vaccinated foxes and raccoon dogs against European Bat Lyssavirus type 1 and 2. Dev. Biol. (Basel) 2006, 125, 195–204. [Google Scholar] [PubMed]
- Colling, A.; Lunt, R.; Bergfeld, J.; McNabb, L.; Halpin, K.; Juzva, S.; Newberry, K.; Morrissy, C.; Loomes, C.; Warner, S.; et al. A network approach for provisional assay recognition of a Hendra virus antibody ELISA: Test validation with low sample numbers from infected horses. J. Vet. Diagn. Invest. 2018, 30, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Knoop, E.V.; Freuling, C.; Kliemt, J.; Selhorst, T.; Conraths, F.; Müller, T. Evaluation of a commercial rabies ELISA as a replacement for serum neutralization assays as part of the pet travel scheme and oral vaccination campaigns of foxes. Berl. Munch. Tierarztl. Wochenschr. 2010, 123, 278–285. [Google Scholar] [PubMed]
- Horton, D.L.; McElhinney, L.M.; Marston, D.A.; Wood, J.L.N.; Russell, C.A.; Lewis, N.; Kuzmin, I.V.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Fooks, A.R.; et al. Quantifying Antigenic Relationships among the Lyssaviruses. J. Virol. 2010, 84, 11841–11848. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | Females | Males | Total | |||
|---|---|---|---|---|---|---|
| Npos/Ntotal | Positives % (95% CI) | Npos/Ntotal | Positives % (95% CI) | Npos/Ntotal | Positives % (95% CI) | |
| Eptesicus serotinus | 2/7 | 28.57% (5.08–64.11) | 4/24 | 16.67% (6.68–35.85) | 6/31 | 19.35% (9.19–36.28) |
| Nyctalus noctula | 5/13 | 38.46% (17.71–64.48) | 2/13 | 15.38% (2.73–42.23) | 7/26 | 26.92%(13.7–46.08) |
| Plecotus auritus | 2/2 | 100% (17.77–100) | 3/4 | 75% (30.06–98.72) | 5/6 | 83.33% (43.65–99.15) |
| Hypsugo savii | 0 | 0/1 | 0% (0–94.81) | 0/1 | 0% (0–94.81) | |
| Myotis dasycneme | 0/1 | 0% (0–94.81) | 0 | 0/1 | 0% (0–94.81) | |
| Vespertillo murinus | 1/6 | 16.67% (0.85–56.35) | 0/3 | 0% (0–56.15) | 1/9 | 11.11% (0.57–43.5) |
| Pipistrellus pipistrellus | 1/1 | 100% (5.13–100) | 0/1 | 0% (0–94.81) | 1/2 | 50% (2.56–97.44) |
| P. pipistrellus/P. pygmaeus | 1/1 | 100% (5.13–100) | 0 | 1/1 | 100% (5.13–100) | |
| Myotis daubentonii | 1/2 | 50% (2.56–97.44) | 2/2 | 100% (17.77–100) | 3/4 | 75% (30.06–98.72) |
| Myotis brandtii | 1/1 | 100% (5.13–100) | 1/1 | 100% (5.13–100) | 2/2 | 2 (100) |
| Barbastella barbastellus | 1/1 | 100% (5.13–100) | 0 | 1/1 | 100% (5.13–100) | |
| Plecotus austriacus | 0/1 | 0% (0–94.81) | 0 | 0/1 | 0% (0–94.81) | |
| Myotis nattereri | 0/1 | 0% (0–94.81) | 0 | 0/1 | 0% (0–94.81) | |
| n/a | 5/10 | 50% (23.66–76.34) | 1/17 | 5.88% (0.3–26.98) | 6/27 | 22.22% (10.61–40.76) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orłowska, A.; Smreczak, M.; Freuling, C.M.; Müller, T.; Trębas, P.; Rola, J. Serological Survey of Lyssaviruses in Polish Bats in the Frame of Passive Rabies Surveillance Using an Enzyme-Linked Immunosorbent Assay. Viruses 2020, 12, 271. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030271
Orłowska A, Smreczak M, Freuling CM, Müller T, Trębas P, Rola J. Serological Survey of Lyssaviruses in Polish Bats in the Frame of Passive Rabies Surveillance Using an Enzyme-Linked Immunosorbent Assay. Viruses. 2020; 12(3):271. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030271
Chicago/Turabian StyleOrłowska, Anna, Marcin Smreczak, Conrad Martin Freuling, Thomas Müller, Paweł Trębas, and Jerzy Rola. 2020. "Serological Survey of Lyssaviruses in Polish Bats in the Frame of Passive Rabies Surveillance Using an Enzyme-Linked Immunosorbent Assay" Viruses 12, no. 3: 271. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030271