Cloning, Prokaryotic Soluble Expression, and Analysis of Antiviral Activity of Two Novel Feline IFN-ω Proteins

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids, Cells, and Viruses
2.2. Animal and Ethics Statement
2.3. Cloning of feIFN-ω Genes
2.4. Analysis of feIFN-ω Gene and Protein Characteristics
2.5. Soluble Expression of Recombinant feIFN-ω
2.6. Determination of the Antiviral Activity of feIFN-ω
2.7. Statistical Analysis
3. Results
3.1. Identification of the Novel Feline IFN-ω Genes
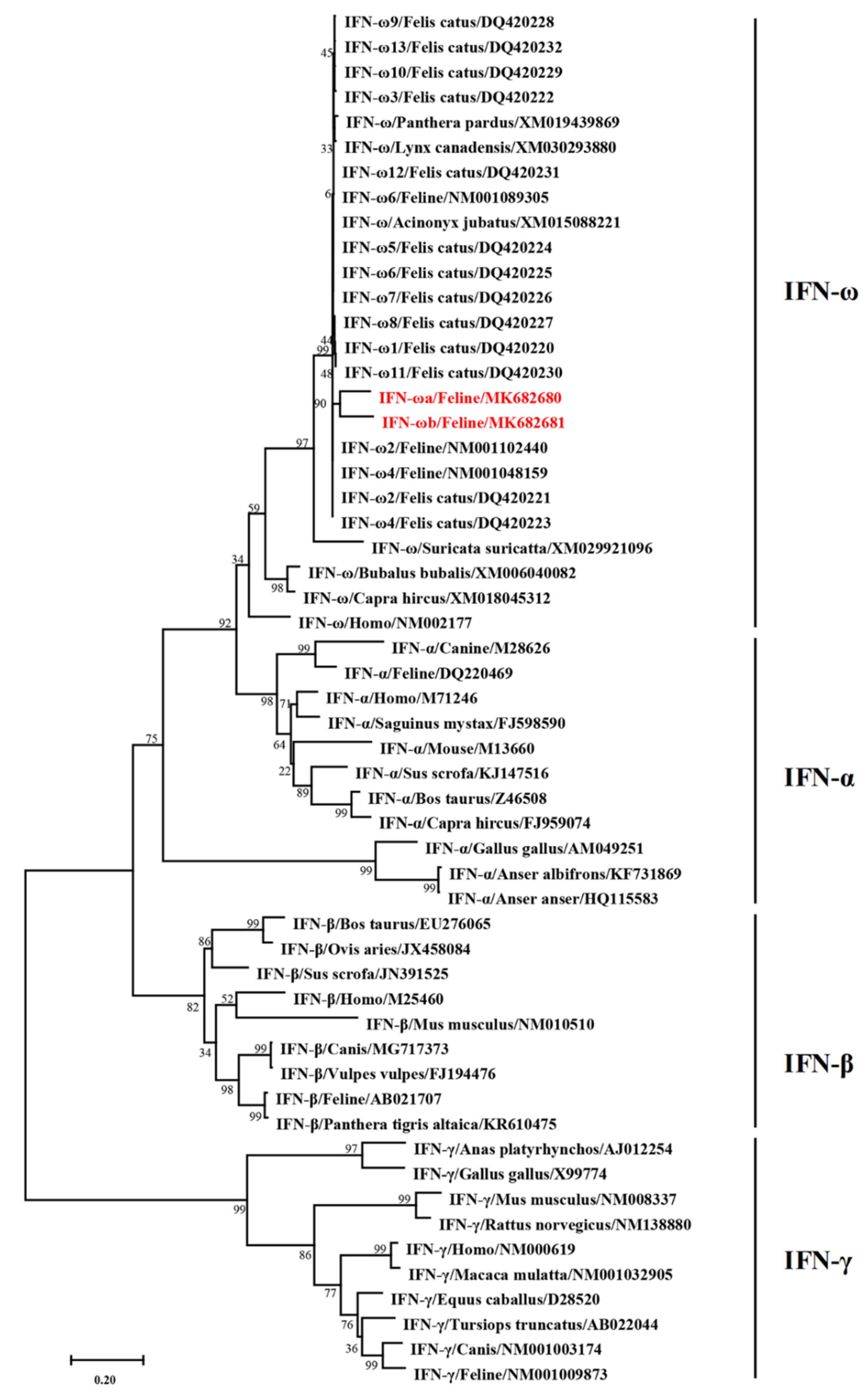
3.2. Phylogenetic Tree Analysis of the Newly Identified Feline IFN-ω Genes
3.3. Bioinformatics Analysis for the Novel Feline IFN-ω Proteins
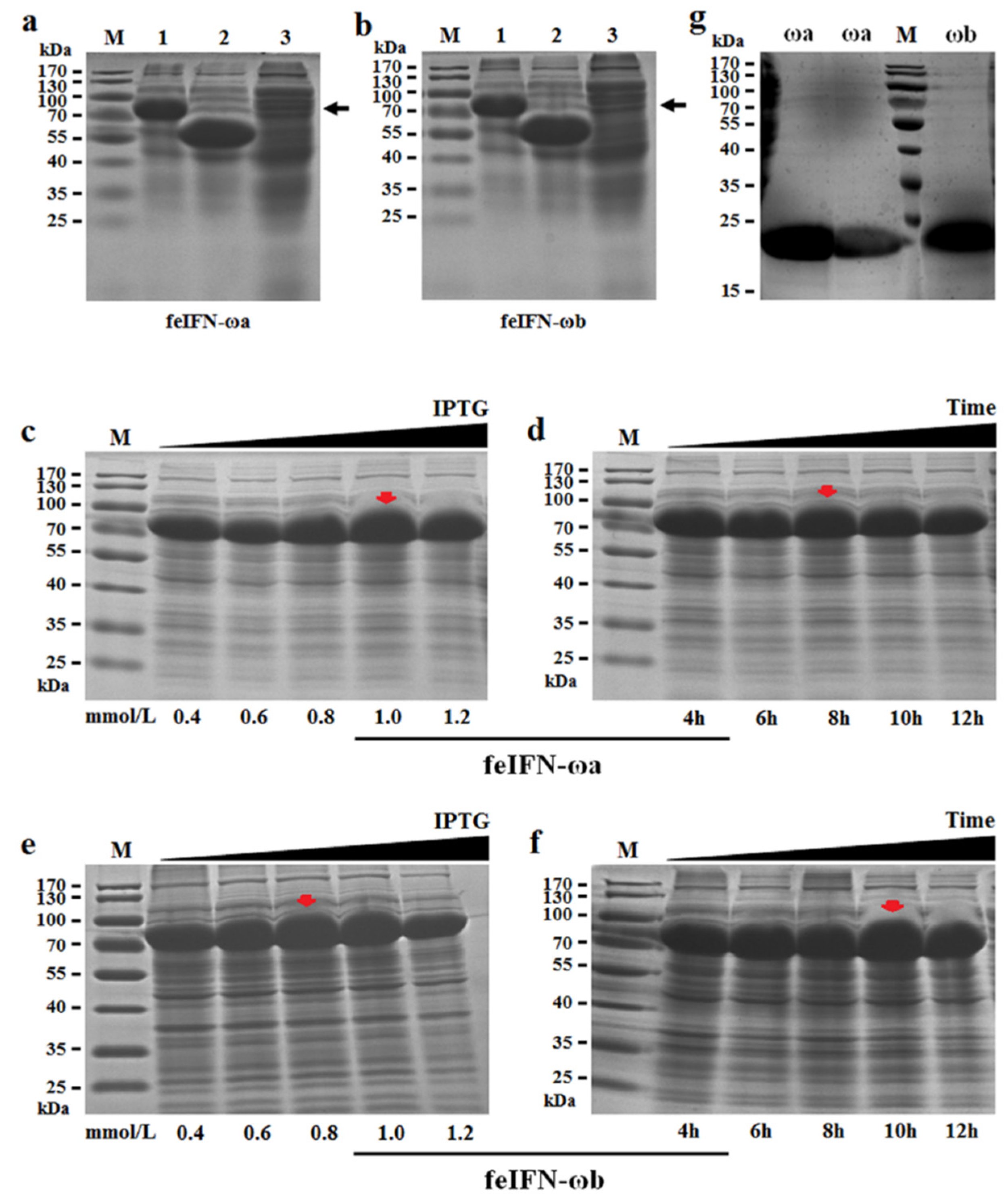
3.4. Soluble Expression and Purification of the Newly Identified feIFN-ω Proteins
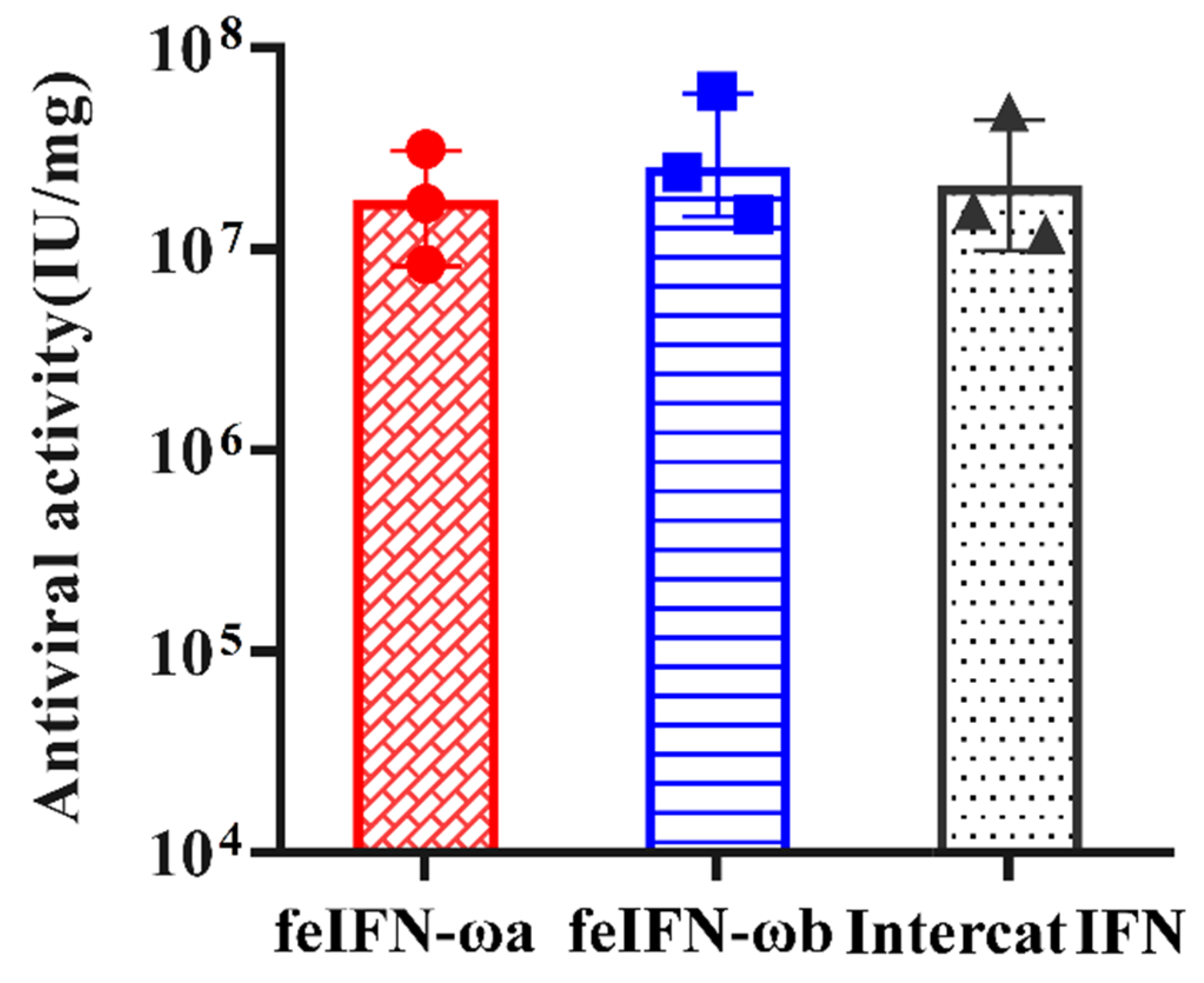
3.5. Determination of the Antiviral Activities of Recombinant feIFN-ωa and feIFN-ωb
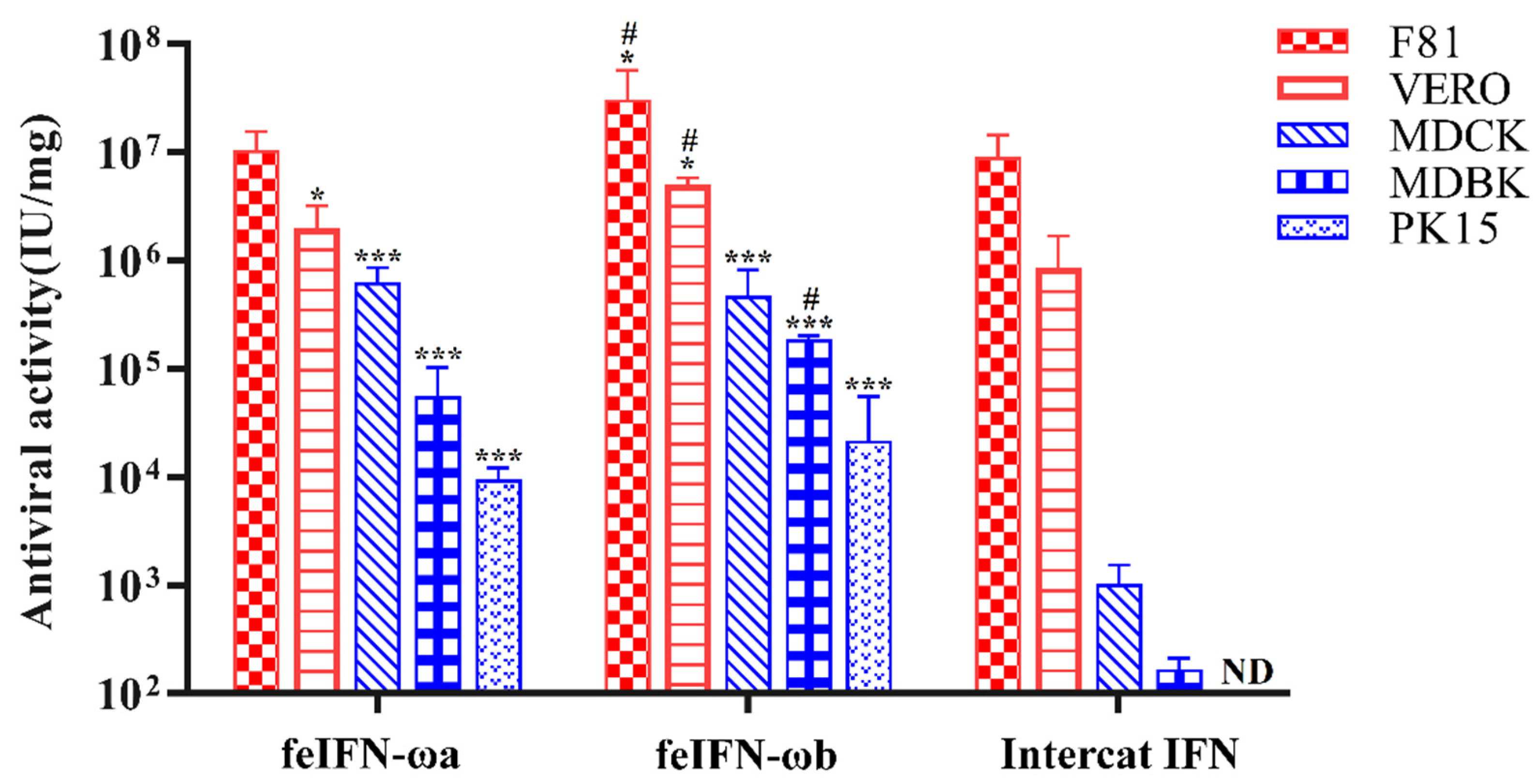
3.6. Species-Specific Antiviral Activities of Recombinant feIFN-ωa and feIFN-ωb
3.7. Broad-Spectrum Antiviral Activities of feIFN-ωa and feIFN-ωb
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Willeberg, P. Epidemiology of naturally occurring feline urologic syndrome. Vet. Clin. N. Am. Anim. Pract. 1984, 14, 455–469. [Google Scholar] [CrossRef]
- VandeWoude, S.; Hofmann-Lehmann, R.; Gonczi, E.; Riond, B.; Meli, M.; Willi, B.; Howard, J.; Schaarschmidt-Kiener, D.; Regli, W.; Gilli, U.; et al. Feline leukemia virus infection: Importance and current situation in Switzerland. J. Virol. 2018, 160, 95–105. [Google Scholar]
- Ritz, S.; Egberink, H.; Hartmann, K. Effect of feline interferon-omega on the survival time and quality of life of cats with feline infectious peritonitis. J. Vet. Intern. Med. 2007, 21, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Westman, M.E.; Malik, R.; Hall, E.; Harris, M.; Norris, J.M. The protective rate of the feline immunodeficiency virus vaccine: An Australian field study. Vaccine 2016, 34, 4752–4758. [Google Scholar] [CrossRef] [PubMed]
- Poncelet, L.; Garigliany, M.; Ando, K.; Franssen, M.; Desmecht, D.; Brion, J.P. Cell cycle S phase markers are expressed in cerebral neuron nuclei of cats infected by the feline panleukopenia virus. Cell Cycle 2016, 15, 3482–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.F.; Zhao, F.R.; Shao, J.J.; Xie, Y.L.; Chang, H.Y.; Zhang, Y.G. Interferon-omega: Current status in clinical applications. Int. Immunopharmacol. 2017, 52, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Zhao, P.; Ma, B.; Guo, G.; Sun, Y.; Xing, M. Cloning, expression and antiviral bioactivity of red-crowned crane interferon-α. Gene 2014, 544, 49–55. [Google Scholar] [CrossRef]
- Leal, R.O.; Gil, S. The use of recombinant feline interferon omega therapy as an immune-modulator in cats naturally infected with feline immunodeficiency virus: New perspectives. Vet. Sci. 2016, 3, 32. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.M.; Liu, L.; Alexenko, A. New and atypical families of type I interferons in mammals: Comparative functions, structures, and evolutionary relationships. Prog. Nucleic Acid Res. Mol. Biol. 1997, 56, 287. [Google Scholar]
- Kotenko, S.V.; Gallagher, G.; Baurin, V.V.; Lewis-Antes, A.; Donnelly, R.P. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat. Immunol. 2003, 4, 69–77. [Google Scholar] [CrossRef]
- Sheppard, P.; Kindsvogel, W.; Xu, W.; Henderson, K.; Schlutsmeyer, S.; Whitmore, T.E.; Kuestner, R.; Garrigues, U.; Birks, C.; Roraback, J.; et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nat. Immunol. 2003, 4, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.M.; Xue, Q.H.; Sun, L.; Zhu, Y.P.; Liu, W.J. Cloning and characterization of a novel feline IFN-omega. J. Interferon Cytokine Res. 2007, 27, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Karine de Mari, L.M.; Sanquer, A.; Lebreux, B.; Hyone-Myong, E. Therapeutic effects of recombinant feline interferon-co on feline leukemia virus (FeLV)-infected and FeLV/feline immunodeficiency virus (FIV)-coinfected symptomatic cats. J. Vet. Intern. Med. 2004, 17, 477–482. [Google Scholar]
- Powers, J.A.; Chiu, E.S.; Kraberger, S.J.; Roelke-Parker, M.; Lowery, I.; Erbeck, K.; Troyer, R.; Carver, S. Feline leukemia virus (FeLV) disease outcomes in a domestic cat breeding colony: Relationship to endogenous FeLV and other chronic viral infections. J. Virol. 2018, 92, e00649-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, A.; Kilbey, A.; Naseer, A.; Levy, L.S.; Ahmad, S.; Watts, C.; Mackay, N.; Cameron, E.; Wilson, S.; Neil, J.C. Barriers to infection of human cells by feline leukemia virus: Insights into resistance to zoonosis. J. Virol. 2017, 91, e02119-16. [Google Scholar] [CrossRef] [Green Version]
- Collado, V.M.; Gómez-Lucía, E.; Tejerizo, G.; Miró, G.; Escolar, E.; Martín, S.; Doménech, A. Effect of type I interferons on the expression of feline leukaemia virus. Vet. Microbiol. 2007, 123, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Domenech, A.; Miro, G.; Collado, V.M.; Ballesteros, N.; Sanjose, L.; Escolar, E.; Martin, S.; Gomez-Lucia, E. Use of recombinant interferon omega in feline retrovirosis: From theory to practice. Vet. Immunol. Immunopathol. 2011, 143, 301–306. [Google Scholar] [CrossRef]
- Li, S.F.; Zhao, F.R.; Gong, M.J.; Shao, J.J.; Xie, Y.L.; Chang, H.Y.; Zhang, Y.G. Antiviral activity of porcine interferon omega 7 against foot-and-mouth disease virus In Vitro. J. Med. Virol. 2019, 91, 208–214. [Google Scholar] [CrossRef] [Green Version]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, N.C. An update on feline infectious peritonitis: Diagnostics and therapeutics. Vet. J. 2014, 201, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Tekes, G.; Thiel, H.J. Feline coronaviruses: Pathogenesis of feline infectious peritonitis. Adv. Virus Res. 2016, 96, 193–218. [Google Scholar] [PubMed]
- Husebye, E.S.; Perheentupa, J.; Rautemaa, R.; Kampe, O. Clinical manifestations and management of patients with autoimmune polyendocrine syndrome type I. J. Intern. Med. 2009, 265, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Leal, R.O.; Gil, S.; Brito, M.T.; McGahie, D.; Maria, M.M.; Tavares, L. The use of oral recombinant feline interferon omega in two cats with type II diabetes mellitus and concurrent feline chronic gingivostomatitis syndrome. Ir. Vet. J. 2013, 66, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampel, V.; Schwarz, B.; Kempf, C.; Köstlin, R.; Schillinger, U.; Küchenhoff, H.; Fenske, N.; Brill, T.; Hirschberger, J. Adjuvant immunotherapy of feline fibrosarcoma with recombinant feline interferon-omega. J. Vet. Intern. Med. 2007, 21, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Litzlbauer, P.; Weber, K.; Mueller, R.S. Oral and subcutaneous therapy of canine atopic dermatitis with recombinant feline interferon omega. Cytokine 2014, 66, 54–59. [Google Scholar] [CrossRef]
- Fan, W.; Xu, L.; Ren, L.; Qu, H.; Li, J.; Liang, J.; Liu, W.; Yang, L.; Luo, T. Functional characterization of canine interferon-lambda. J. Interferon Cytokine Res. 2014, 34, 848–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhao, B.; Wang, F.; Wang, M.; Xie, S.; Wang, S.; Han, C.; Zhu, L. Yak interferon-alpha loaded solid lipid nanoparticles for controlled release. Res. Vet. Sci. 2010, 88, 148–153. [Google Scholar] [CrossRef]
- Cai, D.; Wang, H.; He, P.; Zhu, C.; Wang, Q.; Wei, X.; Nomura, C.T.; Chen, S. A novel strategy to improve protein secretion via overexpression of the SppA signal peptide peptidase in Bacillus licheniformis. Microb. Cell Fact. 2017, 16, 70. [Google Scholar] [CrossRef]
- Lu, X.; Zhong, Q.; Liu, J.; Yang, F.; Lu, C.; Xiong, H.; Li, S.; Zhu, Y.; Wu, L. Efficient expression of chondroitinase ABC I for specific disaccharides detection of chondroitin sulfate. Int. J. Biol. Macromol. 2019, 143, 41–48. [Google Scholar] [CrossRef]
- Buckwold, V.E.; Wei, J.; Huang, Z.; Huang, C.; Nalca, A.; Wells, J.; Russell, J.; Collins, B.; Ptak, R.; Lang, W.; et al. Antiviral activity of CHO-SS cell-derived human omega interferon and other human interferons against HCV RNA replicons and related viruses. Antivir. Res. 2007, 73, 118–125. [Google Scholar] [CrossRef]
- Buckwold, V.E.; Wei, J.; Wenzel-Mathers, M.; Russell, J. Synergistic In Vitro interactions between alpha interferon and ribavirin against bovine viral diarrhea virus and yellow fever virus as surrogate models of hepatitis C virus replication. Antimicrob. Agents Chemother. 2003, 47, 2293–2298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Ueda, Y.; Sato, M.; Yanai, A. Feline interferon production in silkworm by recombinant baculovirus. J. Vet. Med. Sci. 1992, 54, 563–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiwata, K.; Minagawa, T.; Kajimoto, T. Clinical effects of the recombinant feline interferon-omega on experimental parvovirus infection in beagle dogs. J. Vet. Med. Sci. 1998, 60, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schijns, V.E.C.J.; Wierda, C.M.H.; Vahlenkamp, T.W.; Glansbeek, H.; Heijnen, I.; Horzinek, M.C. Feline IFN-γ produced in Spodoptera frugiperda cells by recombinant baculovirus exerts antiviral and immunoregulatory activity. Recent Res. Dev. Immunol. 2000, 2, 265–272. [Google Scholar]
- Argyle, D.J.; Harris, M.; Lawrence, C.; Mcbride, K.; Onions, D.E. Expression of feline recombinant interferon-γ in baculovirus and demonstration of biological activity. Vet. Immunol. Immunopathol. 1998, 64, 97–105. [Google Scholar] [CrossRef]
- Schijns, V.E.J.C.; Scholtes, N.C.; Van Zuilekom, H.I.; Sanders, L.E.H.H.; Nicolson, L.; Argyle, D.J. Facilitation of antibody forming responses to viral vaccine antigens in young cats by recombinant baculovirus-expressed feline IFN. Vaccine 2002, 20, 1718–1724. [Google Scholar] [CrossRef]
- Beck, N.B.; Sidhu, J.S.; Omiecinski, C.J. Baculovirus vectors repress phenobarbital-mediated gene induction and stimulate cytokine expression in primary cultures of rat hepatocytes. Gene Ther. 2000, 7, 1274–1283. [Google Scholar] [CrossRef]
- Sekar, N.; Veetil, S.K.; Neerathilingam, M. Tender coconut water an economical growth medium for the production of recombinant proteins in Escherichia coli. BMC Biotechnol. 2013, 13, 70. [Google Scholar] [CrossRef] [Green Version]
- Terpe, K. Overview of bacterial expression systems for heterologous protein production: From molecular and biochemical fundamentals to commercial systems. Appl. Microbiol. Biotechnol. 2006, 72, 211–222. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, M.; Zhang, L.; Zhang, T.; Ding, J.; Zhuang, Y.; Wang, X.; Yang, Z. An automatic refolding apparatus for preparative-scale protein production. PLoS ONE 2012, 7, e45891. [Google Scholar] [CrossRef] [Green Version]
- Wonderling, R.; Powell, T.; Baldwin, S.; Morales, T.; Snyder, S.; Keiser, K.; Hunter, S.; Best, E.; Mcdermott, M.J.; Milhausen, M. Cloning, expression, purification, and biological activity of five feline type I interferons. Vet. Immunol. Immunopathol. 2002, 89, 13–27. [Google Scholar] [CrossRef]
- He, X.; Korytar, T.; Schatz, J.; Freuling, C.M.; Muller, T.; Kollner, B. Anti-lyssaviral activity of interferons kappa and omega from the serotine bat, Eptesicus serotinus. J. Virol. 2014, 88, 5444–5454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, E.; Thorat, L.; Bhavnani, V.; Bendre, A.D.; Pal, J.K.; Nathb, B.B.; Gaikwad, S.M. Molecular cloning and in silico studies of physiologically significant trehalase from Drosophila melanogaster. Int. J. Biol. Macromol. 2016, 92, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Saini, P.; Wani, S.I.; Kumar, R.; Chhabra, R.; Chimni, S.S.; Sareen, D. Trigger factor assisted folding of the recombinant epoxide hydrolases identified from C. pelagibacter and S. nassauensis. Protein Exp. Purif. 2014, 104, 71–84. [Google Scholar] [CrossRef]
- Ballin, A.C.; Schulz, B.; Helps, C.; Sauter-Louis, C.; Mueller, R.S.; Hartmann, K. Limited efficacy of topical recombinant feline interferon-omega for treatment of cats with acute upper respiratory viral disease. Vet. J. 2014, 202, 466–470. [Google Scholar] [CrossRef]
- Cross, J.C.; Roberts, R.M. Constitutive and trophoblast-specific expression of a class of bovine interferon genes. Proc. Natl. Acad. Sci. USA 1991, 88, 3817–3821. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Guo, Y.; Bao, J.; Liu, Y.; An, D.; Ma, B.; Gao, M.; Wang, J. Characterization and antivirus activities of a novel bovine IFN-omega24. Mol. Immunol. 2015, 66, 357–363. [Google Scholar] [CrossRef]
- Buckwold, V.E.; Lang, W.; Scribner, C.; Blanchett, D.; Alessi, T.; Langecker, P. Safety pharmacology, toxicology and pharmacokinetic assessment of recombinant human omega-interferon produced from CHO-SS cells. Basic Clin. Pharmacol. Toxicol. 2006, 99, 62–70. [Google Scholar] [CrossRef]
- Tiefenthaler, M.; Geisen, F.; Schirmer, M.; Konwalinka, G. A comparison of the antiproliferative properties of recombinant human IFN-alpha 2 and IFN-omega in human bone marrow culture. J. Interferon Cytokine Res. 1997, 17, 327–329. [Google Scholar] [CrossRef]
- Penzo, C.; Ross, M.; Muirhead, R.; Else, R.; Argyle, D.J. Effect of recombinant feline interferon-ω alone and in combination with chemotherapeutic agents on putative tumour-initiating cells and daughter cells derived from canine and feline mammary tumours. Vet. Comp. Oncol. 2009, 7, 222–229. [Google Scholar] [CrossRef]
- Chen, P.F.; Fu, G.F.; Zhang, H.Y.; Xu, G.X.; Hou, Y.Y. Liposomal plasmid dna encoding human thymosin α1and interferon ω1 potently inhibits liver tumor growth in ICR mice. J. Gastroenterol. Hepatol. 2006, 21, 1538–1543. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IFN | Signal Peptide Cleavage Sites a | No. of Phosphorylation Sites b | No. of Glycosylation Sites c | Subcellular Localization d | Transmembrane Region e |
|---|---|---|---|---|---|
| IFN-ωa | Gly23-Cys24 | 15 | N-glycosylation sites (0) O-glycosylation sites (9) | 82.1% extracellular 7.2% intracellular 9.6% mitochondrion | Intracellular |
| IFN-ωb | Gly23-Cys24 | 13 | N-glycosylation sites (0) O-glycosylation sites (6) | 84.3% extracellular 9.2% intracellular 7% mitochondrion | Intracellular |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Li, F.; Han, M.; Jia, S.; Wang, L.; Qiao, X.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Cloning, Prokaryotic Soluble Expression, and Analysis of Antiviral Activity of Two Novel Feline IFN-ω Proteins. Viruses 2020, 12, 335. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030335
Wang X, Li F, Han M, Jia S, Wang L, Qiao X, Jiang Y, Cui W, Tang L, Li Y, et al. Cloning, Prokaryotic Soluble Expression, and Analysis of Antiviral Activity of Two Novel Feline IFN-ω Proteins. Viruses. 2020; 12(3):335. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030335
Chicago/Turabian StyleWang, Xiaona, Fengsai Li, Meijing Han, Shuo Jia, Li Wang, Xinyuan Qiao, Yanping Jiang, Wen Cui, Lijie Tang, Yijing Li, and et al. 2020. "Cloning, Prokaryotic Soluble Expression, and Analysis of Antiviral Activity of Two Novel Feline IFN-ω Proteins" Viruses 12, no. 3: 335. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030335