Coinfections of Novel Polyomavirus, Anelloviruses and a Recombinant Strain of Myxoma Virus-MYXV-Tol Identified in Iberian Hares

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Viral Nucleic Acid Extraction
2.2. Detection of the MYXV-Tol
2.3. Detection and Recovery of Anelloviruses and Polyomavirus Genomes
2.4. Sequence Analysis of MYXV-Tol Regions, Anelloviruses and Polyomaviruses
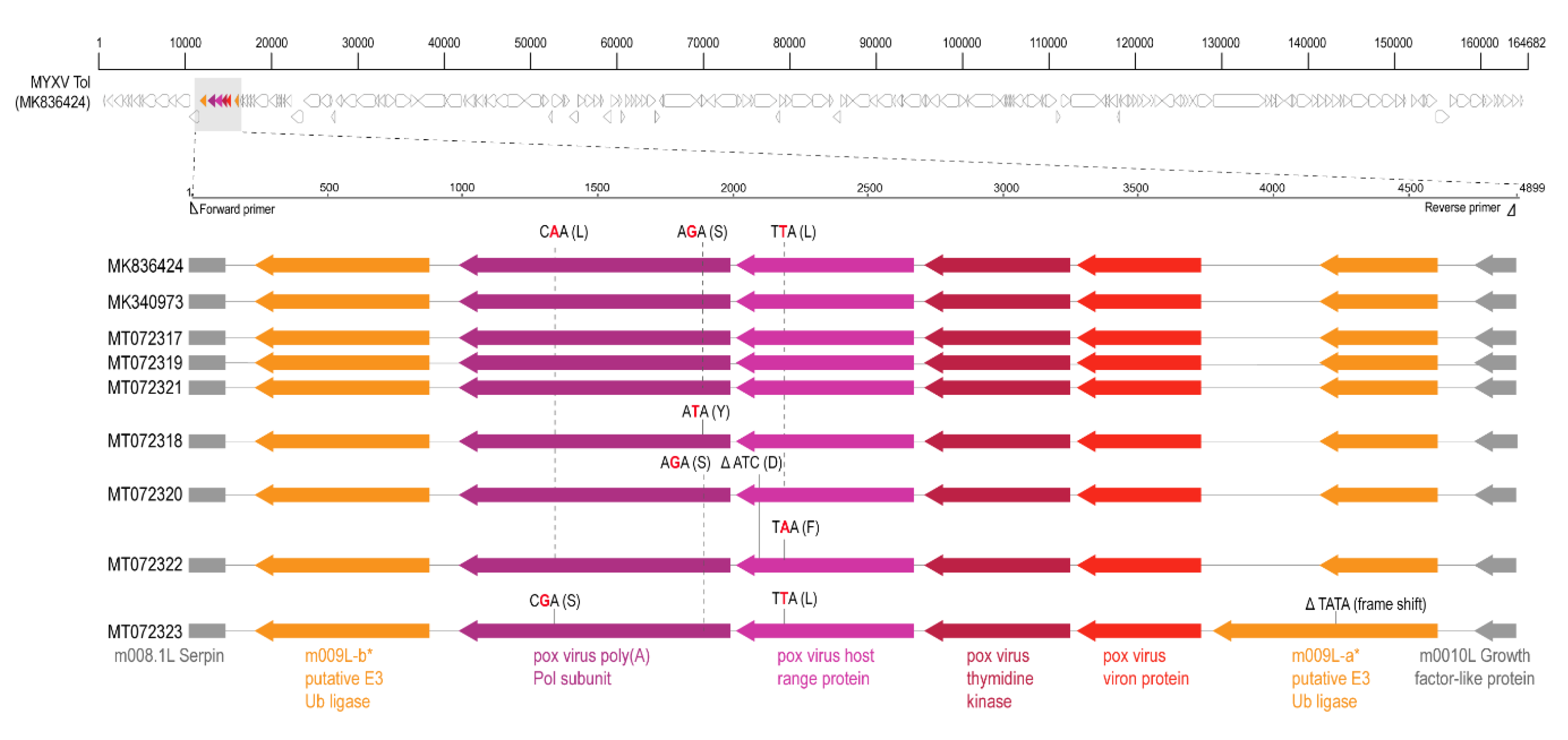
2.4.1. MYXV-Tol Recombinant Region Sequences
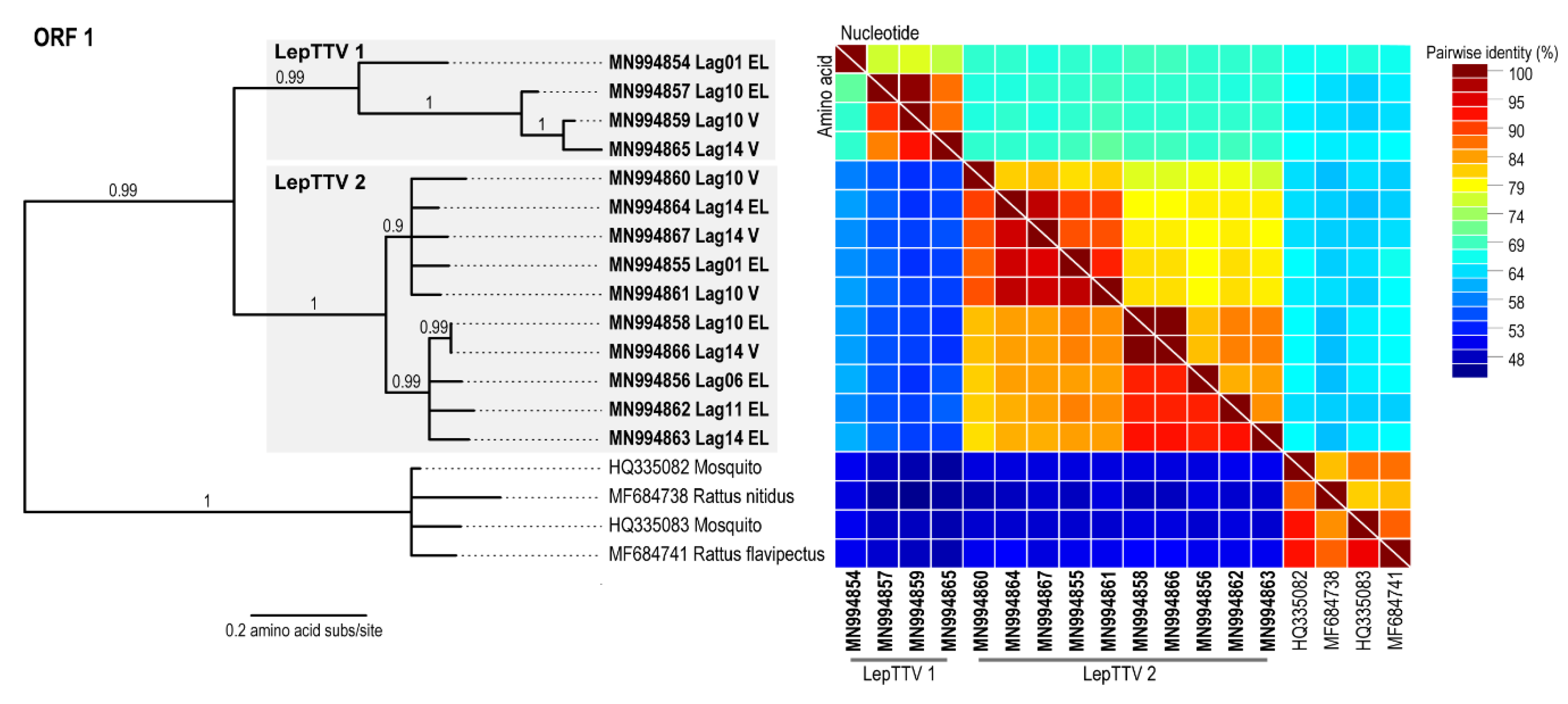
2.4.2. Anellovirus Sequences Analyses
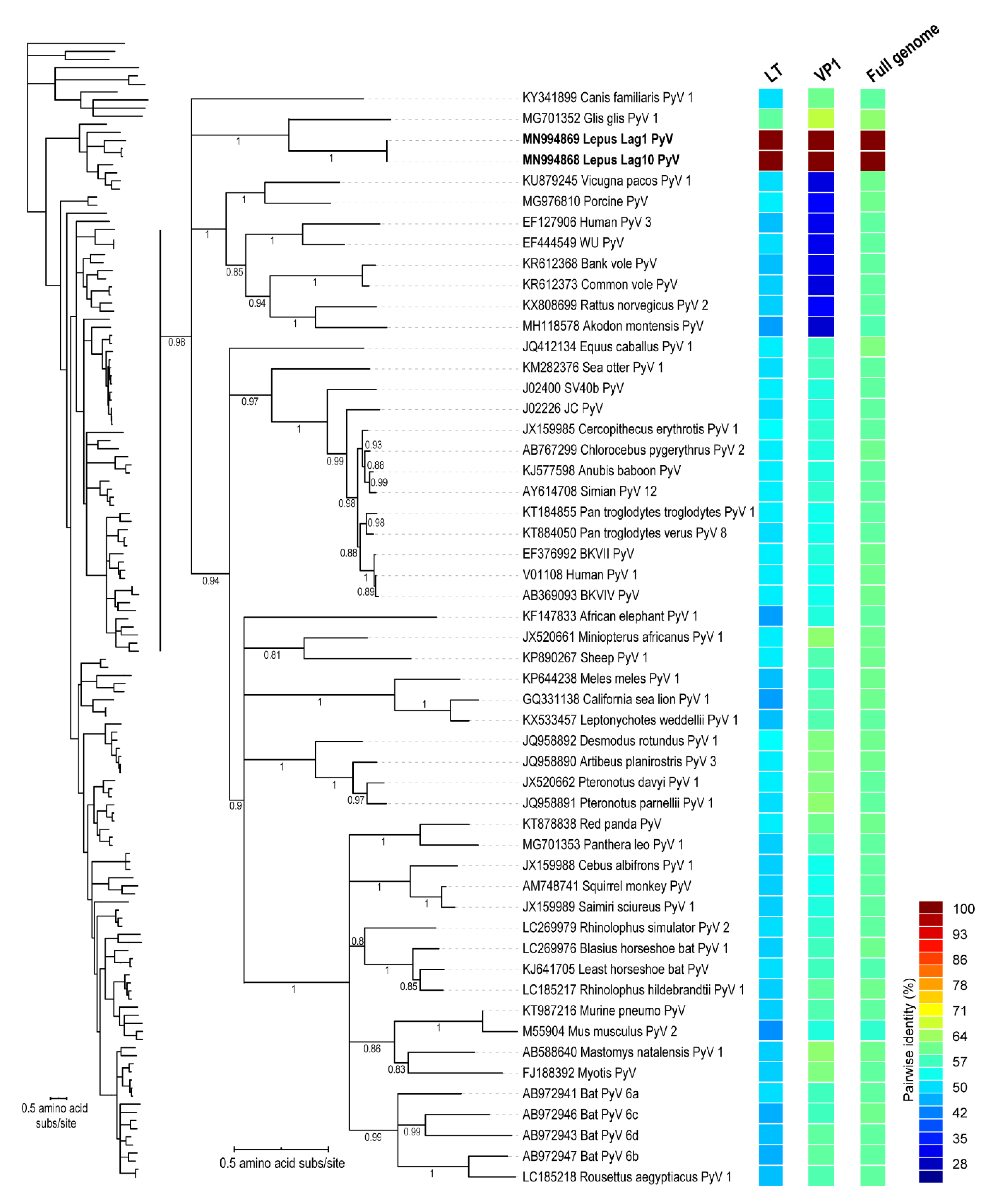
2.4.3. Polyomavirus Sequence Analyses
3. Results and Discussion
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Le Gall-Recule, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guerin, J.L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gall-Recule, G.; Zwingelstein, F.; Fages, M.P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouck, A.; Botti, G.; Lavazza, A.; Marchandeau, S. Characterisation of a non-pathogenic and non-protective infectious rabbit lagovirus related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strive, T.; Wright, J.D.; Robinson, A.J. Identification and partial characterisation of a new Lagovirus in Australian wild rabbits. Virology 2009, 384, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanford, M.M.; Werden, S.J.; McFadden, G. Myxoma virus in the European rabbit: Interactions between the virus and its susceptible host. Vet. Res. 2007, 38, 299–318. [Google Scholar] [CrossRef] [Green Version]
- Nash, P.; Barrett, J.; Cao, J.X.; Hota-Mitchell, S.; Lalani, A.S.; Everett, H.; Xu, X.M.; Robichaud, J.; Hnatiuk, S.; Ainslie, C.; et al. Immunomodulation by viruses: The myxoma virus story. Immunol. Rev. 1999, 168, 103–120. [Google Scholar] [CrossRef]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of myxoma virus on two continents. PLoS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef] [Green Version]
- Tsokana, C.N.; Sokos, C.; Giannakopoulos, A.; Birtsas, P.; Valiakos, G.; Spyrou, V.; Athanasiou, L.V.; Burriel, A.R.; Billinis, C. European Brown hare (Lepus europaeus) as a source of emerging and re-emerging pathogens of Public Health importance: A review. Vet. Med. Sci. 2020. [Google Scholar] [CrossRef]
- Agueda-Pinto, A.; Lemos de Matos, A.; Abrantes, M.; Kraberger, S.; Risalde, M.A.; Gortazar, C.; McFadden, G.; Varsani, A.; Esteves, P.J. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses 2019, 11, 530. [Google Scholar] [CrossRef] [Green Version]
- Dalton, K.P.; Martin, J.M.; Nicieza, I.; Podadera, A.; de Llano, D.; Casais, R.; Gimenez, S.; Badiola, I.; Aguero, M.; Duran, M.; et al. Myxoma virus jumps species to the Iberian hare. Transbound. Emerg. Dis. 2019, 66, 2218–2226. [Google Scholar] [CrossRef]
- Garcia-Bocanegra, I.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.P.; Caballero-Gomez, J.; Aguero, M.; Zorrilla, I.; Gomez-Guillamon, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 66, 2204–2208. [Google Scholar] [CrossRef]
- Abade dos Santos, F.A.; Monteiro, M.; Pinto, A.; Carvalho, C.L.; Peleteiro, M.C.; Carvalho, P.; Mendonça, P.; Carvalho, T.; Duarte, M.D. First description of a herpesvirus infection in genus Lepus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Steel, O.; Kraberger, S.; Sikorski, A.; Young, L.M.; Catchpole, R.J.; Stevens, A.J.; Ladley, J.J.; Coray, D.S.; Stainton, D.; Dayaram, A.; et al. Circular replication-associated protein encoding DNA viruses identified in the faecal matter of various animals in New Zealand. Infect. Genet. Evol. 2016, 43, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Kraberger, S.; Waits, K.; Ivan, J.; Newkirk, E.; VandeWoude, S.; Varsani, A. Identification of circular single-stranded DNA viruses in faecal samples of Canada lynx (Lynx canadensis), moose (Alces alces) and snowshoe hare (Lepus americanus) inhabiting the Colorado San Juan Mountains. Infect. Genet. Evol. 2018, 64, 1–8. [Google Scholar] [CrossRef]
- Nishiyama, S.; Dutia, B.M.; Stewart, J.P.; Meredith, A.L.; Shaw, D.J.; Simmonds, P.; Sharp, C.P. Identification of novel anelloviruses with broad diversity in UK rodents. J. Gen. Virol. 2014, 95, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Spandole, S.; Cimponeriu, D.; Berca, L.M.; Mihaescu, G. Human anelloviruses: An update of molecular, epidemiological and clinical aspects. Arch. Virol. 2015, 160, 893–908. [Google Scholar] [CrossRef] [PubMed]
- Hrazdilova, K.; Slaninkova, E.; Brozova, K.; Modry, D.; Vodicka, R.; Celer, V. New species of Torque Teno miniviruses infecting gorillas and chimpanzees. Virology 2016, 487, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Fahsbender, E.; Burns, J.M.; Kim, S.; Kraberger, S.; Frankfurter, G.; Eilers, A.A.; Shero, M.R.; Beltran, R.; Kirkham, A.; McCorkell, R.; et al. Diverse and highly recombinant anelloviruses associated with Weddell seals in Antarctica. Virus Evol. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Crane, A.; Goebel, M.E.; Kraberger, S.; Stone, A.C.; Varsani, A. Novel anelloviruses identified in buccal swabs of Antarctic fur seals. Virus Genes 2018, 54, 719–723. [Google Scholar] [CrossRef]
- Bigarre, L.; Beven, V.; de Boisseson, C.; Grasland, B.; Rose, N.; Biagini, P.; Jestin, A. Pig anelloviruses are highly prevalent in swine herds in France. J. Gen. Virol. 2005, 86, 631–635. [Google Scholar] [CrossRef]
- Cibulski, S.P.; Teixeira, T.F.; de Sales Lima, F.E.; do Santos, H.F.; Franco, A.C.; Roehe, P.M. A Novel Anelloviridae Species Detected in Tadarida brasiliensis Bats: First Sequence of a Chiropteran Anellovirus. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Giannitti, F.; Low, J.; Keyes, C.; Ullmann, L.S.; Deng, X.; Aleman, M.; Pesavento, P.A.; Pusterla, N.; Delwart, E. Exploring the virome of diseased horses. J. Gen. Virol. 2015, 96, 2721–2733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Doorslaer, K.; Kraberger, S.; Austin, C.; Farkas, K.; Bergeman, M.; Paunil, E.; Davison, W.; Varsani, A. Fish polyomaviruses belong to two distinct evolutionary lineages. J. Gen. Virol. 2018, 99, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Buck, C.B.; Van Doorslaer, K.; Peretti, A.; Geoghegan, E.M.; Tisza, M.J.; An, P.; Katz, J.P.; Pipas, J.M.; McBride, A.A.; Camus, A.C.; et al. The Ancient Evolutionary History of Polyomaviruses. PLoS Pathog. 2016, 12, e1005574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvignac-Spencer, S.; Feltkamp, M.C.; Daugherty, M.D.; Moens, U.; Ramqvist, T.; Johne, R.; Ehlers, B.; Polyomaviridae Study Group of the International Committee on Taxonomy of Viruses. A taxonomy update for the family Polyomaviridae. Arch. Virol. 2016, 161, 1739–1750. [Google Scholar] [CrossRef] [Green Version]
- Di Pietro, F.; Ortenzi, F.; Tilio, M.; Concetti, F.; Napolioni, V. Genomic DNA extraction from whole blood stored from 15- to 30-years at -20 degrees C by rapid phenol-chloroform protocol: A useful tool for genetic epidemiology studies. Mol. Cell. Probes 2011, 25, 44–48. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stover, B.C.; Muller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagini, P. Classification of TTV and related viruses (anelloviruses). Curr. Top. Microbiol. Immunol. 2009, 331, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Li, Y.; Lu, L.; Zheng, D.; Liu, B.; Yang, L.; Su, H.; Dong, J.; Sun, L.; Zhu, Y.; et al. Biodiversity of rodent anelloviruses in China. Emerg. Microbes Infect. 2018, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.F.; Willner, D.L.; Lim, Y.W.; Schmieder, R.; Chau, B.; Nilsson, C.; Anthony, S.; Ruan, Y.; Rohwer, F.; Breitbart, M. Broad surveys of DNA viral diversity obtained through viral metagenomics of mosquitoes. PLoS ONE 2011, 6, e20579. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, H.H.; Steiger, J. Polyomavirus BK. Lancet Infect. Dis. 2003, 3, 611–623. [Google Scholar] [CrossRef]
- Dalianis, T.; Hirsch, H.H. Human polyomaviruses in disease and cancer. Virology 2013, 437, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, B.; Anoh, A.E.; Ben Salem, N.; Broll, S.; Couacy-Hymann, E.; Fischer, D.; Gedvilaite, A.; Ingenhutt, N.; Liebmann, S.; Martin, M.; et al. Novel Polyomaviruses in Mammals from Multiple Orders and Reassessment of Polyomavirus Evolution and Taxonomy. Viruses 2019, 11, 930. [Google Scholar] [CrossRef] [Green Version]
- Moens, U.; Calvignac-Spencer, S.; Lauber, C.; Ramqvist, T.; Feltkamp, M.C.W.; Daugherty, M.D.; Verschoor, E.J.; Ehlers, B.; Ictv Report Consortium. ICTV Virus Taxonomy Profile: Polyomaviridae. J. Gen. Virol. 2017, 98, 1159–1160. [Google Scholar] [CrossRef]
- Freer, G.; Maggi, F.; Pifferi, M.; Di Cicco, M.E.; Peroni, D.G.; Pistello, M. The Virome and Its Major Component, Anellovirus, a Convoluted System Molding Human Immune Defenses and Possibly Affecting the Development of Asthma and Respiratory Diseases in Childhood. Front. Microbiol. 2018, 9, 686. [Google Scholar] [CrossRef] [Green Version]
- McElvania TeKippe, E.; Wylie, K.M.; Deych, E.; Sodergren, E.; Weinstock, G.; Storch, G.A. Increased prevalence of anellovirus in pediatric patients with fever. PLoS ONE 2012, 7, e50937. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Bendinelli, M. Immunobiology of the Torque Teno Viruses and Other Anelloviruses; TT Viruses; Springer: Berlin/Heidelberg, Germany, 2009; pp. 65–90. [Google Scholar]
- Zheng, H.; Ye, L.; Fang, X.; Li, B.; Wang, Y.; Xiang, X.; Kong, L.; Wang, W.; Zeng, Y.; Ye, L.; et al. Torque teno virus (SANBAN isolate) ORF2 protein suppresses NF-kappaB pathways via interaction with IkappaB kinases. J. Virol. 2007, 81, 11917–11924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, R.P.; Burke, J.M.; Cox, J.C.; de Villiers, E.M.; Sullivan, C.S. A human torque teno virus encodes a microRNA that inhibits interferon signaling. PLoS Pathog. 2013, 9, e1003818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignolini, T.; Macera, L.; Antonelli, G.; Pistello, M.; Maggi, F.; Giannecchini, S. Investigation on torquetenovirus (TTV) microRNA transcriptome in vivo. Virus Res. 2016, 217, 18–22. [Google Scholar] [CrossRef]
- Tevethia, M.J.; Pipas, J.M.; Kierstead, T.; Cole, C. Requirements for immortalization of primary mouse embryo fibroblasts probed with mutants bearing deletions in the 3’ end of SV40 gene A. Virology 1988, 162, 76–89. [Google Scholar] [CrossRef]
- Chao, H.H.; Buchmann, A.M.; DeCaprio, J.A. Loss of p19 (ARF) eliminates the requirement for the pRB-binding motif in simian virus 40 large T antigen-mediated transformation. Mol. Cell. Biol. 2000, 20, 7624–7633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; DeCaprio, J.A.; Fluck, M.M.; Schaffhausen, B.S. Cellular Transformation by Simian Virus 40 and Murine Polyoma Virus T Antigens. Semin. Cancer Biol. 2009, 19, 218–228. [Google Scholar] [CrossRef] [Green Version]
- Neu, U.; Stehle, T.; Atwood, W.J. The Polyomaviridae: Contributions of virus structure to our understanding of virus receptors and infectious entry. Virology 2009, 384, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Ferreras, P.; Travaini, A.; Zapata, S.C.; Delibes, M. Short-term responses of mammalian carnivores to a sudden collapse of rabbits in Mediterranean Spain. Basic Appl. Ecol. 2011, 12, 116–124. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.K.; Jennings, N.V.; Harris, S. A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate. Mammal Rev. 2005, 35. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Poxvirus | Anellovirus | Polyomavirus | ||||
|---|---|---|---|---|---|---|
| Animal ID | Location | Tissue Type | MYXV-Tol | LepTTV1 | LepTTV2 | LepPyV1 |
| Lag01 | Spain: Toledo | Eyelid | MK836424 | MN994854 | MN994855 | MN994868 |
| Lag02 | Spain: Cuenca | Ear | - | - | - | - |
| Lag03 | Spain: Ciudad Real | Ear | - | - | - | - |
| Lag04 | Spain: Ciudad Real | Vaginal | MT072323 | - | - | - |
| Lag05 | Spain: Ciudad Real | Ear | - | - | - | - |
| Eyelid | - | - | - | |||
| Lag06 | Spain: Ciudad Real | Ear | MT072322 | - | - | - |
| Eyelid | MT072321 | - | MN994856 | - | ||
| Vaginal | MT072320 | - | - | - | ||
| Lag07 | Spain: Ciudad Real | Ear | MT072319 | - | - | - |
| Eyelid | MT072318 | - | - | - | ||
| Lag10 | Spain | Eyelid | - | MN994857 | MN994858 | MN994869 |
| Vaginal | - | MN994859 | MN994860 MN994861 | - | ||
| Lag11 | Spain: Toledo | Eyelid | - | - | MN994862 | - |
| Vaginal | - | - | - | - | ||
| Lag12 | Spain: Ciudad Real | Ear | - | - | - | - |
| Eyelid | - | - | - | - | ||
| Lag14 | Spain: Madrid | Eyelid | - | - | MN994863 MN994864 | - |
| Vaginal | MT072317 | MN994865 | MN994866 MN994867 | - | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Águeda-Pinto, A.; Kraberger, S.; Lund, M.C.; Gortázar, C.; McFadden, G.; Varsani, A.; Esteves, P.J. Coinfections of Novel Polyomavirus, Anelloviruses and a Recombinant Strain of Myxoma Virus-MYXV-Tol Identified in Iberian Hares. Viruses 2020, 12, 340. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030340
Águeda-Pinto A, Kraberger S, Lund MC, Gortázar C, McFadden G, Varsani A, Esteves PJ. Coinfections of Novel Polyomavirus, Anelloviruses and a Recombinant Strain of Myxoma Virus-MYXV-Tol Identified in Iberian Hares. Viruses. 2020; 12(3):340. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030340
Chicago/Turabian StyleÁgueda-Pinto, Ana, Simona Kraberger, Michael C. Lund, Christian Gortázar, Grant McFadden, Arvind Varsani, and Pedro J. Esteves. 2020. "Coinfections of Novel Polyomavirus, Anelloviruses and a Recombinant Strain of Myxoma Virus-MYXV-Tol Identified in Iberian Hares" Viruses 12, no. 3: 340. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030340