Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Design
2.3. Separation of Lymphocytes and Extractions of Total DNA
2.4. Absolute Quantitation of MDV in Chicken Lymphocytes Using Real-Time PCR
2.5. High-Throughput RNA-Sequencing
2.6. Bioinformatics Analysis of Differentially Expressed Gene Sets
2.7. Relative Quantification of Differentially Expressed Genes in Chicken Lymphocytes by Quantitative Reverse Transcriptase PCR (RT-qPCR)
2.8. Statistical Analysis
3. Results
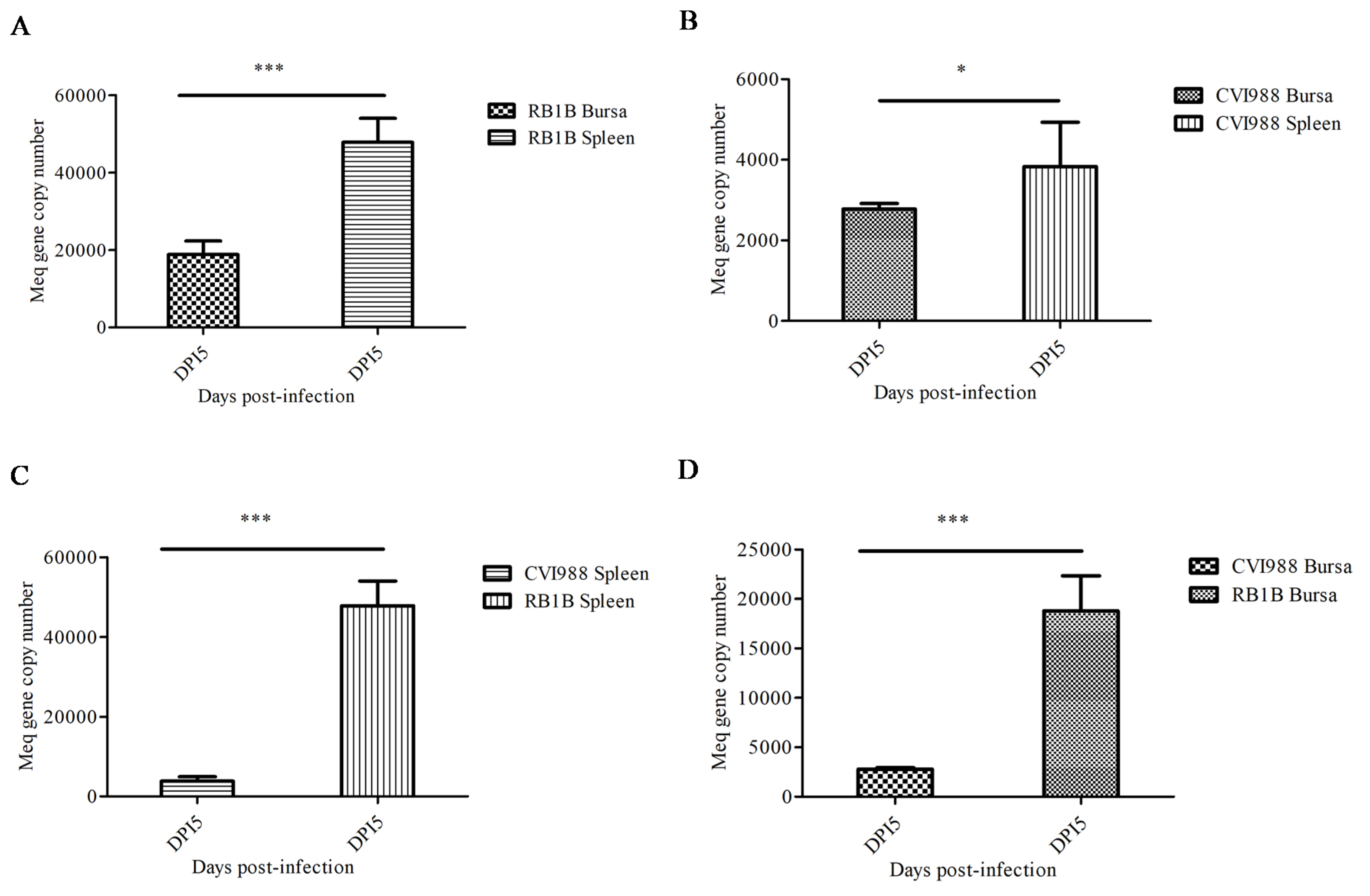
3.1. The Differences in MDV Viral Replication between a Vaccine Strain and Virulent Strain in Lymphoid Organs of the Chicken
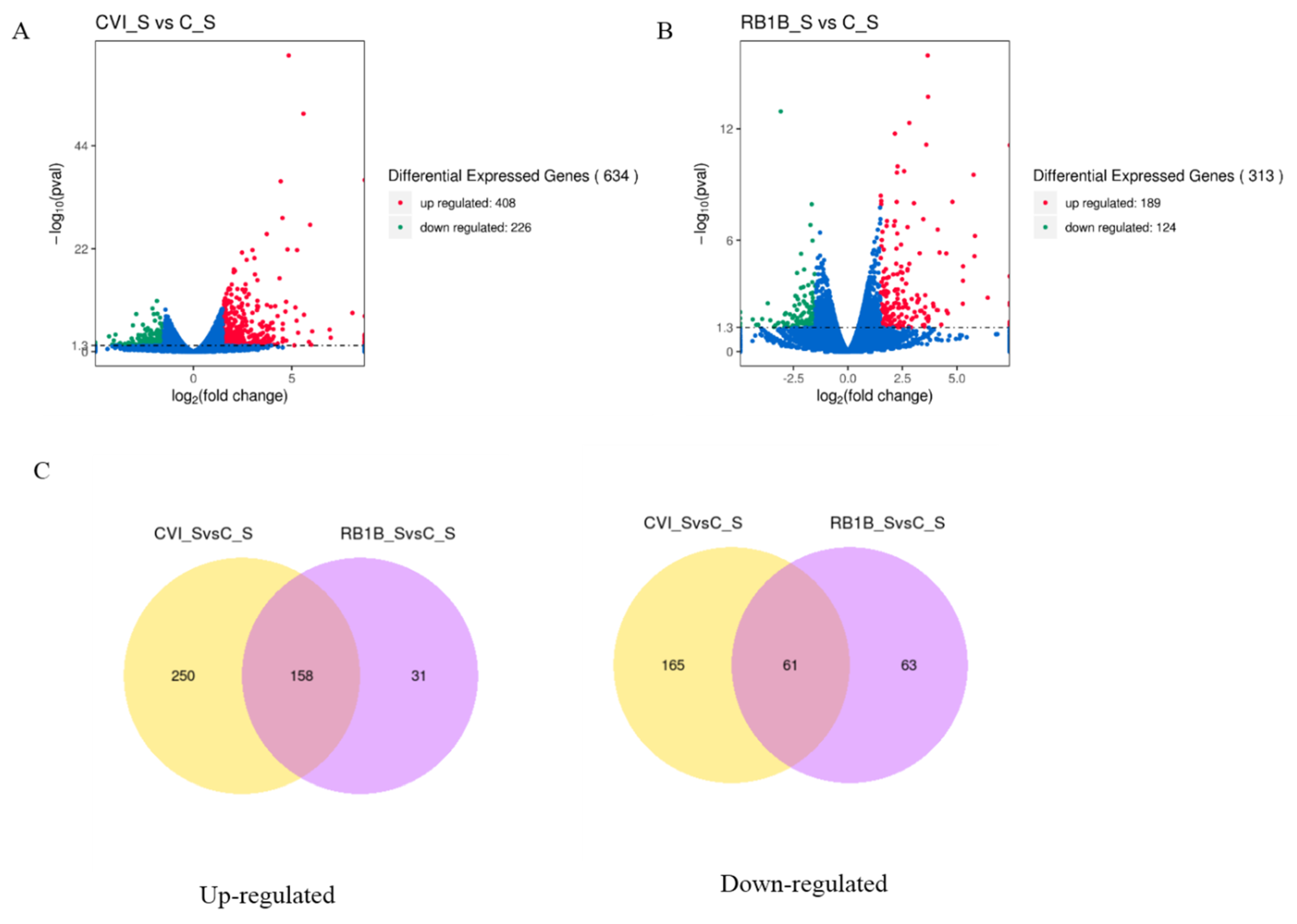
3.2. Differentially Expressed Genes (DEGs) in Splenic Lymphocytes Post-MDV Infection
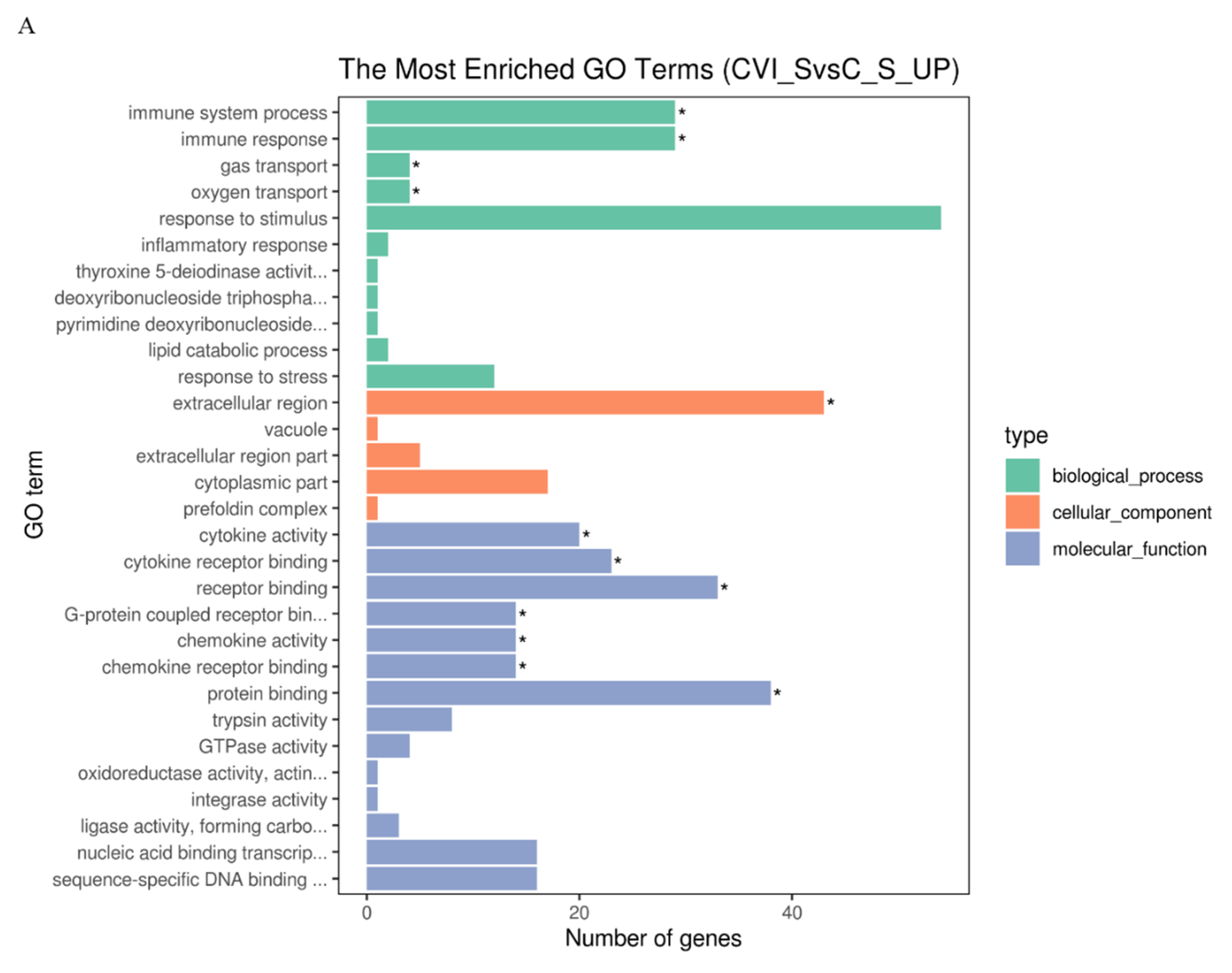
3.3. Gene Ontology (GO) Terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways of DEGs Associated with the Immune Response in Splenic Lymphocytes after MDV Infection
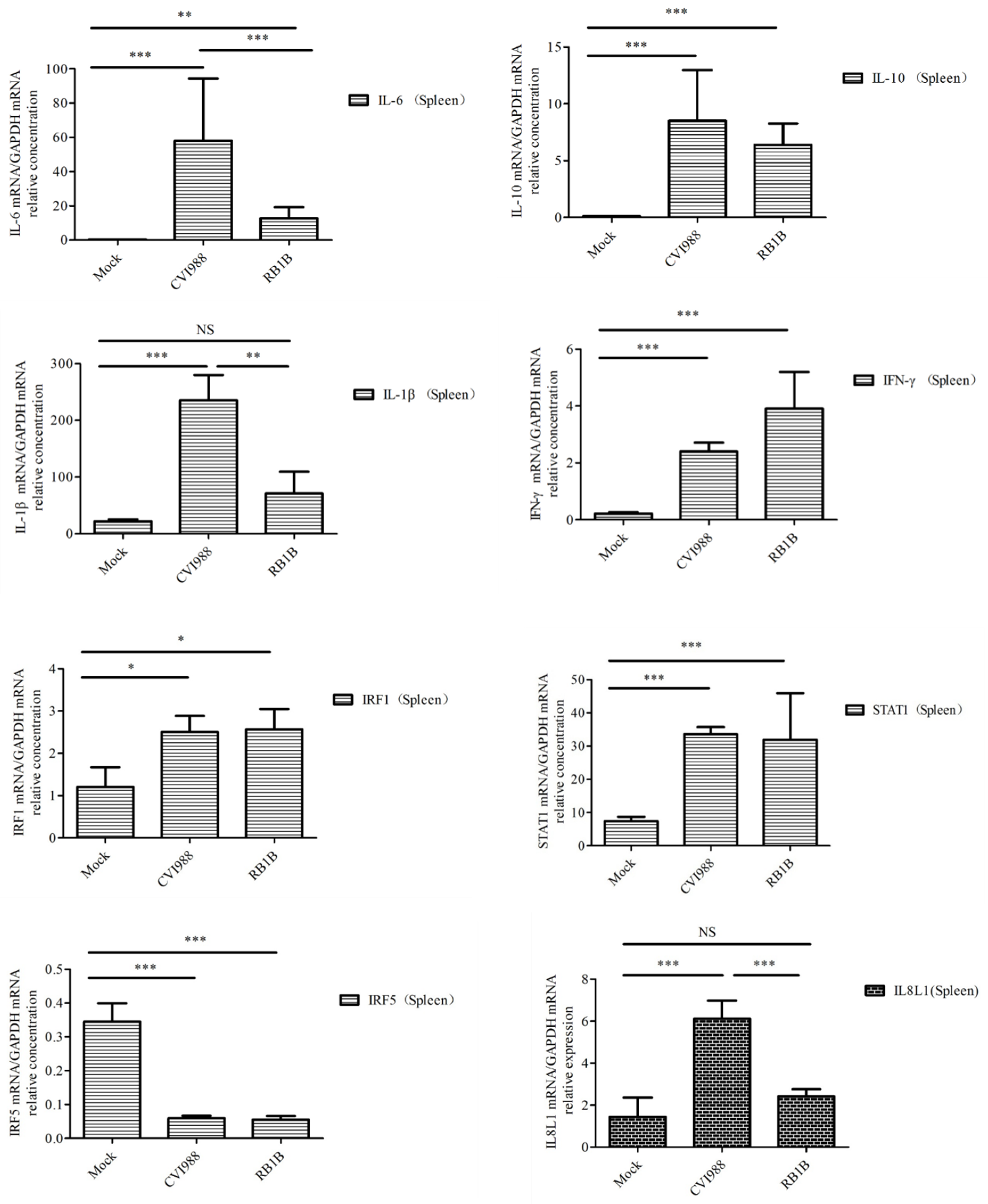
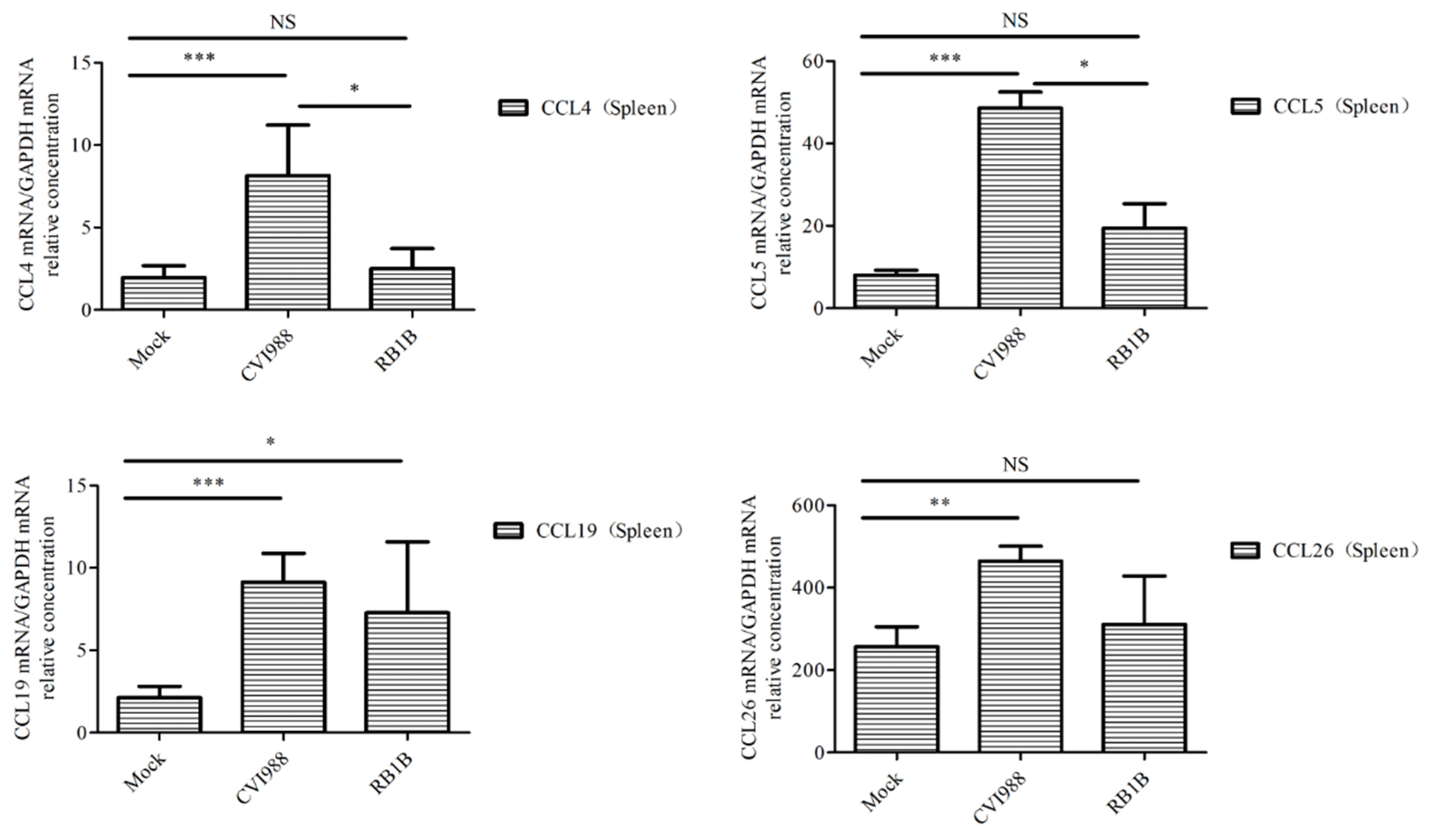
3.4. Cytokines and Chemokines Differentially Expressed in Splenic Lymphocytes after Infection with MDV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davison, F.; Nair, V. Marek’s Disease: An Evolving Problem; Elsevier: Amsterdam, The Netherland, 2004. [Google Scholar]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s disease in chickens: A review with focus on immunology. Vet. Res. 2017, 47, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazerian, K.; Witter, R.L. Cell-Free Transmission and In Vivo Replication of Marek’s Disease Virus. J. Virol. 1970, 5, 388–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Careem, M.F.; Javaheri-Vayeghan, A.; Shanmuganathan, S.; Haghighi, H.R.; Read, L.R.; Haq, K.; Hunter, D.B.; Schat, K.A.; Heidari, M.; Sharif, S. Establishment of an Aerosol-Based Marek’s Disease Virus Infection Model. Avian Dis. 2009, 53, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.A.; Burke, C.N.; Fredrickson, T.N.; DiCapua, R.A. Morphogenesis of Marek’s disease virus in feather follicle epithelium. J. Natl. Cancer Inst. 1975, 55, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Jarosinski, K.W.; Tischer, B.K.; Trapp, S.; Osterrieder, N. Marek’s disease virus: Lytic replication, oncogenesis and control. Expert Rev. Vaccines 2006, 5, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Baaten, B.J.; Staines, K.A.; Smith, L.P.; Skinner, H.; Davison, T.F.; Butter, C. Early replication in pulmonary B cells after infection with Marek’s disease herpesvirus by the respiratory route. Viral Immunol. 2009, 22, 431. [Google Scholar] [CrossRef] [Green Version]
- Calnek, B.W. Pathogenesis of Marek’s Disease Virus Infection. Curr. Top Microbiolimmunol. 2001, 255, 25–55. [Google Scholar]
- Shek, W.R.; Calnek, B.W.; Schat, K.A.; Chen, C.H. Characterization of Marek’s disease virus-infected lymphocytes: Discrimination between cytolytically and latently infected cells. J. Natl. Cancer Inst. 1983, 70, 485–491. [Google Scholar]
- Robinson, C.M.; Cheng, H.H.; Delany, M.E. Temporal Kinetics of Marek’s Disease Herpesvirus: Integration Occurs Early after Infection in Both B and T Cells. Cytogenet Genome Res. 2014, 144, 142–154. [Google Scholar] [CrossRef]
- Friedman, A.; Shalem-Meilin, E.; Heller, E.D. Marek’s disease vaccines cause temporary U-lymphocyte dysfunction and reduced resistance to infection in chicks. Avian Pathol. 1992, 21, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Abbassi, H.; Dambrine, G.; Cherel, Y.; Coudert, F.; Naciri, M. Interaction of Marek’s disease virus and Cryptosporidium baileyi in experimentally infected chickens. Avian Dis. 2000, 44, 776–789. [Google Scholar] [CrossRef]
- Kleven, S.H.; Eidson, C.S.; Anderson, D.P.; Fletcher, O.J. Decrease of antibody response to Mycoplasma synoviae in chickens infected with Marek’s disease herpesvirus. Am. J. Vet. Res. 1972, 33, 2037. [Google Scholar] [PubMed]
- Reddy, S.; Suresh, M.; Karaca, K.; Sharma, J.; McMillen, J.; Schwartz, R. Antigen-specific lymphoproliferative responses to tetanus toxoid: A means for the evaluation of Marek’s disease virus-induced immunosuppression in chickens. Vaccine 1996, 14, 1695–1702. [Google Scholar] [CrossRef]
- Barrow, A.D.; Burgess, S.C.; Baigent, S.J.; Howes, K.; Nair, V.K. Infection of macrophages by a lymphotropic herpesvirus: A new tropism for Marek’s disease virus. J. Gen. Virol. 2003, 84, 2635–2645. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Wang, L.; Marroquin, J.; Yan, J. Targeting of antigens to B cells augments antigen-specific T-cell responses and breaks immune tolerance to tumor-associated antigen MUC1. Blood 2008, 112, 2817. [Google Scholar] [CrossRef] [PubMed]
- Baigent, S.J. Replication kinetics of Marek’s disease vaccine virus in feathers and lymphoid tissues using PCR and virus isolation. J. Gen. Virol. 2005, 86, 2989–2998. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Increased Virulence of Marek’s Disease Virus Field Isolates. Avian Dis. 1997, 41, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Protective Efficacy of Marek’s Disease Vaccines. Avian Dis. 1993, 37, 53–59. [Google Scholar]
- Witter, R.L.; Nazerian, K.; Purchase, H.G.; Burgoyne, G.H. Isolation from turkeys of a cell-associated herpesvirus antigenically related to Marek’s disease virus. Am. J. Vet. Res. 1970, 31, 525–538. [Google Scholar]
- Eidson, C.S.; Page, R.K.; Kleven, S.H. Effectiveness of cell-free or cell-associated turkey herpesvirus vaccine against Marek’s disease in chickens as influenced by maternal antibody, vaccine dose, and time of exposure to Marek’s disease virus. Avian Dis. 1978, 22, 583–597. [Google Scholar] [CrossRef]
- Witter, R.L.; Sharma, J.M.; Fadly, A.M. Pathogenicity of Variant Marek’s Disease Virus Isolants in Vaccinated and Unvaccinated Chickens. Avian Dis. 1980, 24, 210. [Google Scholar] [CrossRef]
- Rispens, B.H.; Van, V.H.; Mastenbroek, N. Control of Marek’s disease in the Netherlands. I. Isolation of an avirulent Marek’s disease virus (strain CVI 988) and its use in laboratory vaccination trials. Avian Dis. 1972, 16, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L.; Lee, L.F. Polyvalent Marek’s disease vaccines: Safety, efficacy and protective synergism in chickens with maternal antibodies1. Avian Pathol. 1984, 13, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baigent, S.J.; Nair, V.K.; Le Galludec, H. Real-time pcr for differential quantification of cvi988 vaccine virus and virulent strains of marek’s disease virus. J. Virol. Methods 2016, 233, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.; Lillehoj, H.S.; Kaspers, B.; Bacon, L.D. Antigen-specific T cell proliferation following coccidianinfection. Poult. Sci. 1993, 72, 2084–2094. [Google Scholar] [CrossRef]
- Spatz, S.J.; Petherbridge, L.; Zhao, Y.; Nair, V. Comparative full-length sequence analysis of oncogenic and vaccine (rispens) strains of marek’s disease virus. J. Gen. Virol. 2007, 88, 1080–1096. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, S.; Ma, C. Absolute quantification of a very virulent Marek’s disease virus dynamic quantity and distributions in different tissues 1. Poult. Sci. 2015, 94, 1150. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 1, 1. [Google Scholar]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level-the DESeq Package; European Molecular Biology Laboratory (EMBL): Heidelberg, Germany, 2012; Volume 10, pp. 22–45. [Google Scholar]
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing Under Dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar]
- Kaiser, P.; Underwood, G.; Davison, F. Differential Cytokine Responses following Marek’s Disease Virus Infection of Chickens Differing in Resistance to Marek’s Disease. J. Virol. 2003, 77, 762–768. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.; Sadeyen, J.R.; Paton, I.R.; Hocking, P.M.; Salmon, N.; Fife, M.; Nair, V.; Burt, D.W.; Kaiser, P. Systems Analysis of Immune Responses in Marek’s Disease Virus-Infected Chickens Identifies a Gene Involved in Susceptibility and Highlights a Possible Novel Pathogenicity Mechanism. J. Virol. 2011, 85, 11146–11158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Careem, M.F.; Hunter, B.D.; Parvizi, P.; Haghighi, H.R.; Thanthrige-Don, N.; Sharif, S. Cytokine gene expression patterns associated with immunization against Marek’s disease in chickens. Vaccine 2007, 25, 0–432. [Google Scholar] [CrossRef] [PubMed]
- Sarson, A.J.; Parvizi, P.; Lepp, D.; Quinton, M.; Sharif, S. Transcriptional analysis of host responses to Marek’s disease virus infection in genetically resistant and susceptible chickens. Anim. Genet. 2008, 39, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Calnek, B.W.; Harris, R.W.; Buscaglia, C.; Schat, K.A.; Lucio, B. Relationship between the Immunosuppressive Potential and the Pathotype of Marek’s Disease Virus Isolates. Avian Dis. 1998, 42, 124–132. [Google Scholar] [CrossRef]
- Berthault, C.; Larcher, T.; Härtle, S.; Vautherot, J.F.; Trapp-Fragnet, L.; Denesvre, C. Atrophy of primary lymphoid organs induced by Marek’s disease virus during early infection is associated with increased apoptosis, inhibition of cell proliferation and a severe B-lymphopenia. Vet. Res. 2018, 49, 31. [Google Scholar] [CrossRef] [Green Version]
- Baigent, S.J.; Ross, L.J.; Davison, T.F. Differential susceptibility to Marek’s disease is associated with differences in number, but not phenotype or location, of pp38+ lymphocytes. J. Gen. Virol. 1998, 79, 2795–2802. [Google Scholar] [CrossRef] [Green Version]
- Calnek, B.W.; Schat, K.A.; Ross, L.J.; Chen, C.L. Further characterization of Marek’s disease virus-infected lymphocytes. II In vitro infection. Int. J. Cancer 1984, 33, 399–406. [Google Scholar] [CrossRef]
- Dang, L.; Teng, M.; Li, H.W.; Li, H.Z.; Ma, S.M.; Zhao, P.; Li, X.J.; Deng, R.G.; Zhang, G.P.; Luo, J. Dynamic Changes in the Splenic Transcriptome of Chickens during the Early Infection and Progress of Marek’s Disease. Sci. Rep. 2017, 7, 11648. [Google Scholar] [CrossRef] [Green Version]
- Jeurissen, S.H.M. Structure and function of the chicken spleen. Res. Immunol. 1991, 142, 352–355. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Radzio-Basu, J.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional Innate Immune Response of the Developing Chicken Embryo to Newcastle Disease Virus Infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T. Factors Affecting B-Cell Growth and Differentiation. Annu. Rev. Immunol. 1985, 3, 133–157. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Tanner, E.; Tosato, G. Regulation of B-cell growth and immunoglobulin gene transcription by interleukin-6. Blood 1992, 79, 452–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdin, N.; Rousset, F.; Banchereau, J. B-Cell-Derived IL-10: Production and Function. Methods 1997, 11, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Nova-Lamperti, E.; Chana, P.; Mobillo, P.; Runglall, M.; Kamra, Y.; McGregor, R.; Lord, G.M.; Lechler, R.I.; Lombardi, G.; Hernandez-Fuentes, M.P. Increased CD40 ligation and reduced BCR signalling leads to higher IL-10 production in B cells from tolerant kidney transplant patients. Transplantation 2017, 101, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, J.P.; Cardin, R.D.; Branum, K.C.; Doherty, P.C. CD4(+) T cell-mediated control of a gamma-herpesvirus in B cell-deficient mice is mediated by IFN-gamma. Proc. Natl. Acad. Sci. USA 1999, 96, 5135–5140. [Google Scholar] [CrossRef] [Green Version]
- Jud, A.; Kotur, M.; Berger, C.; Gysin, C.; Nadal, D.; Lünemann, A. Tonsillar CD56 bright NKG2A +, NK cells restrict primary Epstein-Barr virus infection in B cells via IFN-γ. Oncotarget 2017, 8, 6130–6141. [Google Scholar] [CrossRef] [Green Version]
- Moser, B.; Loetsche, P. Lymphocyte traffic control by chemokines. Nat. Immunol. 2001, 2, 123–128. [Google Scholar] [CrossRef]
- Luther, S.A.; Cyster, J.G. Chemokines as regulators of T cell differentiation. Nat. Immunol. 2001, 2, 102–107. [Google Scholar] [CrossRef]
- Cocchi, F.; DeVico, A.L.; Garzino-Demo, A.; Arya, S.K.; Gallo, R.C.; Lusso, P. Identification of RANTES, MIP-1alpha, and MIP-1beta as the Major HIV-Suppressive Factors Produced by CD8+ T Cells. Science 1995, 270, 1811–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bystry, R.S.; Aluvihare, V.; Welch, K.A.; Kallikourdis, M.; Betz, A.G. B cells and professional APCs recruit regulatory T cells via CCL4. Nat. Immunol. 2001, 2, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, A.N.; Wang, Y.; Huang, Y.C.; Irvine, D.J. Homeostatic Lymphoid Chemokines Synergize with Adhesion Ligands to Trigger T and B Lymphocyte Chemokinesis. J. Immunol. 2006, 177, 2340–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucendo, A.J.; de Rezende, L.; Comas, C.; Caballero, T.; Bellón, T. Treatment with topical steroids downregulates IL-5, eotaxin-1/CCL11, and eotaxin-3/CCL26 gene expression in eosinophilic esophagitis. Am. J. Gastroenterol 2008, 103, 2184–2193. [Google Scholar] [CrossRef] [PubMed]
- Kagami, S.; Kakinuma, T.; Saeki, H.; Tsunemi, Y.; Fujita, H.; Nakamura, K.; Takekoshi, T.; Kishimoto, M.; Mitsui, H.; Torii, H. Significant elevation of serum levels of eotaxin-3/CCL26, but not of eotaxin-2/CCL24, in patients with atopic dermatitis: Serum eotaxin-3/CCL26 levels reflect the disease activity of atopic dermatitis. Clin. Exp. Immunol. 2003, 134, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.T.; Selvaraj, R.K.; Kamil, J.P.; Osterrieder, N.; Kaufer, B.B. Marek’s disease viral interleukin-8 promotes lymphoma formation through targeted recruitment of b cells and CD4+ CD25+ T cells. J. Virol. 2012, 86, 8536–8545. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers * | Sequence (5′-3′) | Amplicon Size (bp) |
|---|---|---|
| GAPDH-F GAPDH-R | AGAACATCATCCCAGCGT AGCCTTCACTACCCTCTTG | 182 bp |
| IL-6-F IL-6-R | AAATCCCTCCTCGCCAATCT CCCTCACGGTCTTCTCCATAAA | 106 bp |
| IL-4-F IL-4-R | AGCACTGCCACAAGAACCT GCTAGTTGGTGGAAGAAGGTAC | 140 bp |
| IL-10-F IL-10-R | GCTGCGCTTCTACACAGATG TCCCGTTCTCATCCATCTTC | 203 bp |
| IL-1β-F IL-1β-R | CCCGCTTCATCTTCTACCGC GCTTGTAGGTGGCGATGTTG | 159 bp |
| IFN-γ-F IFN-γ-R | GCCGCACATCAAACACATATCTG GCGCTGGATTCTCAAGTCGTTC | 127 bp |
| IRF1-F IRF1-R | TGGGGTCGTCCTCAGAAGAT AGAAGCCTTTCCCCTCAACG | 105 bp |
| IRF5-F IRF5-R | AACAAGAGCCGTGAGTTCCG CTGAAGTCATCCCCGGTCAC | 122 bp |
| STAT1-F STAT1-R | CCAAGCGGATGGGCTTCTAT GCTGATCCGCGTTAAGTCCT | 163 bp |
| IL8L1-F IL8L1-R | CCTCACTGCAAGAATGTGGA GGAGGAGGTAGGACGTTTTTG | 153 bp |
| CCL4-F CCL4-R | CCTTCAGCTTTGTGGCAGAC GGTGATGAACACAACACCAGC | 71 bp |
| CCL5-F CCL5-R | GGCTGATACAACCGTGTGCT CCTGGTGATGAACACAACTGC | 121 bp |
| CCL19-F CCL19-R | TGTATGCTGGCAACAACGTC CACAGAGACGCTTGCCCTTT | 156 bp |
| CCL26-F CCL26-R | TTCAGATGGCCTACCCACAAC GACTCCTCGGGGTTTACACA | 162 bp |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, H.; Kong, Z.; Mehboob, A.; Jiang, B.; Xu, J.; Cai, Y.; Liu, W.; Hong, J.; Li, Y. Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection. Viruses 2020, 12, 354. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030354
Jin H, Kong Z, Mehboob A, Jiang B, Xu J, Cai Y, Liu W, Hong J, Li Y. Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection. Viruses. 2020; 12(3):354. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030354
Chicago/Turabian StyleJin, Huan, Zimeng Kong, Arslan Mehboob, Bo Jiang, Jian Xu, Yunhong Cai, Wenxiao Liu, Jiabing Hong, and Yongqing Li. 2020. "Transcriptional Profiles Associated with Marek’s Disease Virus in Bursa and Spleen Lymphocytes Reveal Contrasting Immune Responses during Early Cytolytic Infection" Viruses 12, no. 3: 354. https://0-doi-org.brum.beds.ac.uk/10.3390/v12030354