Broad Antiviral Activity of Ginkgolic Acid against Chikungunya, Mayaro, Una, and Zika Viruses

 ,
,  and
and 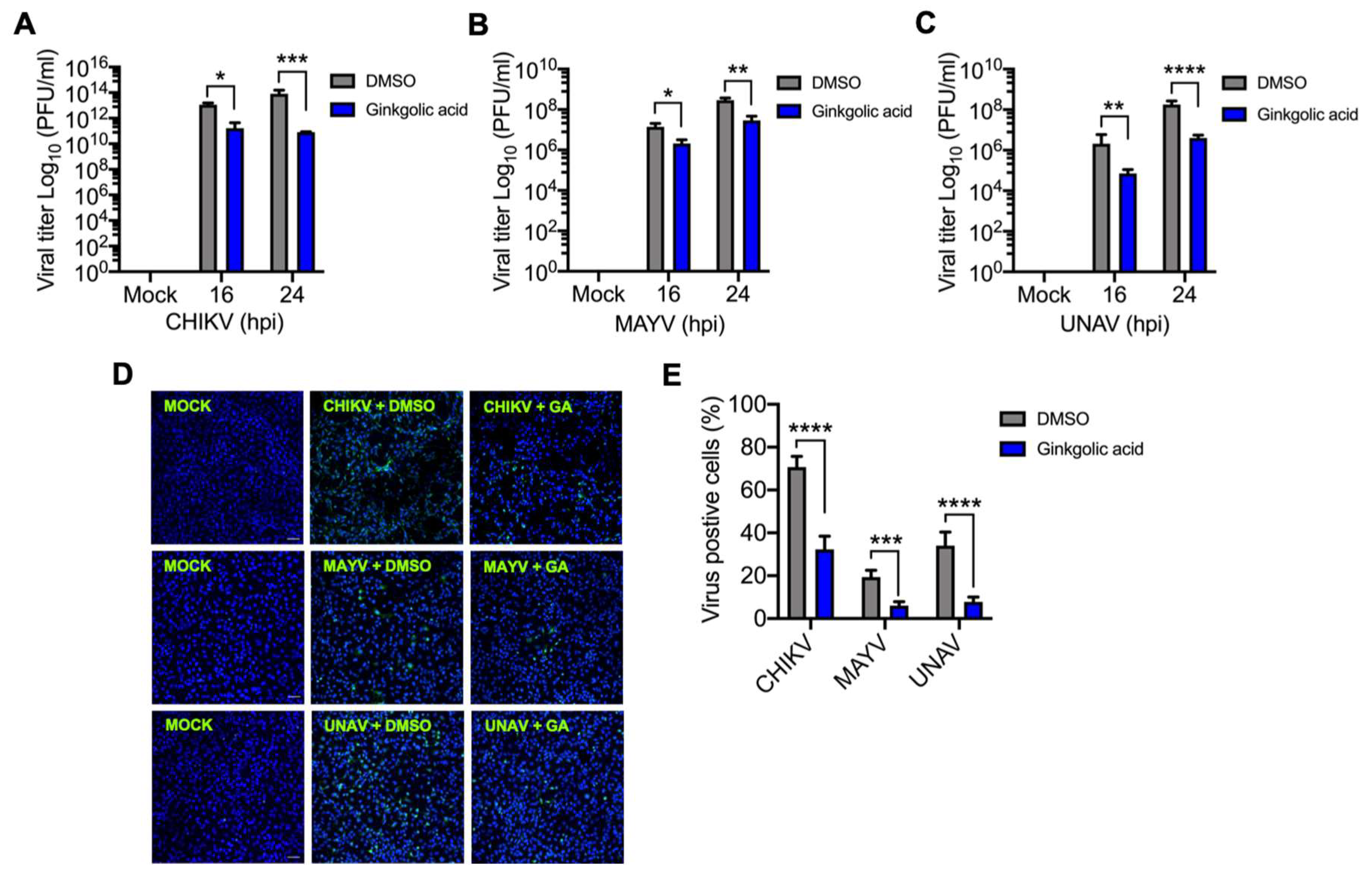{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line Cultures and Reagents
2.2. Virus Strains and Propagation
2.3. Cytotoxicity Analysis
2.4. Virus Plaque-Forming Assay
2.5. Viral Infection Assays
2.6. Immunofluorescence Assay
2.7. Protein Analysis
2.8. Statistical Analysis
3. Results
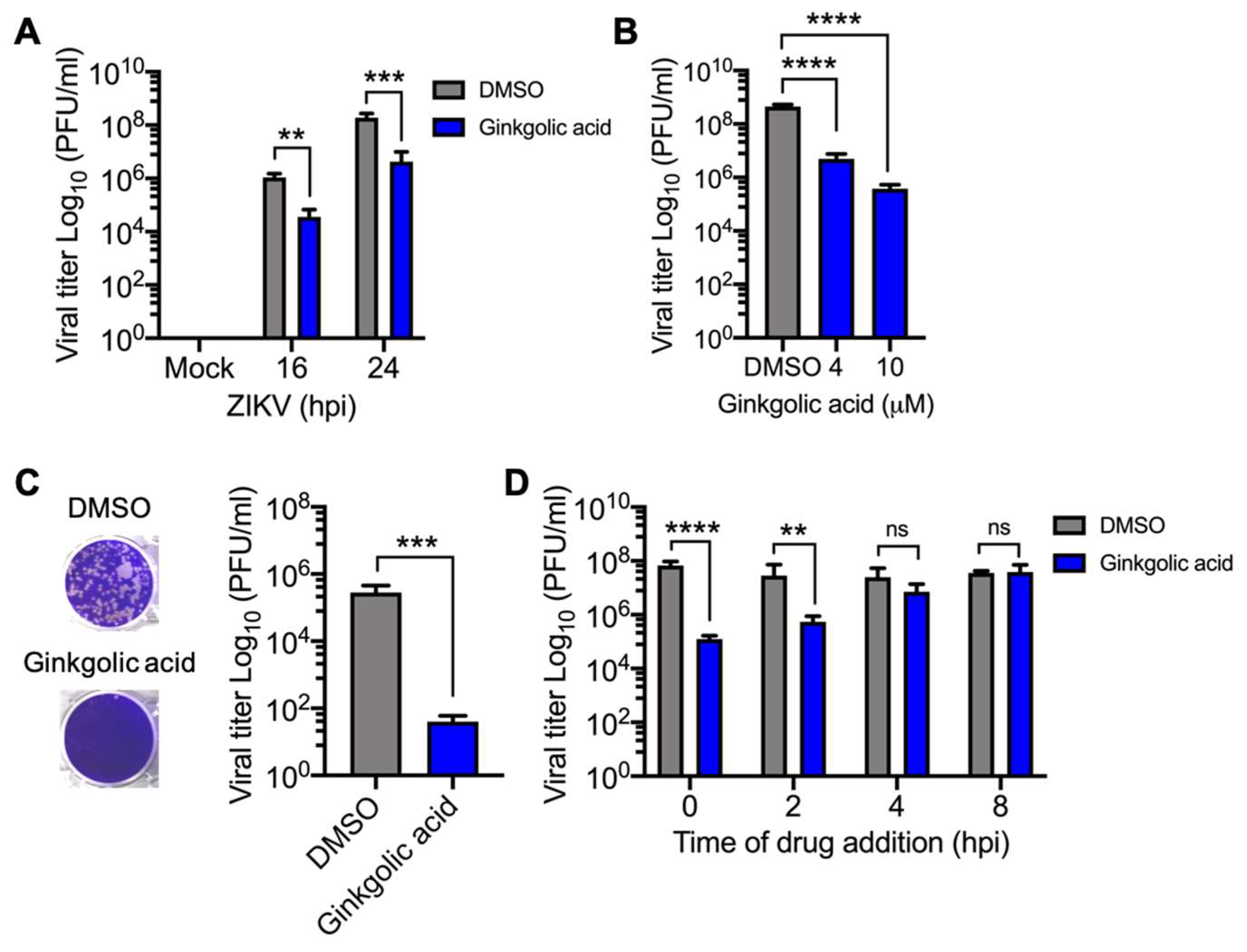
3.1. Ginkgolic Acid Reduces Alphavirus Progeny Yield
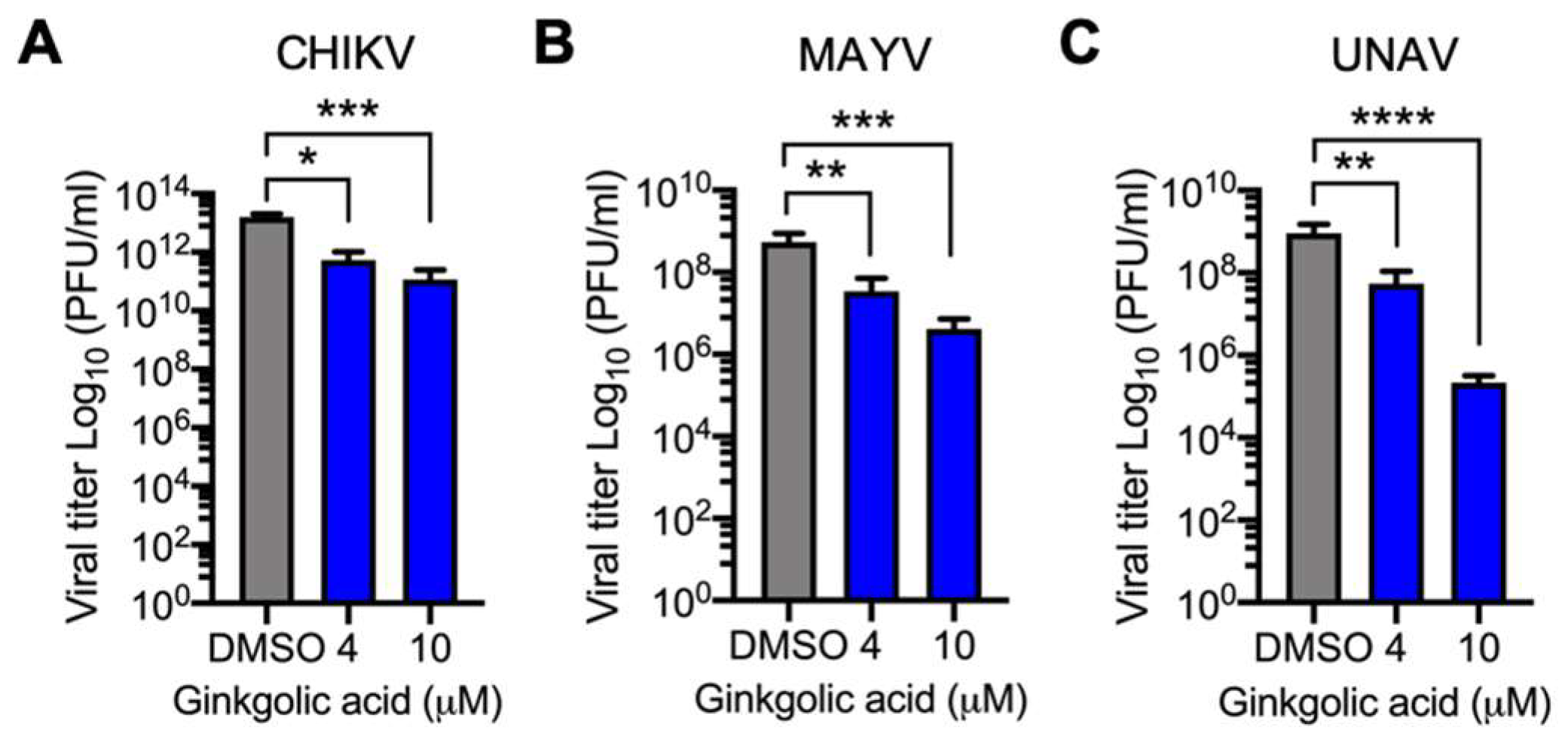
3.2. Alphavirus Titer Decrease Is Ginkgolic Acid Concentration-Dependent
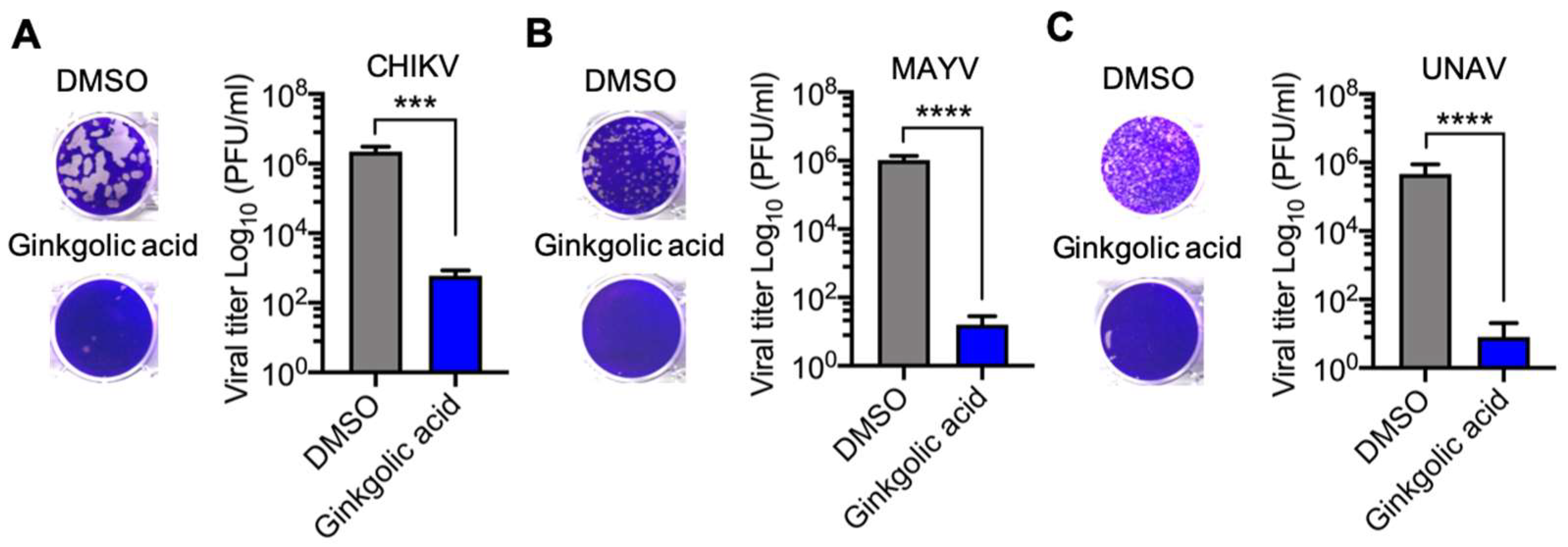
3.3. Virucidal Activity of Ginkgolic Acid against Alphavirus
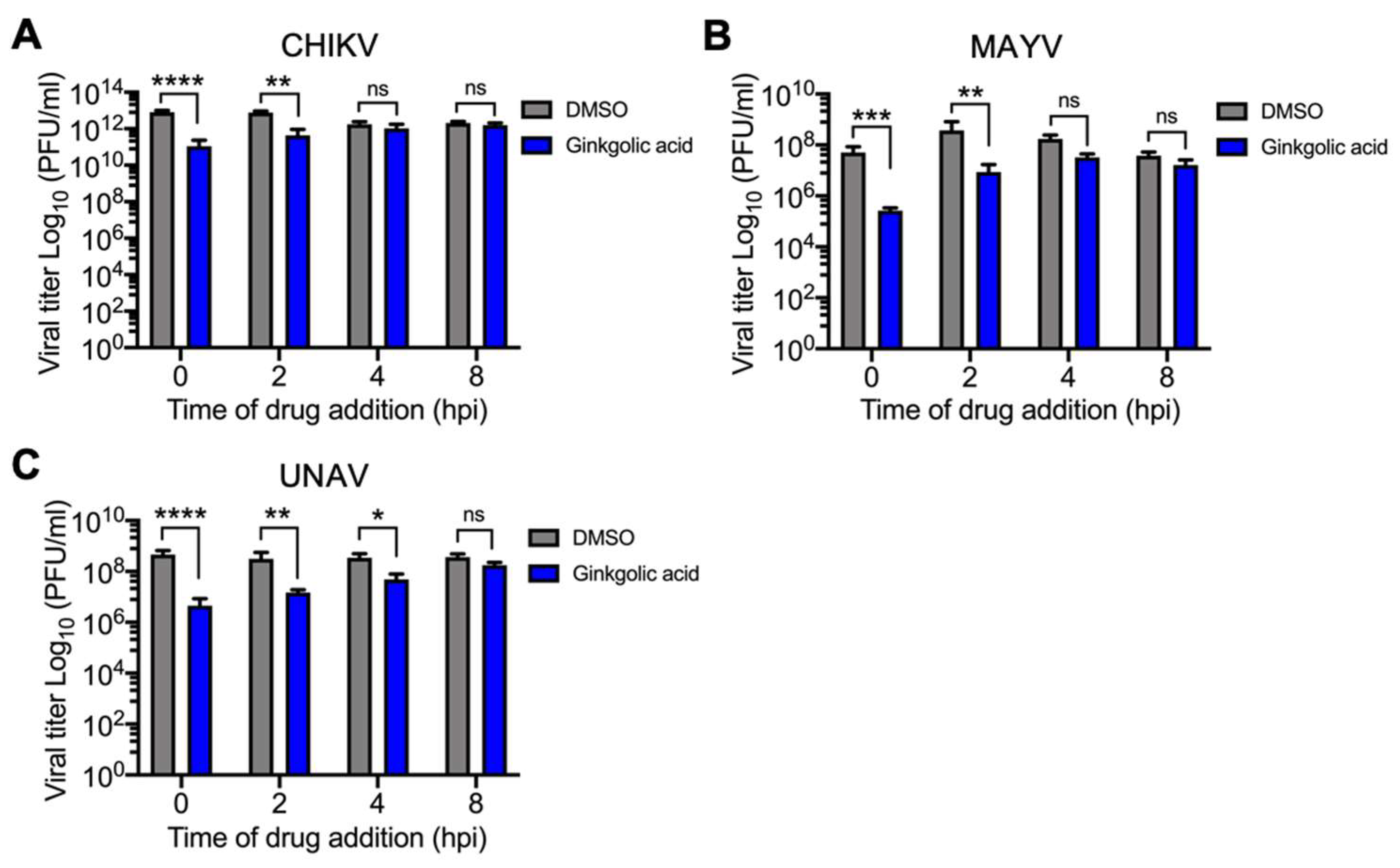
3.4. Ginkgolic Acid Disrupts an Early Stage in the Alphavirus Replication Cycle
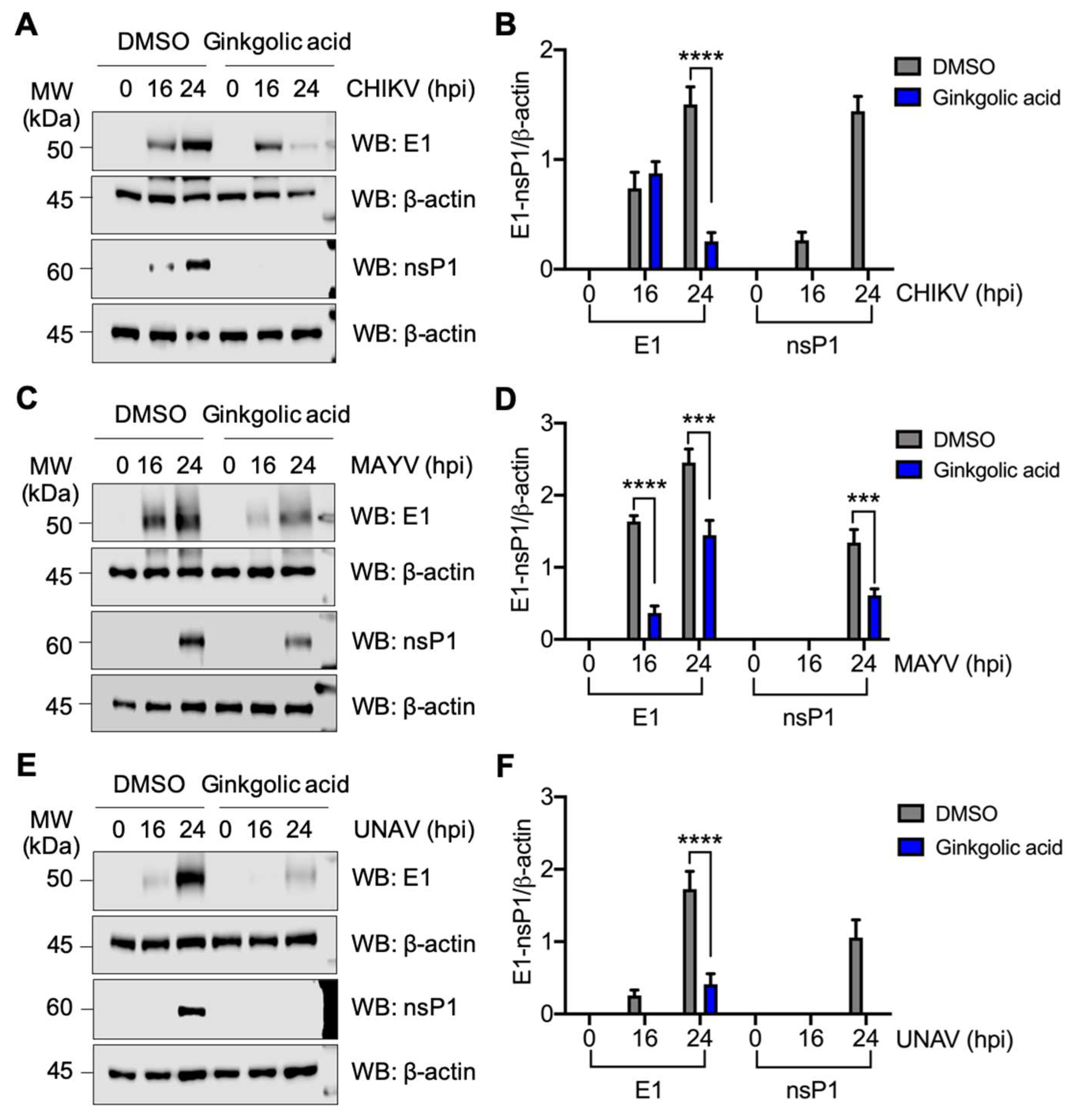
3.5. Ginkgolic acid Suppresses the Expression of Alphavirus Proteins
3.6. Ginkgolic Acid also Blocks Zika Virus Replication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weaver, S.C. Urbanization and geographic expansion of zoonotic arboviral diseases: Mechanisms and potential strategies for prevention. Trends Microbiol. 2013, 21, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.Y.; Ng, M.M.; Chu, J.J. Replication of alphaviruses: A review on the entry process of alphaviruses into cells. Adv. Virol 2011, 2011, 249640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazeaud, C.; Freppel, W.; Chatel-Chaix, L. The Multiples Fates of the Flavivirus RNA Genome During Pathogenesis. Front. Genet. 2018, 9, 595. [Google Scholar] [CrossRef] [PubMed]
- Conway, M.J.; Colpitts, T.M.; Fikrig, E. Role of the Vector in Arbovirus Transmission. Annu. Rev. Virol 2014, 1, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, E.B.; Kramer, L.D. Zika Virus Mosquito Vectors: Competence, Biology, and Vector Control. J. Infect. Dis. 2017, 216, S976–S990. [Google Scholar] [CrossRef] [Green Version]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef]
- Rupp, J.C.; Sokoloski, K.J.; Gebhart, N.N.; Hardy, R.W. Alphavirus RNA synthesis and non-structural protein functions. J. Gen. Virol 2015, 96, 2483–2500. [Google Scholar] [CrossRef]
- Martins, K.A.; Gregory, M.K.; Valdez, S.M.; Sprague, T.R.; Encinales, L.; Pacheco, N.; Cure, C.; Porras-Ramirez, A.; Rico-Mendoza, A.; Chang, A.; et al. Neutralizing Antibodies from Convalescent Chikungunya Virus Patients Can Cross-Neutralize Mayaro and Una Viruses. Am. J. Trop Med. Hyg. 2019, 100, 1541–1544. [Google Scholar] [CrossRef]
- Suhrbier, A.; Jaffar-Bandjee, M.C.; Gasque, P. Arthritogenic alphaviruses--an overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef]
- Lwande, O.W.; Obanda, V.; Bucht, G.; Mosomtai, G.; Otieno, V.; Ahlm, C.; Evander, M. Global emergence of Alphaviruses that cause arthritis in humans. Infect. Ecol. Epidemiol. 2015, 5, 29853. [Google Scholar] [CrossRef] [Green Version]
- Bonifay, T.; Prince, C.; Neyra, C.; Demar, M.; Rousset, D.; Kallel, H.; Nacher, M.; Djossou, F.; Epelboin, L.; Char Chik Working, g. Atypical and severe manifestations of chikungunya virus infection in French Guiana: A hospital-based study. PLoS ONE 2018, 13, e0207406. [Google Scholar] [CrossRef] [Green Version]
- Brito, C.A.A. Alert: Severe cases and deaths associated with Chikungunya in Brazil. Rev. Soc. Bras. Med. Trop. 2017, 50, 585–589. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.V.; Murdock, C.C.; Drake, J.M. Anticipating Emerging Mosquito-borne Flaviviruses in the USA: What Comes after Zika? Trends Parasitol. 2018, 34, 544–547. [Google Scholar] [CrossRef]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef]
- Vasconcelos, P.F.; Calisher, C.H. Emergence of Human Arboviral Diseases in the Americas, 2000-2016. Vector Borne Zoonotic Dis. 2016, 16, 295–301. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [Green Version]
- Van Beek, T.A.; Wintermans, M.S. Preparative isolation and dual column high-performance liquid chromatography of ginkgolic acids from Ginkgo biloba. J. Chromatogr. A 2001, 930, 109–117. [Google Scholar] [CrossRef]
- Qiao, L.; Zheng, J.; Jin, X.; Wei, G.; Wang, G.; Sun, X.; Li, X. Ginkgolic acid inhibits the invasiveness of colon cancer cells through AMPK activation. Oncol. Lett. 2017, 14, 5831–5838. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Duan, W.; Han, S.; Lei, J.; Xu, Q.; Chen, X.; Jiang, Z.; Nan, L.; Li, J.; Chen, K.; et al. Ginkgolic acid suppresses the development of pancreatic cancer by inhibiting pathways driving lipogenesis. Oncotarget 2015, 6, 20993–21003. [Google Scholar] [CrossRef] [Green Version]
- Hua, Z.; Wu, C.; Fan, G.; Tang, Z.; Cao, F. The antibacterial activity and mechanism of ginkgolic acid C15:1. BMC Biotechnol. 2017, 17, 5. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.X.; Wu, L.; Jiang, X.G.; Feng, Y.Y.; Cao, J.P. Anti-Toxoplasma gondii activity of GAS in vitro. J. Ethnopharmacol. 2008, 118, 503–507. [Google Scholar] [CrossRef]
- Lu, J.M.; Yan, S.; Jamaluddin, S.; Weakley, S.M.; Liang, Z.; Siwak, E.B.; Yao, Q.; Chen, C. Ginkgolic acid inhibits HIV protease activity and HIV infection in vitro. Med. Sci. Monit. 2012, 18, BR293–BR298. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, I.; Ito, A.; Hirai, G.; Nishimura, S.; Kawasaki, H.; Saitoh, H.; Kimura, K.; Sodeoka, M.; Yoshida, M. Ginkgolic acid inhibits protein SUMOylation by blocking formation of the E1-SUMO intermediate. Chem. Biol. 2009, 16, 133–140. [Google Scholar] [CrossRef]
- Powers, A.M.; Aguilar, P.V.; Chandler, L.J.; Brault, A.C.; Meakins, T.A.; Watts, D.; Russell, K.L.; Olson, J.; Vasconcelos, P.F.; Da Rosa, A.T.; et al. Genetic relationships among Mayaro and Una viruses suggest distinct patterns of transmission. Am. J. Trop. Med. Hyg. 2006, 75, 461–469. [Google Scholar] [CrossRef]
- Causey, O.R.; Casals, J.; Shope, R.E.; Udomsakdi, S. Aura and Una, Two New Group a Arthropod-Borne Viruses. Am. J. Trop. Med. Hyg. 1963, 12, 777–781. [Google Scholar] [CrossRef]
- Carrera, J.P.; Diaz, Y.; Denis, B.; Barahona de Mosca, I.; Rodriguez, D.; Cedeno, I.; Arauz, D.; Gonzalez, P.; Cerezo, L.; Moreno, L.; et al. Unusual pattern of chikungunya virus epidemic in the Americas, the Panamanian experience. PLoS Negl. Trop Dis. 2017, 11, e0005338. [Google Scholar] [CrossRef]
- Llamas-Gonzalez, Y.Y.; Campos, D.; Pascale, J.M.; Arbiza, J.; Gonzalez-Santamaria, J. A Functional Ubiquitin-Proteasome System is Required for Efficient Replication of New World Mayaro and Una Alphaviruses. Viruses 2019, 11, 370. [Google Scholar] [CrossRef] [Green Version]
- Marston, H.D.; Folkers, G.K.; Morens, D.M.; Fauci, A.S. Emerging viral diseases: Confronting threats with new technologies. Sci. Transl. Med. 2014, 6, 253ps210. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.S.; Higgs, S.; Vanlandingham, D.L. Emergence and re-emergence of mosquito-borne arboviruses. Curr. Opin. Virol. 2019, 34, 104–109. [Google Scholar] [CrossRef]
- Liang, G.; Gao, X.; Gould, E.A. Factors responsible for the emergence of arboviruses; strategies, challenges and limitations for their control. Emerg. Microbes Infect. 2015, 4, e18. [Google Scholar] [CrossRef]
- Ugwu, C.E.; Jiang, Y.Y.; Wu, L.; Xu, Y.X.; Yin, J.H.; Duan, L.P.; Chen, S.X.; Liu, H.; Pan, W.; Quan, H.; et al. In vitro Screening of Ginkgolic Acids for Antiparasitic Activity against Cryptosporidium andersoni. Biomed. Environ. Sci. 2019, 32, 300–303. [Google Scholar] [CrossRef]
- Li, J.; Li, A.; Li, M.; Liu, Y.; Zhao, W.; Gao, D. Ginkgolic acid exerts an anti-inflammatory effect in human umbilical vein endothelial cells induced by ox-LDL. Pharmazie 2018, 73, 408–412. [Google Scholar] [CrossRef]
- Zhou, C.; Li, X.; Du, W.; Feng, Y.; Kong, X.; Li, Y.; Xiao, L.; Zhang, P. Antitumor effects of ginkgolic acid in human cancer cell occur via cell cycle arrest and decrease the Bcl-2/Bax ratio to induce apoptosis. Chemotherapy 2010, 56, 393–402. [Google Scholar] [CrossRef]
- Lee, J.S.; Hattori, M.; Kim, J. Inhibition of HIV-1 protease and RNase H of HIV-1 reverse transcriptase activities by long chain phenols from the sarcotestas of Ginkgo biloba. Planta Med. 2008, 74, 532–534. [Google Scholar] [CrossRef] [Green Version]
- Mounce, B.C.; Cesaro, T.; Carrau, L.; Vallet, T.; Vignuzzi, M. Curcumin inhibits Zika and chikungunya virus infection by inhibiting cell binding. Antivir. Res. 2017, 142, 148–157. [Google Scholar] [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral activity of luteolin against Japanese encephalitis virus. Virus Res. 2016, 220, 112–116. [Google Scholar] [CrossRef]
- Weber, C.; Sliva, K.; von Rhein, C.; Kummerer, B.M.; Schnierle, B.S. The green tea catechin, epigallocatechin gallate inhibits chikungunya virus infection. Antivir. Res. 2015, 113, 1–3. [Google Scholar] [CrossRef]
- Lin, L.T.; Chen, T.Y.; Lin, S.C.; Chung, C.Y.; Lin, T.C.; Wang, G.H.; Anderson, R.; Lin, C.C.; Richardson, C.D. Broad-spectrum antiviral activity of chebulagic acid and punicalagin against viruses that use glycosaminoglycans for entry. BMC Microbiol. 2013, 13, 187. [Google Scholar] [CrossRef] [Green Version]
- Bertol, J.W.; Rigotto, C.; de Padua, R.M.; Kreis, W.; Barardi, C.R.; Braga, F.C.; Simoes, C.M. Antiherpes activity of glucoevatromonoside, a cardenolide isolated from a Brazilian cultivar of Digitalis lanata. Antivir. Res. 2011, 92, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, S.; Sauter, D.; Wang, K.; Zhang, R.; Sun, B.; Karioti, A.; Bilia, A.R.; Efferth, T.; Schwarz, W. Kaempferol derivatives as antiviral drugs against the 3a channel protein of coronavirus. Planta Med. 2014, 80, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Berg, K.; Braun, C.; Krug, I.; Schrenk, D. Evaluation of the cytotoxic and mutagenic potential of three ginkgolic acids. Toxicology 2015, 327, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Zeng, S. Cytotoxicity of ginkgolic acid in HepG2 cells and primary rat hepatocytes. Toxicol. Lett. 2009, 187, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Peng, Y.; Shang, E.; Zhao, M.; Yan, L.; Zhu, Z.; Tao, J.; Su, S.; Guo, S.; Duan, J.A. Metabolic profiling of the hepatotoxicity and nephrotoxicity of Ginkgolic acids in rats using ultra-performance liquid chromatography-high-definition mass spectrometry. Chem. Biol. Interact. 2017, 273, 11–17. [Google Scholar] [CrossRef]
- Xia, H.; Wang, X.; Li, L.; Wang, S.; Guo, C.; Liu, Y.; Yu, L.; Jiang, H.; Zeng, S. Development of high performance liquid chromatography/electrospray ionization mass spectrometry for assay of ginkgolic acid (15:1) in rat plasma and its application to pharmacokinetics study. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 2701–2706. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos, D.; Navarro, S.; Llamas-González, Y.Y.; Sugasti, M.; González-Santamaría, J. Broad Antiviral Activity of Ginkgolic Acid against Chikungunya, Mayaro, Una, and Zika Viruses. Viruses 2020, 12, 449. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040449
Campos D, Navarro S, Llamas-González YY, Sugasti M, González-Santamaría J. Broad Antiviral Activity of Ginkgolic Acid against Chikungunya, Mayaro, Una, and Zika Viruses. Viruses. 2020; 12(4):449. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040449
Chicago/Turabian StyleCampos, Dalkiria, Susana Navarro, Yessica Yadira Llamas-González, Madelaine Sugasti, and José González-Santamaría. 2020. "Broad Antiviral Activity of Ginkgolic Acid against Chikungunya, Mayaro, Una, and Zika Viruses" Viruses 12, no. 4: 449. https://0-doi-org.brum.beds.ac.uk/10.3390/v12040449