Antiviral and Cytotoxic Activity of Different Plant Parts of Banana (Musa spp.)

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
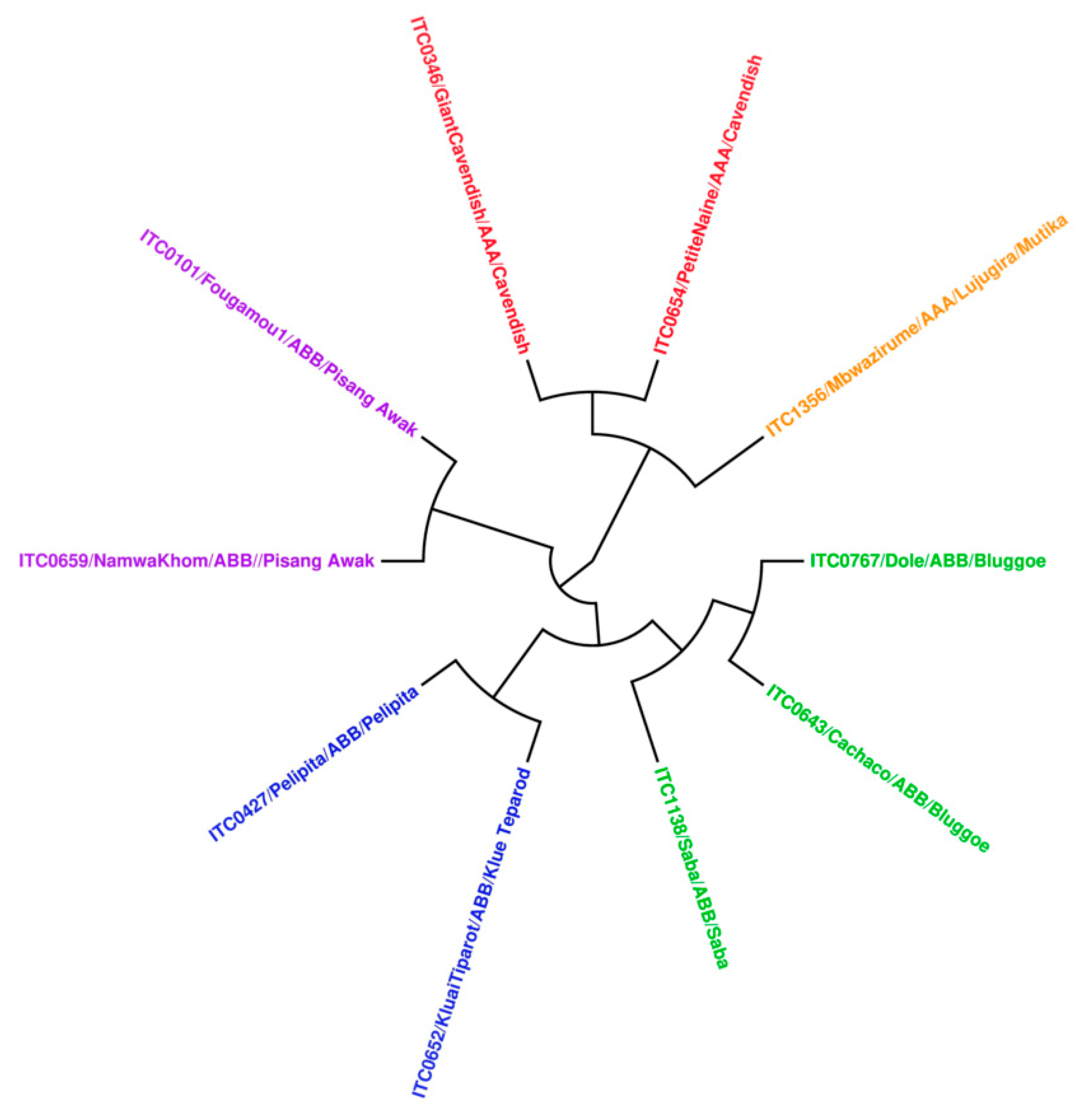
2.1. Collection of Samples
2.2. Extraction Preparation
2.3. Antiviral Test
2.4. Thin-Layer Chromatography (TLC) and Determination of Total Phenolic Content (TPC)
2.5. Statistical Analysis
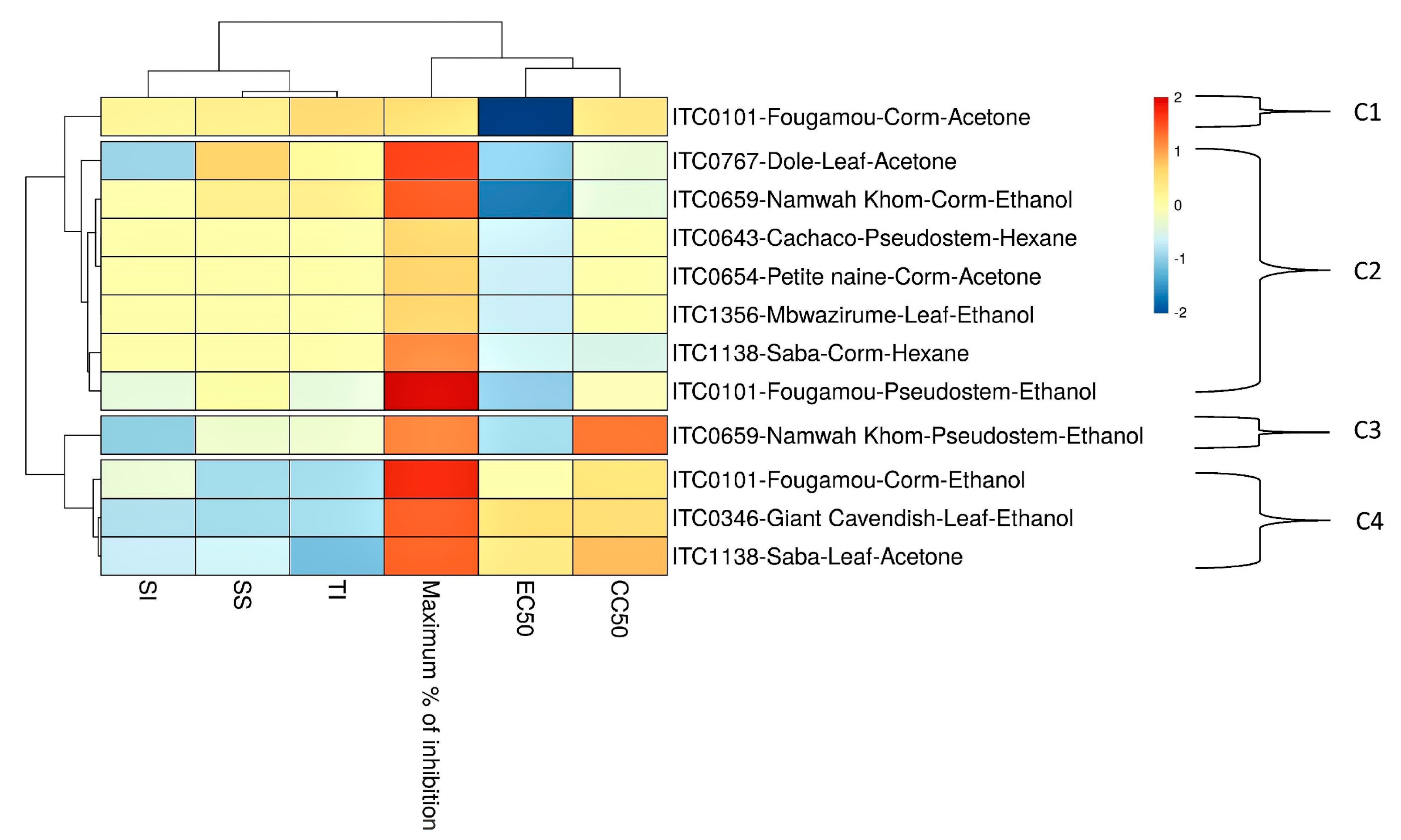
3. Results
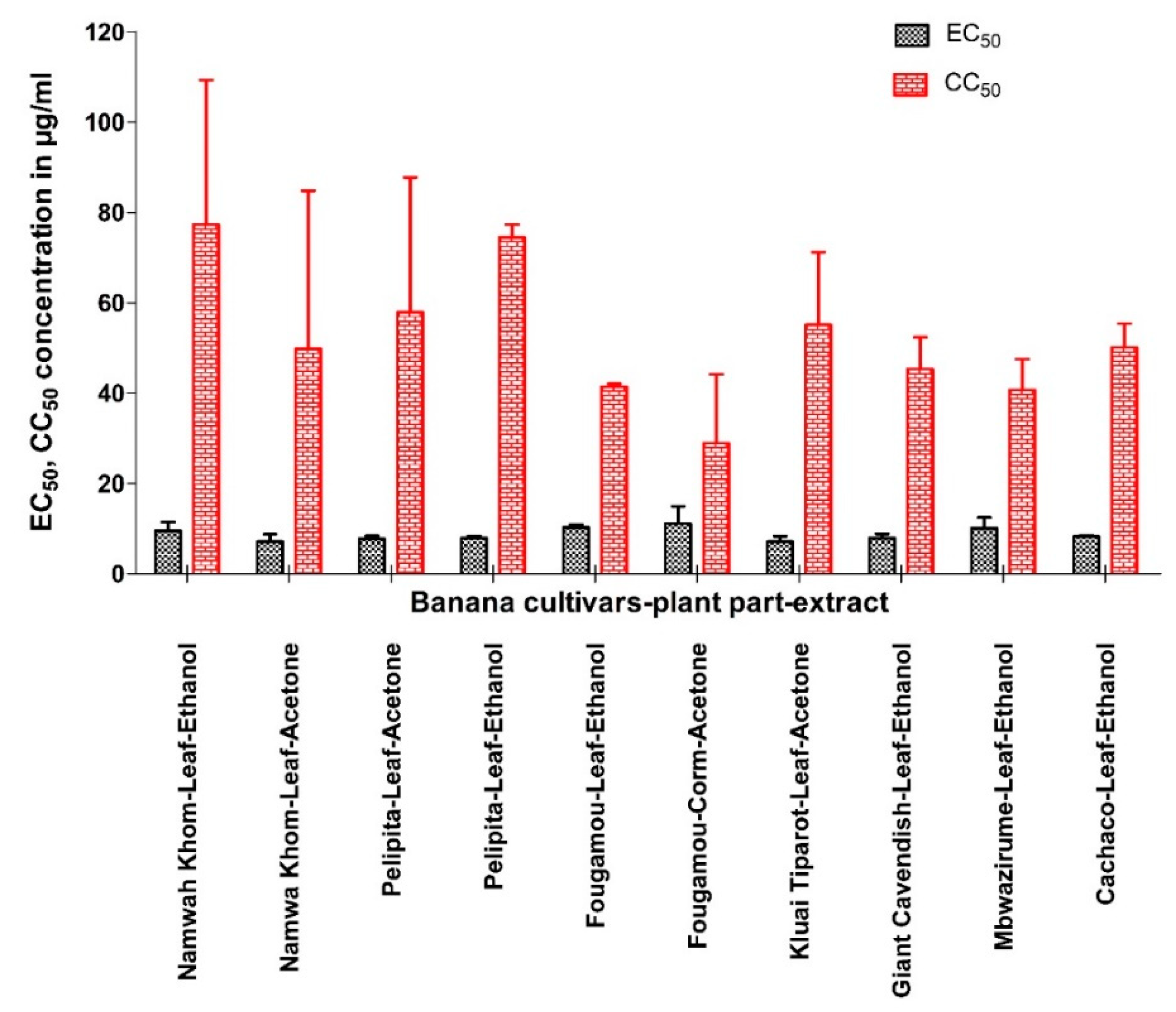
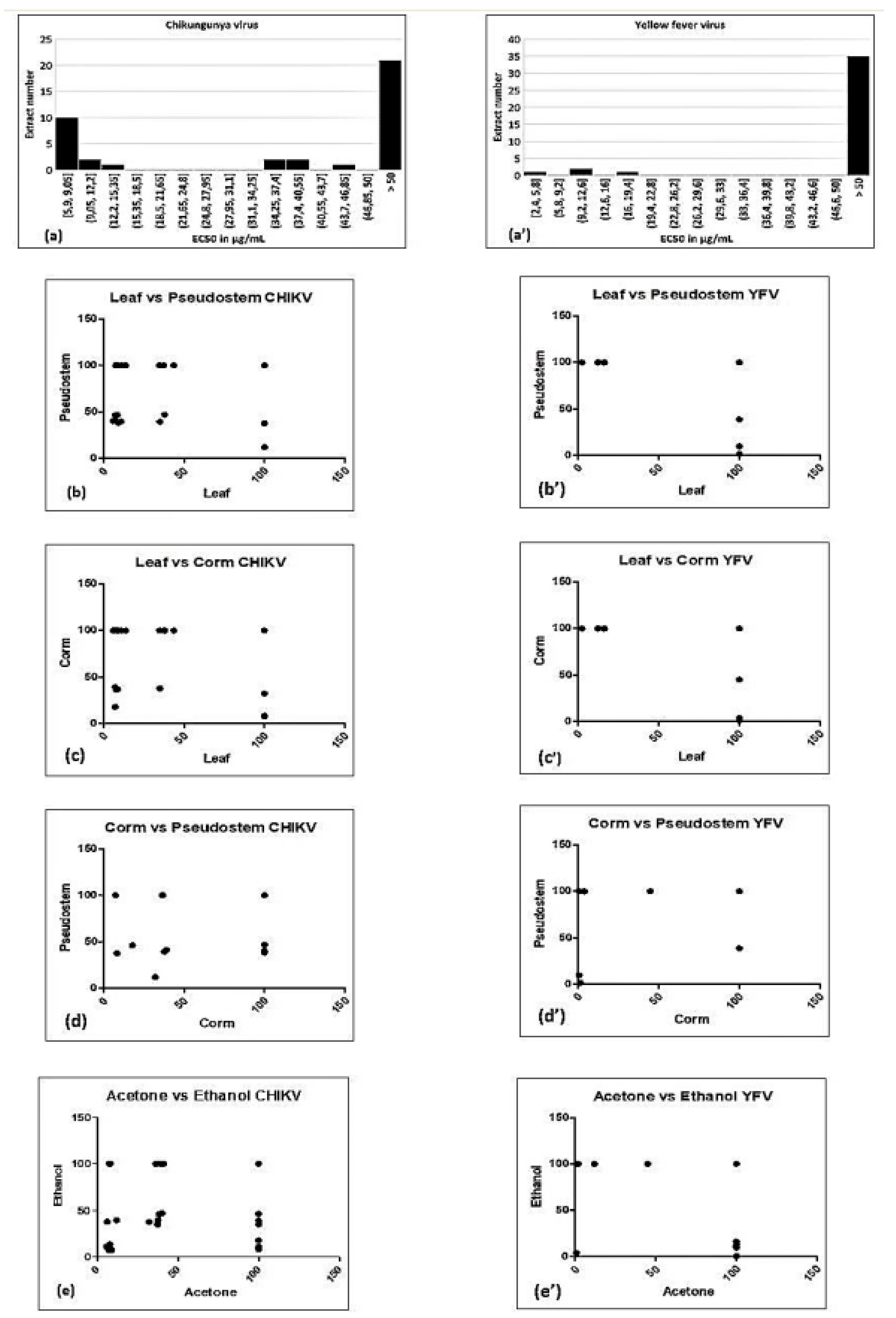
3.1. Activity of Extracts against CHIKV
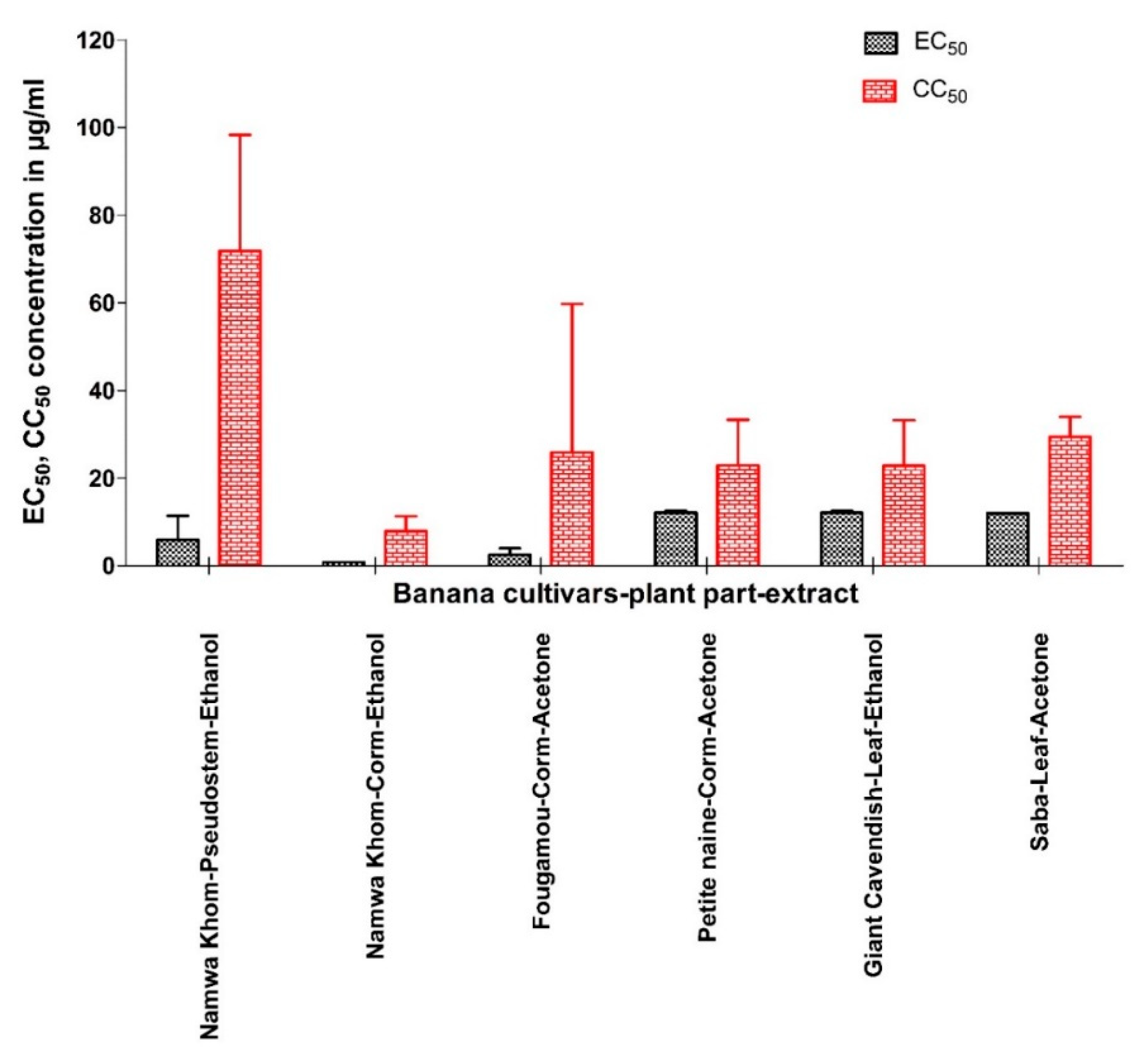
3.2. Activity of Extracts against YFV
3.3. TLC and TPC
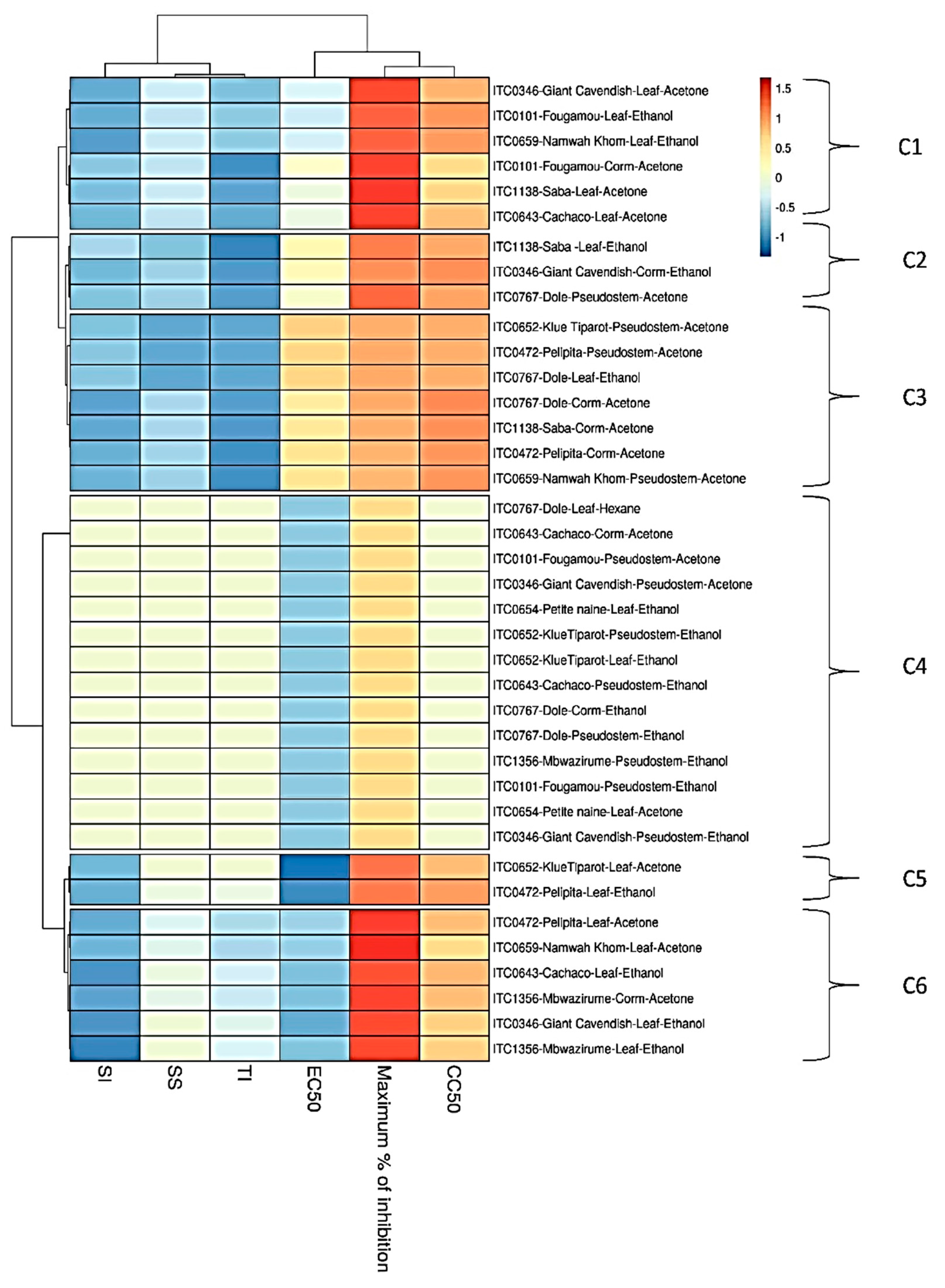
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. Banana Market Review Preliminary Results for 2018, Rome, Italy, 2018. Available online: www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Bananas/Documents/Banana_Market_Review_Prelim_Results_2018.pdf (accessed on 13 May 2020).
- Heslop-Harrison, J.S.; Schwarzacher, T. Domestication, Genomics and the Future for Banana. Ann. Bot. 2007, 100, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swennen, R.; Rosales, F. Bananas. In Encyclopedia of Agricultural Science; Academic Press: New York, NY, USA, 1994. [Google Scholar]
- Wuyts, N.; Swennen, R.; De Waele, D. Activity of phenylalanine ammonia-lyase, peroxidase and polyphenol oxidase in roots of banana (Musa acuminata AAA, cvs Grande Naine and Yangambi km5) before and after infection with Radopholus similis. Nematology 2006, 8, 201–209. [Google Scholar] [CrossRef]
- Amah, D.; Alamu, E.; Adesokan, M.; van Biljon, A.; Maziya-dixon, B.; Swennen, R.; Labuschagne, M. Variability of carotenoids in a Musa germplasm collection and implications for provitamin A biofortification. Food Chem. X 2019, 2, 100024. [Google Scholar] [CrossRef]
- Amah, D.; van Biljon, A.; Brown, A.; Perkins-Veazie, P.; Swennen, R.; Labuschagne, M. Recent advances in banana (Musa spp.) biofortification to alleviate vitamin A deficiency. Crit. Rev. Food Sci. Nutr. 2018, 59, 3498–3510. [Google Scholar] [CrossRef] [Green Version]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Bioactive compounds in banana and their associated health benefits—A review. Food Chem. 2016, 206, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.; Maraschin, M. Banana (Musa spp.) from peel to pulp: Ethnopharmacology, source of bioactive compounds and its relevance for human health. J. Ethnopharmacol. 2015, 160, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Passo Tsamo, C.V.; Herent, M.-F.; Tomekpe, K.; Happi Emaga, T.; Quetin-Leclercq, J.; Rogez, H.; Larondelle, Y.; Andre, C.M. Effect of boiling on phenolic profiles determined using HPLC/ESI-LTQ-Orbitrap-MS, physico-chemical parameters of six plantain banana cultivars (Musa sp). J. Food Compos. Anal. 2015, 44, 158–169. [Google Scholar] [CrossRef]
- Panda, S.K.; Patra, N.; Sahoo, G.; Bastia, A.K.; Dutta, S.K. Anti-Diarrheal Activities of Medicinal Plants of Similipal Biosphere Reserve, Odisha, India. Int. J. Med. Aromat. Plants 2012, 2, 123–134. [Google Scholar]
- Panda, S.K.; Brahma, S.; Dutta, S.K. Selective antifungal action of crude extracts of Cassia fistula L.: A preliminary study on Candida and Aspergillus species. Malays. J. Microbiol. 2010, 6, 62–68. [Google Scholar]
- Goel, R.K.; Sairam, K. Anti-ulcer drugs from indigenous sources with emphasis on Musa sapientum, tamrabhasma, Asparagus racemosus and Zingiber officinale. Indian J. Pharmacol. 2002, 34, 100–110. [Google Scholar]
- Gomathy, R.; Vijayalekshmi, N.R.; Kurup, P.A. Hypolipidemic principle of the inflorescence stalk of plantain (Musa sapientum). J. Biosci. 1989, 14, 301–309. [Google Scholar] [CrossRef]
- Gomathy, R.; Vijayalekshmi, N.R.; Kurup, P.A. Hypoglycemic action of the pectin present in the juice of the inflorescence stalk of plantain (Musa sapientum)—Mechanism of action. J. Biosci. 1990, 15, 297–303. [Google Scholar] [CrossRef]
- Agarwal, P.K.; Singh, A.; Gaurav, K.; Goel, S.; Khanna, H.D.; Goel, R.K. Evaluation of wound healing activity of extracts of plantain banana (Musa sapientum var. paradisiaca) in rats. Indian J. Exp Biol. 2009, 47, 32–40. [Google Scholar] [PubMed]
- Swanson, M.D.; Winter, H.C.; Goldstein, I.J.; Markovitz, D.M. A lectin isolated from bananas is a potent inhibitor of HIV replication. J. Biol. Chem. 2010, 285, 8646–8655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, M.D.; Boudreaux, D.M.; Salmon, L.; Chugh, J.; Winter, H.C.; Meagher, J.L.; André, S.; Murphy, P.V.; Oscarson, S.; Roy, R.; et al. Engineering a Therapeutic Lectin by Uncoupling Mitogenicity from Antiviral Activity. Cell 2015, 163, 746–758. [Google Scholar] [CrossRef] [Green Version]
- Risterucci, A.M.; Hippolyte, I.; Perrier, X.; Xia, L.; Caig, V.; Evers, M.; Huttner, E.; Kilian, A.; Glaszmann, J.C. Development and assessment of Diversity Arrays Technology for high-throughput DNA analyses in Musa. Theor. Appl. Genet. 2009, 119, 1093–1103. [Google Scholar] [CrossRef]
- Panda, S.K. Ethno-medicinal uses and screening of plants for antibacterial activity from Similipal Biosphere Reserve, Odisha, India. J. Ethnopharmacol. 2014, 151, 158–175. [Google Scholar] [CrossRef]
- Panda, S.K.; Padhi, L.; Leyssen, P.; Liu, M.; Neyts, J.; Luyten, W. Antimicrobial, anthelmintic, and antiviral activity of plants traditionally used for treating infectious disease in the Similipal Biosphere Reserve, Odisha, India. Front. Pharmacol. 2017, 8, 658. [Google Scholar] [CrossRef] [Green Version]
- Ledoux, A.; Cao, M.; Jansen, O.; Mamede, L.; Campos, P.-E.; Payet, B.; Clerc, P.; Grondin, I.; Girard-Valenciennes, E.; Hermann, T.; et al. Antiplasmodial, anti-chikungunya virus and antioxidant activities of 64 endemic plants from the Mascarene Islands. Int. J. Antimicrob. Agents 2018, 52, 622–628. [Google Scholar] [CrossRef]
- Saudi, M.; Zmurko, J.; Kaptein, S.; Rozenski, J.; Neyts, J.; Van Aerschot, A. Synthesis and evaluation of imidazole-4,5-and pyrazine-2,3-dicarboxamides targeting dengue and yellow fever virus. Eur. J. Med. Chem. 2014, 87, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Mohanta, Y.K.; Padhi, L.; Luyten, W. Antimicrobial activity of select edible plants from Odisha, India against food-borne pathogens. LWT 2019, 113, 108246. [Google Scholar] [CrossRef]
- Jouneghani, R.S.; Castro, A.H.F.; Panda, S.K.; Swennen, R.; Luyten, W. Antimicrobial Activity of Selected Banana Cultivars Against Important Human Pathogens, Including Candida Biofilm. Foods 2020, 9, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michener, C.D.; Sokal, R.R. A Quantitative Approach to A Problem in Classification. Evolution 1957, 11, 130–162. [Google Scholar] [CrossRef]
- Christelová, P.; De Langhe, E.; Hřibová, E.; Čížková, J.; Sardos, J.; Hušáková, M.; Van den houwe, I.; Sutanto, A.; Kepler, A.K.; Swennen, R.; et al. Molecular and cytological characterization of the global Musa germplasm collection provides insights into the treasure of banana diversity. Biodivers. Conserv. 2017, 26, 801–824. [Google Scholar] [CrossRef]
- Patil, H.M.; Bhaskar, V.V. Medicinal knowledge system of tribals of Nandurbar district, Maharashtra. Indian J. Tradit. Knowl. 2006, 5, 327–330. [Google Scholar]
- Kumar, V. Ethno-medicinal plants in five forest ranges in Dang’s district, South Gujarat, India. Ann. Pharm. Pharm. Sci. 2015, 6, 33–42. [Google Scholar]
- Dutta, N.K.; Dave, K.H.; Desal, S.M.; Mhasalkar, M.Y. Anti-variola and anti-vaccinia principles from seeds of Banakadali (Ensete superbum Cheesm, Musaceae). Indian J. Med. Res. 1968, 56, 735–741. [Google Scholar]
- Corona, M.A.; Gómez-Patiño, M.B.; de Flores, M.J.; Ruiz, L.A.; Martinez, B.M.; Arrieta-Baez, D. An Integrated Analysis of the Musa Paradisiaca Peel, Using UHPLC-ESI, FT-IR and Confocal Microscopy Techniques. Ann. Chromatogr. Sep. Tech. 2015, 1, 1005. [Google Scholar]
- Someya, S.; Yoshiki, Y.; Okubo, K. Antioxidant compounds from bananas (Musa Cavendish). Food Chem. 2002, 79, 351–354. [Google Scholar] [CrossRef]
- Medini, F.; Megdiche, W.; Mshvildadze, V.; Pichette, A.; Legault, J.; St-Gelais, A.; Ksouri, R. Antiviral-guided fractionation and isolation of phenolic compounds from Limonium densiflorum hydroalcoholic extract. Comptes Rendus Chim. 2016, 19, 726–732. [Google Scholar] [CrossRef]
- Petrescu, A.M.; Paunescu, V.; Ilia, G. The antiviral activity and cytotoxicity of 15 natural phenolic compounds with previously demonstrated antifungal activity. J. Environ. Sci. Health. Part B Pestic. Food Contam. Agric. Wastes 2019, 54, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Aquino, C.F.; Salomão, L.C.C.; Ribeiro, S.M.R.; De Siqueira, D.L.; Cecon, P.R. Carboidratos, compostos fenólicos e atividade antioxidante em polpas e cascas de 15 cultivares de bananeira. Rev. Bras. Frutic. 2016, 38. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Das, R.; Leyssen, P.; Neyts, J.; Luyten, W. Assessing medicinal plants traditionally used in the Chirang Reserve Forest, Northeast India for antimicrobial activity. J. Ethnopharmacol. 2018, 225, 220–233. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ITC | Cultivar | Genome | Subgroup | DArT |
|---|---|---|---|---|
| ITC0767 | Dole | ABB | Bluggoe | Other ABB |
| ITC0643 | Cachaco | ABB | Bluggoe | Musa balbisiana |
| ITC1138 | Saba | ABB | Saba | Other ABB |
| ITC0652 | Kluai Tiparot | ABB | unknown | Musa balbisiana |
| ITC0472 | Pelipita | ABB | unknown | Other ABB |
| ITC0659 | Namwah Khom | ABB | Pisang Awak | Pisang Awak |
| ITC0101 | Fougamou | ABB | Pisang Awak | Pisang Awak |
| ITC0654 * | Petite Naine | AAA | Cavendish | AAA Cavendish |
| ITC0346 | Giant Cavendish | AAA | Cavendish | AAA Cavendish |
| ITC1356 | Mbwazirume | AAA | Mutika/Lujugira | AAAh |
| Code No. | Variety | Parts Used | Extract Tested | EC50 (µg/mL) | EC90 (µg/mL) | Maximum % Inhibition | CC50 (µg/mL) | SI | SS | TI |
|---|---|---|---|---|---|---|---|---|---|---|
| BAVARIE1_001 | Saba | Leaf | Acetone | 7.88 | 12.6 | 72.2 | 23.2 | 2.94 | 5.05 | 2.36 |
| BAVARIE1_003 | Saba | Corm | Acetone | 36.3 | - | 73.3 | 100 | 2.76 | 5.13 | 2.26 |
| BAVARIE1_004 | Pelipita | Leaf | Acetone | 7.21 | - | 80.6 | 38.1 | 5.28 | 11 | 7.97 |
| BAVARIE1_005 | Pelipita | Pseudostem | Acetone | 41.2 | - | 69.9 | 69 | 1.67 | 0.026 | 0.006 |
| BAVARIE1_006 | Pelipita | Corm | Acetone | 38.9 | - | 71 | 100 | 2.57 | 4.31 | 1.77 |
| BAVARIE1_007 | Kluai Tiparot | Leaf | Acetone | 6.27 | 16 | 100 | 63.1 | 10.1 | 25.3 | 25.4 |
| BAVARIE1_008 | Kluai Tiparot | Pseudostem | Acetone | 40.3 | - | 60 | 63 | 1.57 | 0.093 | 0.018 |
| BAVARIE1_010 | Petit Naine | Leaf | Acetone | 37.5 | - | 76.8 | >100 | >2.67 | >6.4 | >2.72 |
| BAVARIE1_0013 | Cavendish | Leaf | Ethanol | 7.29 | 16.1 | 100 | 44.7 | 6.13 | 19.8 | 15.6 |
| BAVARIE1_0014 | Cavendish | Pseudostem | Ethanol | 46.2 | 89 | 97.1 | >100 | >2.16 | >7.89 | >2.64 |
| BAVARIE1_0015 | Cavendish | Corm | Ethanol | 17.8 | - | 65.4 | 63.1 | 3.55 | 4.91 | 2.7 |
| BAVARIE1_0016 | Fougamou | Leaf | Ethanol | 10.8 | - | 82.8 | 60.9 | 5.65 | 9.66 | 7.26 |
| BAVARIE1_0017 | Fougamou | Pseudostem | Ethanol | 39.7 | 67 | 100 | >100 | >2.52 | >11.6 | >4.65 |
| BAVARIE1_0019 | Mbwazirume | Leaf | Ethanol | 8.41 | 15 | 100 | 44.7 | 5.32 | 18.9 | 13.7 |
| BAVARIE1_0020 | Mbwazirume | Pseudostem | Ethanol | 39 | 67.6 | 92.2 | >100 | >2.56 | >10.3 | >4.22 |
| BAVARIE1_0022 | Dole | Leaf | Ethanol | 35 | - | 59.28 | 58.5 | 1.67 | 0.164 | 0.036 |
| BAVARIE1_0023 | Dole | Pseudostem | Ethanol | 39.4 | - | 85.4 | >100 | >2.54 | >8.76 | >3.55 |
| BAVARIE1_0024 | Dole | Corm | Ethanol | 37.8 | - | 85.8 | >100 | >2.65 | >9.3 | >3.93 |
| BAVARIE1_0025 | Cachaco | Leaf | Ethanol | 8.41 | 15.7 | 91.4 | 50.2 | 5.97 | 17 | 13.2 |
| BAVARIE1_0026 | Cachaco | Pseudostem | Ethanol | 46.5 | - | 82.5 | >100 | >2.15 | >6.07 | >2.02 |
| BAVARIE1_0028 | Namwah Khom | Leaf | Ethanol | 10.9 | >20 | 79.3 | 54.7 | 5 | 10.2 | 7.17 |
| BAVARIE1_0031 | Saba | Leaf | Ethanol | 13.7 | - | 65.2 | 44.7 | 3.27 | 2.46 | 1.26 |
| BAVARIE1_0034 | Pelipita | Leaf | Ethanol | 7.59 | 13.3 | 92.3 | 72.5 | 9.55 | 21.9 | 21.5 |
| BAVARIE1_0037 | Kluai Tiparot | Leaf | Ethanol | 37.9 | 63.7 | 92.6 | >100 | >2.64 | >11 | >4.62 |
| BAVARIE1_0038 | Kluai Tiparot | Pseudostem | Ethanol | 47 | - | 82.5 | >100 | >2.13 | >6.01 | >1.97 |
| BAVARIE1_0040 | Petit Naine | Leaf | Ethanol | 34.8 | - | 77.6 | >100 | >2.87 | >7.28 | >3.33 |
| BAVARIE1_0043 | Cavendish | Leaf | Acetone | 9.06 | 17.8 | 76.4 | 38.1 | 4.2 | 8.15 | 5.08 |
| BAVARIE1_0044 | Cavendish | Pseudostem | Acetone | 38.4 | - | 85.7 | >100 | >2.6 | >9.09 | >3.78 |
| BAVARIE1_0047 | Fougamou | Pseudostem | Acetone | 37.7 | - | 80.6 | >100 | >2.65 | >7.93 | >3.36 |
| BAVARIE1_0048 | Fougamou | Corm | Acetone | 8.25 | - | 68.47 | 20 | 2.43 | 3.55 | 1.37 |
| BAVARIE1_0051 | Mbwazirume | Corm | Acetone | 7.47 | 20.0 | 90 | 44.7 | 5.99 | 14.9 | 11.6 |
| BAVARIE1_0053 | Dole | Pseudostem | Acetone | 12.1 | - | 70.72 | 44.7 | 3.69 | 4.41 | 2.5 |
| BAVARIE1_0054 | Dole | Corm | Acetone | 32.2 | - | 74.8 | 100 | 3.11 | 6.1 | 3.01 |
| BAVARIE1_0055 | Cachaco | Leaf | Acetone | 8.56 | 15.9 | 72.6 | 29.9 | 3.49 | 5.64 | 3.06 |
| BAVARIE1_0057 | Cachaco | Corm | Acetone | 36.7 | - | 66.7 | >100 | >2.73 | >6.69 | >2.92 |
| BAVARIE1_0058 | Namwah Khom | Leaf | Acetone | 5.94 | 10.4 | 79.86 | 27.6 | 4.65 | 9.67 | 6.45 |
| BAVARIE1_0059 | Namwah Khom | Pseudostem | Acetone | 40.2 | - | 69.91 | 100 | 2.49 | 3.95 | 1.56 |
| BAVARIE1_0100 | Dole | Leaf | Hexane | 43.7 | - | 77.38 | >100 | >2.29 | >5.87 | >2.11 |
| Positive control | Favipiravir | - | - | 2.65 | - | - | >100 | >37.7 | - | - |
| Code No. | Variety | Parts Used | Extract Tested | EC50 (µg/mL) | EC90 (µg/mL) | Maximum % Inhibition | CC50 (µg/mL) | SI | SS | TI |
|---|---|---|---|---|---|---|---|---|---|---|
| BAVARIE1_001 | Saba | Leaf | Acetone | 12.2 | >20 | 64 | 26.2 | 2.14 | 2.27 | 0.75 |
| BAVARIE1_0012 | Petit Naine | Corm | Acetone | 44.8 | 88 | 97.57 | >100 | >2.23 | >8.29 | >2.89 |
| BAVARIE1_0013 | Cavendish | Leaf | Ethanol | 12.5 | >20 | 52.05 | 13.8 | 1.1 | 0.358 | 0.015 |
| BAVARIE1_0017 | Fougamou | Pseudostem | Ethanol | 1.55 | 2.62 | 75.27 | 5.23 | 3.37 | 6.56 | 3.46 |
| BAVARIE1_0019 | Mbwazirume | Leaf | Ethanol | 16.3 | - | 89.81 | >100 | >6.15 | >19.2 | >15.1 |
| BAVARIE1_0029 | Namwah Khom | Pseudostem | Ethanol | 9.77 | - | 89.64 | 90.6 | 9.27 | 19.2 | 18.5 |
| BAVARIE1_0030 | Namwah Khom | Corm | Ethanol | 0.57 | <0.8 | 80 | 5.77 | 9.97 | 17.1 | 17 |
| BAVARIE1_0048 | Fougamou | Corm | Acetone | 1.44 | 2.31 | 100 | 76.5 | 53.1 | 63.2 | 109 |
| BAVARIE1_0052 | Dole | Leaf | Acetone | 2.41 | 51.7 | 100 | 6.76 | 2.81 | 29.3 | 13.1 |
| BAVARIE1_0104 | Cachaco | Pseudostem | Hexane | 38.5 | 74.8 | 107 | >100 | >2.6 | >11.9 | >4.93 |
| BAVARIE1_0111 | Saba | Corm | Hexane | <0.8 | <0.8 | 57.4 | 1.12 | >1.4 | >0.54 | >0.08 |
| BAVARIE1_0118 | Fougamou | Corm | Ethanol | 3.84 | >100 | 51.31 | 8.94 | 2.33 | 0.241 | 0.088 |
| Positive control | 2′,5′-bis-O-trityl uridine | - | 1.2 μM | - | - | >100 μM | >80 | - | - | |
| Parts | Parameters | CHIKV | YFV |
|---|---|---|---|
| Leaf vs. Pseudostem | Spearman r | 0.4965 | −0.09467 |
| 95% confidence interval | 0.2096 to 0.7046 | −0.4026 to 0.2325 | |
| P value (two-tailed) | 0.0011 | 0.5612 | |
| Is the correlation significant? (alpha = 0.05) | Yes | No | |
| Leaf vs. Corm | Spearman r | 0.1872 | −0.1392 |
| 95% confidence interval | −0.1414 to 0.4787 | −0.4398 to 0.1894 | |
| P value (two-tailed) | 0.2473 | 0.3916 | |
| Is the correlation significant? (alpha = 0.05) | No | No | |
| Corm vs. Pseudostem | Spearman r | 0.3755 | 0.4228 |
| 95% confidence interval | 0.06294 to 0.6210 | 0.1188 to 0.6544 | |
| P value (two-tailed) | 0.0169 | 0.0066 | |
| Is the correlation significant? (alpha = 0.05) | Yes | Yes | |
| Acetone vs. Ethanol | Spearman r | 0.2695 | 0.0442 |
| 95% confidence interval | −0.1117 to 0.5815 | −0.2799 to 0.3593 | |
| P value (two-tailed) | 0.1499 | 0.7863 | |
| Is the correlation significant? (alpha = 0.05) | No | No |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, S.K.; Castro, A.H.F.; Jouneghani, R.S.; Leyssen, P.; Neyts, J.; Swennen, R.; Luyten, W. Antiviral and Cytotoxic Activity of Different Plant Parts of Banana (Musa spp.). Viruses 2020, 12, 549. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050549
Panda SK, Castro AHF, Jouneghani RS, Leyssen P, Neyts J, Swennen R, Luyten W. Antiviral and Cytotoxic Activity of Different Plant Parts of Banana (Musa spp.). Viruses. 2020; 12(5):549. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050549
Chicago/Turabian StylePanda, Sujogya Kumar, Ana Hortência Fonsêca Castro, Ramin Saleh Jouneghani, Pieter Leyssen, Johan Neyts, Rony Swennen, and Walter Luyten. 2020. "Antiviral and Cytotoxic Activity of Different Plant Parts of Banana (Musa spp.)" Viruses 12, no. 5: 549. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050549