Phenotypic and Genotypic Comparison of a Live-Attenuated Genotype I Japanese Encephalitis Virus SD12-F120 Strain with Its Virulent Parental SD12 Strain


 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Viruses and Cells
2.3. Attenuation of SD12 by Serial Passaging in BHK-21 Cells
2.4. Plaque Size Determination
2.5. Neuroinvasiveness and Neurovirulence Tests in Mice
2.6. Vaccination and Challenge
2.7. Detection of JEV Replication in Mouse Brains
2.8. Detection of Expression of NS1 and NS1’ in BHK-21 Cells
2.9. Viral Genome Sequencing
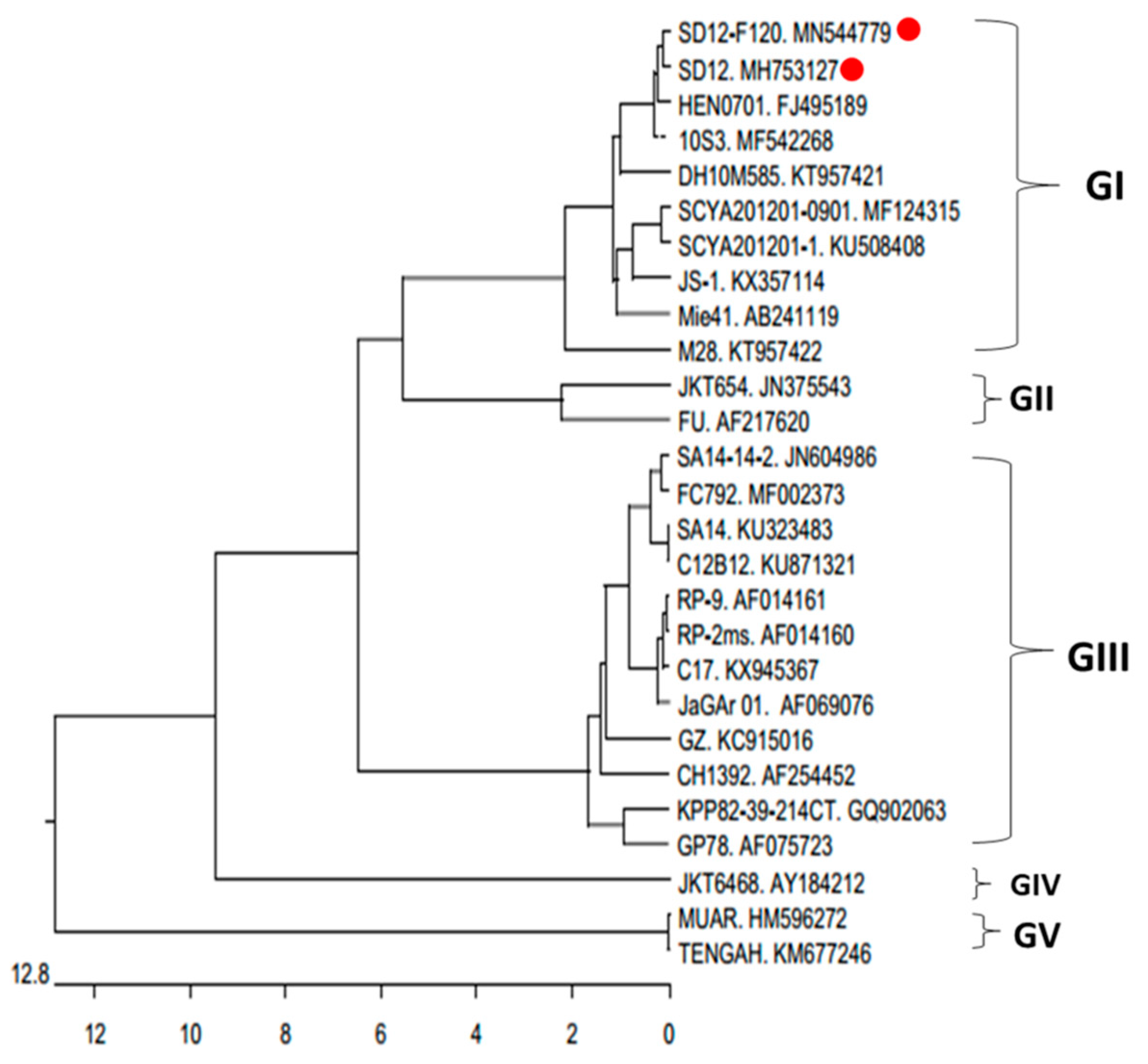
2.10. Sequence Analysis
2.11. Statistical Analysis
3. Results
3.1. Replication Phenotype and NS1ʹ Expression in BHK-21 Cells
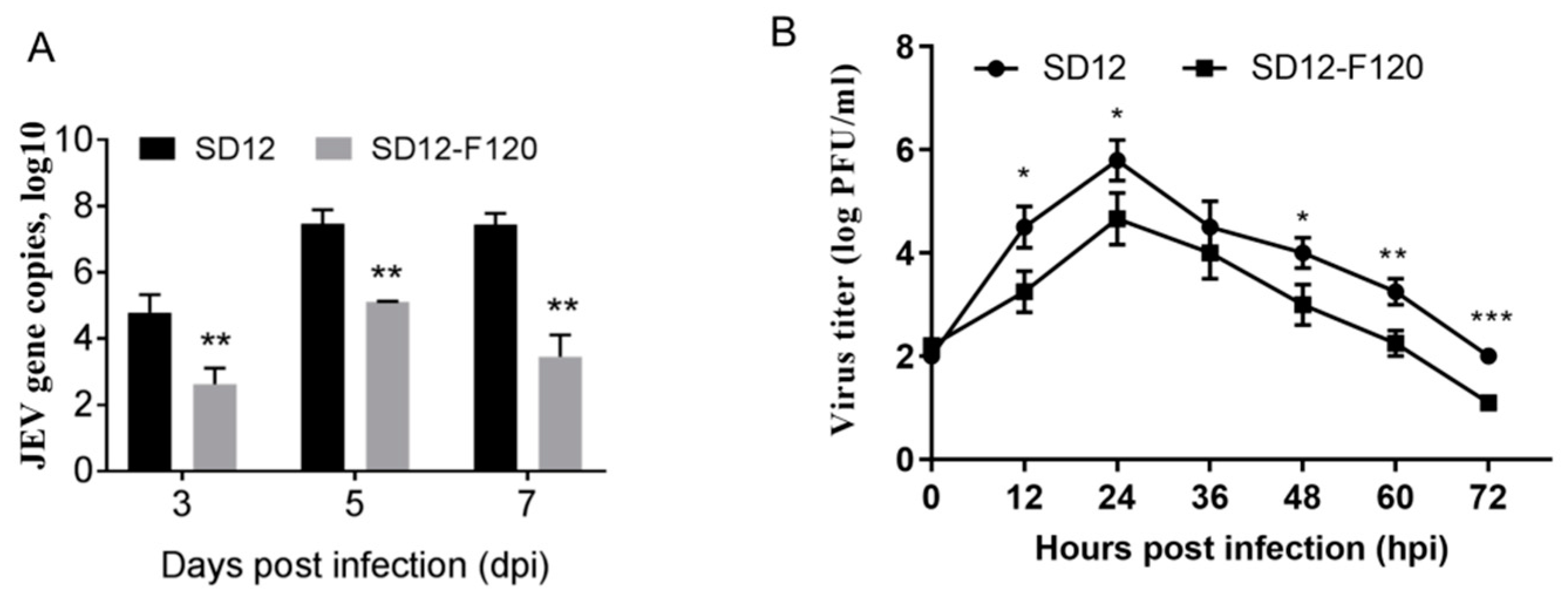
3.2. Reduced Replication Efficiency of the Attenuated SD12-F120 Strain in Mouse Brains and Mouse Primary Neuron Cells
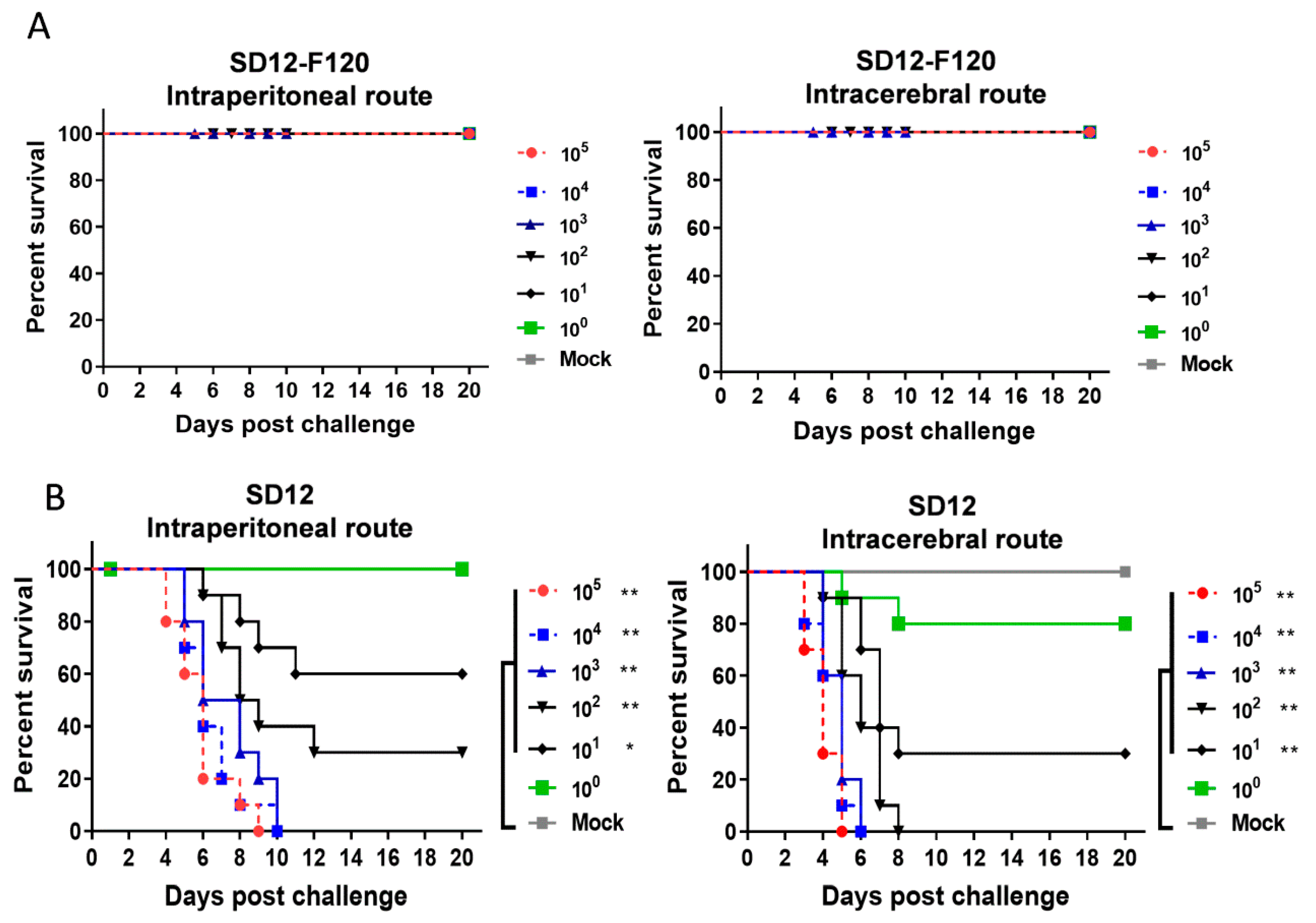
3.3. Differences in Neuroinvasiveness and Neurovirulence between the Attenuated SD12-F120 and Its Virulent Parental SD12 Strains in Mice
3.4. Protective Efficacy of the Attenuated SD12-F120 Strain against Its Virulent Parental SD12 Challenge Strain in Mice
3.5. Overview of Nucleotide and Amino Acid Variations between the Attenuated SD12-F120 and Its Virulent Parental SD12 Strains
3.6. Comparison of Amino Acid Substitutions in E Protein among Five Isogenic Attenuated and Virulent Strain Pairs
3.7. Comparison of Variations in Other Regions among Five Isogenic Attenuated and Virulent Strain Pairs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- van den Hurk, A.F.; Ritchie, S.A.; Mackenzie, J.S. Ecology and geographical expansion of Japanese encephalitis virus. Annu. Rev. Entomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis: A systematic review. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef]
- Solomon, T.; Ni, H.; Beasley, D.W.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef] [Green Version]
- Unni, S.K.; Růžek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microbes Infect. 2011, 13, 312–321. [Google Scholar] [CrossRef]
- Lindenbach, B.; Thiel, H.; Rice, C. Flaviviruses: The viruses and their replication. In Fields Virology, 5th ed.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1101–1152. [Google Scholar]
- Brinton, M.A.; Basu, M. Functions of the 3′ and 5′ genome RNA regions of members of the genus Flavivirus. Virus. Res. 2015, 206, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Kim, J.M.; Song, B.H.; Yun, S.I.; Yun, G.N.; Byun, S.J.; Lee, Y.M. Profiling of viral proteins expressed from the genomic RNA of Japanese encephalitis virus using a panel of 15 region-specific polyclonal rabbit antisera: Implications for viral gene expression. PLoS ONE 2015, 10, e0124318. [Google Scholar] [CrossRef] [Green Version]
- Bollati, M.; Alvarez, K.; Assenberg, R.; Baronti, C.; Canard, B.; Cook, S.; Coutard, B.; Decroly, E.; de Lamballerie, X.; Gould, E.A.; et al. Structure and functionality in flavivirus NS-proteins: Perspectives for drug design. Antivir. Res. 2010, 87, 125–148. [Google Scholar] [CrossRef]
- Brinton, M.A. Replication cycle and molecular biology of the West Nile virus. Viruses 2013, 6, 13–53. [Google Scholar] [CrossRef] [Green Version]
- Uchil, P.D.; Satchidanandam, V. Phylogenetic analysis of Japanese encephalitis virus: Envelope gene based analysis reveals a fifth genotype, geographic clustering, and multiple introductions of the virus into the Indian subcontinent. Am. J. Trop. Med. Hyg. 2001, 65, 242–251. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, H.L.; Yang, W.H.; Zhang, Y.Z.; Huang, L.J.; Deng, S.Z.; Sun, Y.J.; Yang, D.J.; Zhou, J.H. Molecular epidemiology of Japanese encephalitis viruses isolated in Yunnan province, 1977–2010. Zhonghua Liu Xing Bing Xue Za Zhi 2016, 37, 1519–1525. (In Chinese) [Google Scholar]
- Han, N.; Adams, J.; Fang, W.; Liu, S.Q.; Rayner, S. Investigation of the genotype III to genotype I shift in Japanese encephalitis virus and the impact on human cases. Virol. Sin. 2015, 30, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Schuh, A.J.; Ward, M.J.; Brown, A.J.; Barrett, A.D. Phylogeography of Japanese encephalitis virus: Genotype is associated with climate. PLoS Negl. Trop. Dis. 2013, 7, e2411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.Y.; Takasaki, T.; Fu, S.H.; Sun, X.H.; Zhang, H.L.; Wang, Z.X.; Hao, Z.Y.; Zhang, J.K.; Tang, Q.; Kotaki, A.; et al. Molecular epidemiological analysis of Japanese encephalitis virus in China. J. Gen. Virol. 2007, 88, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.L.; Liu, H.; Wang, H.Y.; Fu, S.H.; Liu, H.Z.; Zhang, H.L.; Li, M.H.; Gao, X.Y.; Wang, J.L.; Sun, X.H.; et al. Emergence of genotype I of Japanese encephalitis virus as the dominant genotype in Asia. J. Virol. 2011, 85, 9847–9853. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Li, C.; Di, D.; Cappelle, J.; Liu, L.; Wang, X.; Pang, L.; Xu, J.; Liu, K.; Li, B.; et al. Differential replication efficiencies between Japanese encephalitis virus genotype I and III in avian cultured cells and young domestic ducklings. PLoS Negl. Trop. Dis. 2018, 12, e0007046. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.C.; Liang, J.J.; Chen, J.M.; Lin, J.W.; Chen, Y.Y.; Su, K.H.; Lin, C.C.; Tu, W.C.; Chiou, M.T.; Ou, S.C. NS2B/NS3 mutations enhance the infectivity of genotype I Japanese encephalitis virus in amplifying hosts. PLoS Pathog. 2019, 15, e1007992. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccin. Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Wang, X.; Zhang, J.; Guo, S.; Pang, L.; Shi, K.; Liu, K.; Shao, D.; Qiu, Y.; Liu, L.; et al. Partial cross-protection between Japanese encephalitis virus genotype I and III in mice. PLoS Negl. Trop. Dis. 2019, 13, e0007601. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yu, Y.; Li, M.; Liang, G.; Wang, H.; Jia, L.; Dong, G. Study on the protective efficacy of SA14-14-2 attenuated Japanese encephalitis against different JE virus isolates circulating in China. Vaccine 2011, 29, 2127–2130. [Google Scholar] [CrossRef]
- Sarkar, A.; Banik, A.; Pathak, B.K.; Mukhopadhyay, S.K.; Chatterjee, S. Envelope protein gene based molecular characterization of Japanese encephalitis virus clinical isolates from West Bengal, India: A comparative approach with respect to SA14-14-2 live attenuated vaccine strain. BMC Infect. Dis. 2013, 13, 368. [Google Scholar] [CrossRef]
- Zhang, J.S.; Zhao, Q.M.; Guo, X.F.; Zuo, S.Q.; Cheng, J.X.; Jia, N.; Wu, C.; Dai, P.F.; Zhao, J.Y. Isolation and genetic characteristics of human genotype 1 Japanese encephalitis virus, China, 2009. PLoS ONE 2011, 6, e16418. [Google Scholar] [CrossRef]
- Halstead, S.B.; Thomas, S.J. New Japanese encephalitis vaccines: Alternatives to production in mouse brain. Expert Rev. Vaccines 2011, 10, 355–364. [Google Scholar] [CrossRef]
- Qian, P.; Zhi, X.; Wang, B.; Zhang, H.; Chen, H.; Li, X. Construction and immune efficacy of recombinant pseudorabies virus expressing PrM-E proteins of Japanese encephalitis virus genotype I. Virol. J. 2015, 12, 214. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.C.; Chen, J.M.; Lin, J.W.; Chen, Y.Y.; Wu, G.H.; Su, K.H.; Chiou, M.T.; Wu, S.R.; Yin, J.H.; Liao, J.W.; et al. Genotype I of Japanese Encephalitis Virus Virus-like Particles Elicit Sterilizing Immunity against Genotype I and III Viral Challenge in Swine. Sci. Rep. 2018, 8, 7481. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y. Phenotypic and genotypic characteristics of Japanese encephalitis attenuated live vaccine virus SA14-14-2 and their stabilities. Vaccine 2010, 28, 3635–3641. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Song, B.H.; Kim, J.K.; Yun, G.N.; Lee, E.Y.; Li, L.; Kuhn, R.J.; Rossmann, M.G.; Morrey, J.D.; Lee, Y.M. A molecularly cloned, live-attenuated japanese encephalitis vaccine SA14-14-2 virus: A conserved single amino acid in the ij Hairpin of the Viral E glycoprotein determines neurovirulence in mice. PLoS Pathog. 2014, 10, e1004290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.I.; Song, B.H.; Polejaeva, I.A.; Davies, C.J.; White, K.L.; Lee, Y.M. Comparison of the live-attenuated Japanese encephalitis vaccine SA14-14-2 strain with its pre-attenuated virulent parent SA14 strain: Similarities and differences in vitro and in vivo. J. Gen. Virol. 2016, 97, 2575–2591. [Google Scholar] [CrossRef]
- Yang, J.; Yang, H.; Li, Z.; Wang, W.; Lin, H.; Liu, L.; Ni, Q.; Liu, X.; Zeng, X.; Wu, Y.; et al. Envelope Protein Mutations L107F and E138K Are Important for Neurovirulence Attenuation for Japanese Encephalitis Virus SA14-14-2 Strain. Viruses 2017, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Wang, X.; Cui, G.; Pang, L.; Xu, J.; Li, C.; Zhang, J.; Liu, K.; Li, B.; Shao, D.; et al. Possible pathogenicity of Japanese encephalitis virus in newly hatched domestic ducklings. Vet. Microbiol. 2018, 227, 8–11. [Google Scholar] [CrossRef]
- Zheng, X.; Zheng, H.; Tong, W.; Li, G.; Wang, T.; Li, L.; Gao, F.; Shan, T.; Yu, H.; Zhou, Y.; et al. Acidity/Alkalinity of Japanese Encephalitis Virus E Protein Residue 138 Alters Neurovirulence in Mice. J. Virol. 2018, 92, e00108–e00118. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Yuan, L.; Wu, R.; Liu, H.; Wen, X.; Huang, X.; Wen, Y.; Ma, X.; Yan, Q.; Huang, Y.; Zhao, Q.; et al. Tissue tropism and molecular characterization of a Japanese encephalitis virus strain isolated from pigs in southwest China. Virus Res. 2016, 215, 55–64. [Google Scholar] [CrossRef]
- Qiu, Y.; Shen, Y.; Li, X.; Liu, Q.; Ma, Z. Polyclonal antibody to porcine p53 protein: A new tool for studying the p53 pathway in a porcine model. Biochem. Biophys. Res. Commun. 2008, 377, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Atkins, J.F. A conserved predicted pseudoknot in the NS2A-encoding sequence of West Nile and Japanese encephalitis flaviviruses suggests NS1′ may derive from ribosomal frameshifting. Virol. J. 2009, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Li, X.F.; Zhao, H.; Li, S.H.; Deng, Y.Q.; Cao, R.Y.; Song, K.Y.; Wang, H.J.; Hua, R.H.; Yu, Y.X.; et al. A single nucleotide mutation in NS2A of Japanese encephalitis-live vaccine virus (SA14-14-2) ablates NS1′ formation and contributes to attenuation. J. Gen. Virol. 2012, 93, 1959–1964. [Google Scholar] [CrossRef]
- Takamatsu, Y.; Raekiansyah, M.; Morita, K.; Hayasaka, D. NS1′ Protein Expression in the JaOArS982 Strain of Japanese Encephalitis Virus Does Not Enhance Virulence in Mice. Trop. Med. Health. 2015, 43, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Wu, R.; Feng, Y.; Zhao, Q.; Wen, X.; Huang, X.; Wen, Y.; Yan, Q.; Huang, Y.; Ma, X.; et al. Genomic changes in an attenuated genotype I Japanese encephalitis virus and comparison with virulent parental strain. Virus Genes 2018, 54, 424–431. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, R.; Zhao, Q.; Chang, Y.F.; Wen, X.; Feng, Y.; Huang, X.; Wen, Y.; Yan, Q.; Huang, Y.; et al. Mutation of I176R in the E coding region weakens Japanese encephalitis virus neurovirulence, but not its growth rate in BHK-21 cells. Arch. Virol. 2018, 163, 1351–1355. [Google Scholar] [CrossRef]
- Lin, Y.L.; Liao, C.L.; Yeh, C.T.; Chang, C.H.; Huang, Y.L.; Huang, Y.Y.; Jan, J.T.; Chin, C.; Chen, L.K. A highly attenuated strain of Japanese encephalitis virus induces a protective immune response in mice. Virus Res. 1996, 44, 45–56. [Google Scholar] [CrossRef]
- Chambers, T.J.; Droll, D.A.; Jiang, X.; Wold, W.S.; Nickells, J.A. JE Nakayama/JE SA14-14-2 virus structural region intertypic viruses: Biological properties in the mouse model of neuroinvasive disease. Virology 2007, 366, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Yun, S.I.; Song, B.H.; Hahn, Y.S.; Lee, C.H.; Oh, H.W.; Lee, Y.M. A single N-linked glycosylation site in the Japanese encephalitis virus prM protein is critical for cell type-specific prM protein biogenesis, virus particle release, and pathogenicity in mice. J. Virol. 2008, 82, 7846–7862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wispelaere, M.; Khou, C.; Frenkiel, M.P.; Desprès, P.; Pardigon, N. A Single Amino Acid Substitution in the M Protein Attenuates Japanese Encephalitis Virus in Mammalian Hosts. J. Virol. 2015, 90, 2676–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gromowski, G.D.; Firestone, C.Y.; Whitehead, S.S. Genetic Determinants of Japanese Encephalitis Virus Vaccine Strain SA14-14-2 That Govern Attenuation of Virulence in Mice. J. Virol. 2015, 89, 6328–6337. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Li, X.F.; Ye, Q.; Wang, H.J.; Deng, Y.Q.; Zhu, S.Y.; Zhang, Y.; Li, S.H.; Qin, C.F. Characterization of live-attenuated Japanese encephalitis vaccine virus SA14-14-2. Vaccine 2014, 32, 2675–2681. [Google Scholar] [CrossRef] [PubMed]
- Luca, V.C.; AbiMansour, J.; Nelson, C.A.; Fremont, D.H. Crystal structure of the Japanese encephalitis virus envelope protein. J. Virol. 2012, 86, 2337–2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, H.; Barrett, A.D. Attenuation of Japanese encephalitis virus by selection of its mouse brain membrane receptor preparation escape variants. Virology 1998, 241, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiou, S.S.; Liu, H.; Chuang, C.K.; Lin, C.C.; Chen, W.J. Fitness of Japanese encephalitis virus to Neuro-2a cells is determined by interactions of the viral envelope protein with highly sulfated glycosaminoglycans on the cell surface. J. Med. Virol. 2005, 76, 583–592. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, X.; Na, R.; Li, L.; Warkentin, E.; Witt, J.; Lu, X.; Yu, Y.; Wei, Y.; Peng, G.; et al. The structure differences of Japanese encephalitis virus SA14 and SA14-14-2 E proteins elucidate the virulence attenuation mechanism. Protein Cell 2019, 10, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Poonsiri, T.; Wright, G.S.A.; Diamond, M.S.; Turtle, L.; Solomon, T.; Antonyuk, S.V. Structural Study of the C-Terminal Domain of Nonstructural Protein 1 from Japanese Encephalitis Virus. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, M.; Sharma, N.; Singh, S.K. Flavivirus NS1: A multifaceted enigmatic viral protein. Virol. J. 2016, 13, 131. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Shi, Z.; Li, S.; Wang, X.; Qiu, Y.; Shao, D.; Wei, J.; Tong, G.; Ma, Z. Characterization of nonstructural protein 3 of a neurovirulent Japanese encephalitis virus strain isolated from a pig. Virol. J. 2011, 8, 209. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, T.; Unno, H.; Mori, Y.; Tani, H.; Moriishi, K.; Takamizawa, A.; Agoh, M.; Tsukihara, T.; Matsuura, Y. Crystal structure of the catalytic domain of Japanese encephalitis virus NS3 helicase/nucleoside triphosphatase at a resolution of 1.8 A. Virology 2008, 373, 426–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmurko, J.; Neyts, J.; Dallmeier, K. Flaviviral NS4b, chameleon and jack-in-the-box roles in viral replication and pathogenesis, and a molecular target for antiviral intervention. Rev. Med. Virol. 2015, 25, 205–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Gong, P. Crystal Structure of the full-length Japanese encephalitis virus NS5 reveals a conserved methyltransferase-polymerase interface. PLoS Pathog. 2013, 9, e1003549. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Chen, Z.; Li, Y.; Zhao, Z.; He, W.; Zohaib, A.; Song, Y.; Deng, C.; Zhang, B.; Chen, H.; et al. Japanese Encephalitis Virus NS5 Inhibits Type I Interferon (IFN) Production by Blocking the Nuclear Translocation of IFN Regulatory Factor 3 and NF-κB. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Yoshii, K.; Phongphaew, W.; Muto, M.; Hirano, M.; Orba, Y.; Sawa, H.; Kariwa, H. West Nile virus capsid protein inhibits autophagy by AMP-activated protein kinase degradation in neurological disease development. PLoS Pathog. 2020, 16, e1008238. [Google Scholar] [CrossRef]
- Li, X.F.; Jiang, T.; Yu, X.D.; Deng, Y.Q.; Zhao, H.; Zhu, Q.Y.; Qin, E.D.; Qin, C.F. RNA elements within the 5′ untranslated region of the West Nile virus genome are critical for RNA synthesis and virus replication. J. Gen. Virol. 2010, 91, 1218–1223. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Position | Nucleotide Changes | Amino Acid Changes | Amino Acid Substitutions | |||
|---|---|---|---|---|---|---|---|
| Nucleotide | Amino Acid | SD12-F120 | SD12 | SD12-F120 | SD12 | ||
| PrM | 540 | --* | T | C | -- | -- | -- |
| E | 1338 | -- | G | T | -- | -- | -- |
| 1390 + 1391 | 138 | AG | GA | R | E | E138R | |
| 1505 | 176 | C | T | T | I | I176T | |
| 2055 | -- | C | T | -- | -- | -- | |
| 2460 | -- | A | G | -- | -- | -- | |
| NS1 | 2694 | -- | A | G | -- | -- | -- |
| 3011 | 177 | A | G | E | G | G177E | |
| 3132 | -- | G | A | -- | -- | -- | |
| NS2A | 3924 | -- | C | T | -- | -- | -- |
| NS2B | 4503 | -- | G | A | -- | -- | -- |
| 4513 | -- | C | T | -- | -- | -- | |
| NS3 | 4922 | 104 | C | T | P | L | L104P |
| 4974 | -- | A | G | -- | -- | -- | |
| 6006 | -- | T | C | -- | -- | -- | |
| 6191 | 527 | G | T | G | V | V527G | |
| NS4A | 6726 | -- | T | C | -- | -- | -- |
| 6735 | -- | T | C | -- | -- | -- | |
| NS4B | 7407 | 46 | C | T | A | T | T46A |
| NS5 | 7688 | 3 | G | C | P | A | A3P |
| 8238 | -- | C | T | -- | -- | -- | |
| 8439 | -- | C | T | -- | -- | -- | |
| 8506 | 276 | A | G | K | E | E276K | |
| 8571 | -- | A | C | -- | -- | -- | |
| 8715 | -- | T | C | -- | -- | -- | |
| 9810 | -- | T | C | -- | -- | -- | |
| 9840 | -- | T | C | -- | -- | -- | |
| 10005 | -- | T | C | -- | -- | -- | |
| Position | Genotype I | Genotype III | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | ||||||
| Attenuated SD12-F120 | Virulent SD12 | Attenuated SCYA201201-0901 | Virulent SCYA201201 | Attenuated 10S3 | Virulent HEN0701 | Attenuated SA14-14-2 | Virulent SA14 | Attenuated RP-2ms | Virulent RP-9 | |
| 72 | A | A | T* | A | A | A | A | A | A | A |
| 107 | L | L | L | L | L | L | F | L | L | L |
| 138 | R | E | E | E | R | E | K | E | K | E |
| 176 | T | I | R | I | I | I | G | I | I | I |
| 177 | T | T | T | T | T | T | A | T | T | T |
| 244 | E | E | E | E | E | E | G | E | E | E |
| 251 | S | S | Y | S | S | S | S | S | S | S |
| 264 | Q | Q | Q | Q | Q | Q | H | Q | Q | Q |
| 273 | E | E | K | E | E | E | E | E | E | E |
| 279 | K | K | K | K | K | K | M | K | K | K |
| 315 | A | A | A | A | A | A | V | A | A | A |
| 439 | K | K | K | K | K | K | R | K | K | K |
| Total number of substitution | 2 | 4 | 1 | 9 | 1 | |||||
| Protein | Position | Genotype I | Genotype III | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | |||||||
| Attenuated SD12-F120 | Virulent SD12 | Attenuated SCYA201201-0901 | Virulent SCYA201201 | Attenuated 10S3 | Virulent HEN0701 | Attenuated SA14-14-2 | Virulent SA14 | Attenuated RP-2ms | Virulent RP-9 | ||
| C | 66 | L | L | L | L | L | L | S | L | L | L |
| prM | 86 | H | H | R* | H | H | H | H | H | H | H |
| 109 | E | E | D | E | E | E | E | E | E | E | |
| NS1 | 100 | A | A | T | A | A | A | A | A | A | A |
| 177 | E | G | E | E | E | E | E | E | E | E | |
| 237 | E | E | E | E | G | E | E | E | E | E | |
| 269 | D | D | D | N | D | D | D | D | D | D | |
| 351 | D | D | D | D | D | D | H | D | D | D | |
| NS2B | 44 | V | V | A | V | V | V | V | V | V | V |
| 63 | E | E | E | E | E | E | D | E | E | E | |
| 65 | E | E | E | E | E | E | G | D | E | D | |
| NS3 | 59 | M | M | M | M | M | M | V | M | M | M |
| 102 | P | P | P | P | P | Q | P | P | P | P | |
| 105 | P | L | P | P | P | P | G | A | A | A | |
| 527 | G | V | G | G | G | G | G | G | G | G | |
| NS4A | 63 | E | E | E | E | D | E | E | E | E | E |
| 96 | A | A | T | A | A | A | A | A | A | A | |
| 168 | G | G | E | G | G | G | G | G | G | G | |
| 248 | F | F | L | F | F | F | F | F | F | F | |
| NS4B | 46 | A | T | T | T | T | T | T | T | T | T |
| 106 | I | I | I | I | I | I | V | I | I | I | |
| 192 | V | V | V | V | A | V | V | V | V | V | |
| NS5 | 3 | P | A | P | P | P | P | P | P | P | P |
| 277 | K | E | E | E | E | K | E | E | E | E | |
| 313 | V | V | V | V | A | V | V | V | V | V | |
| 386 | H | H | H | H | H | H | Y | H | Y | Y | |
| 623 | V | V | I | V | V | V | V | V | V | V | |
| 879 | I | I | T | I | I | I | I | I | I | I | |
| Total number of substitutions | 6 | 10 | 6 | 8 | 1 | ||||||
| Region | Position | Genotype I | Genotype III | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | Isogenic Strain Pair | |||||||
| Attenuated SD12-120 | Virulent SD12-01 | Attenuated SCYA201201-0901 | Virulent SCYA201201 | Attenuated 10S3 | Virulent HEN0701 | Attenuated SA14-14-2 | Virulent SA14 | Attenuated RP-2ms | Virulent RP-9 | ||
| 5′-UTR | 39 | U | U | U | U | U | U | A* | U | U | U |
| 43 | A | A | G | A | A | A | A | A | A | A | |
| 52 | G | G | A | G | G | G | G | G | G | G | |
| 3′-UTR | 10408 | C | C | U | C | -- | C | -- | -- | -- | -- |
| 10428 | U | U | U | U | U | U | C | U | U | U | |
| 10679 | C | C | U | C | C | C | C | C | C | C | |
| 10705 | C | C | C | C | T | C | C | C | C | C | |
| 10731 | A | A | G | A | A | A | A | A | A | A | |
| 10960 | G | G | G | G | A | G | G | G | G | G | |
| Total number of substitutions | 0 | 5 | 2 | 1 | 0 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anwar, M.N.; Wang, X.; Hameed, M.; Wahaab, A.; Li, C.; Sharma, M.; Pang, L.; Malik, M.I.; Liu, K.; Li, B.; et al. Phenotypic and Genotypic Comparison of a Live-Attenuated Genotype I Japanese Encephalitis Virus SD12-F120 Strain with Its Virulent Parental SD12 Strain. Viruses 2020, 12, 552. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050552
Anwar MN, Wang X, Hameed M, Wahaab A, Li C, Sharma M, Pang L, Malik MI, Liu K, Li B, et al. Phenotypic and Genotypic Comparison of a Live-Attenuated Genotype I Japanese Encephalitis Virus SD12-F120 Strain with Its Virulent Parental SD12 Strain. Viruses. 2020; 12(5):552. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050552
Chicago/Turabian StyleAnwar, Muhammad Naveed, Xin Wang, Muddassar Hameed, Abdul Wahaab, Chenxi Li, Mona Sharma, Linlin Pang, Muhammad Irfan Malik, Ke Liu, Beibei Li, and et al. 2020. "Phenotypic and Genotypic Comparison of a Live-Attenuated Genotype I Japanese Encephalitis Virus SD12-F120 Strain with Its Virulent Parental SD12 Strain" Viruses 12, no. 5: 552. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050552