Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Species
2.2. Phage Cocktails
2.3. Bacterial Identification
2.4. Mixed-Species Planktonic Cultures and Phage Cocktail Treatment
2.5. In Vitro Mixed-Species Biofilm Development and Phage Cocktail Treatment
2.6. Viable Bacterial Cell Count
2.7. Statistical Analysis
3. Results
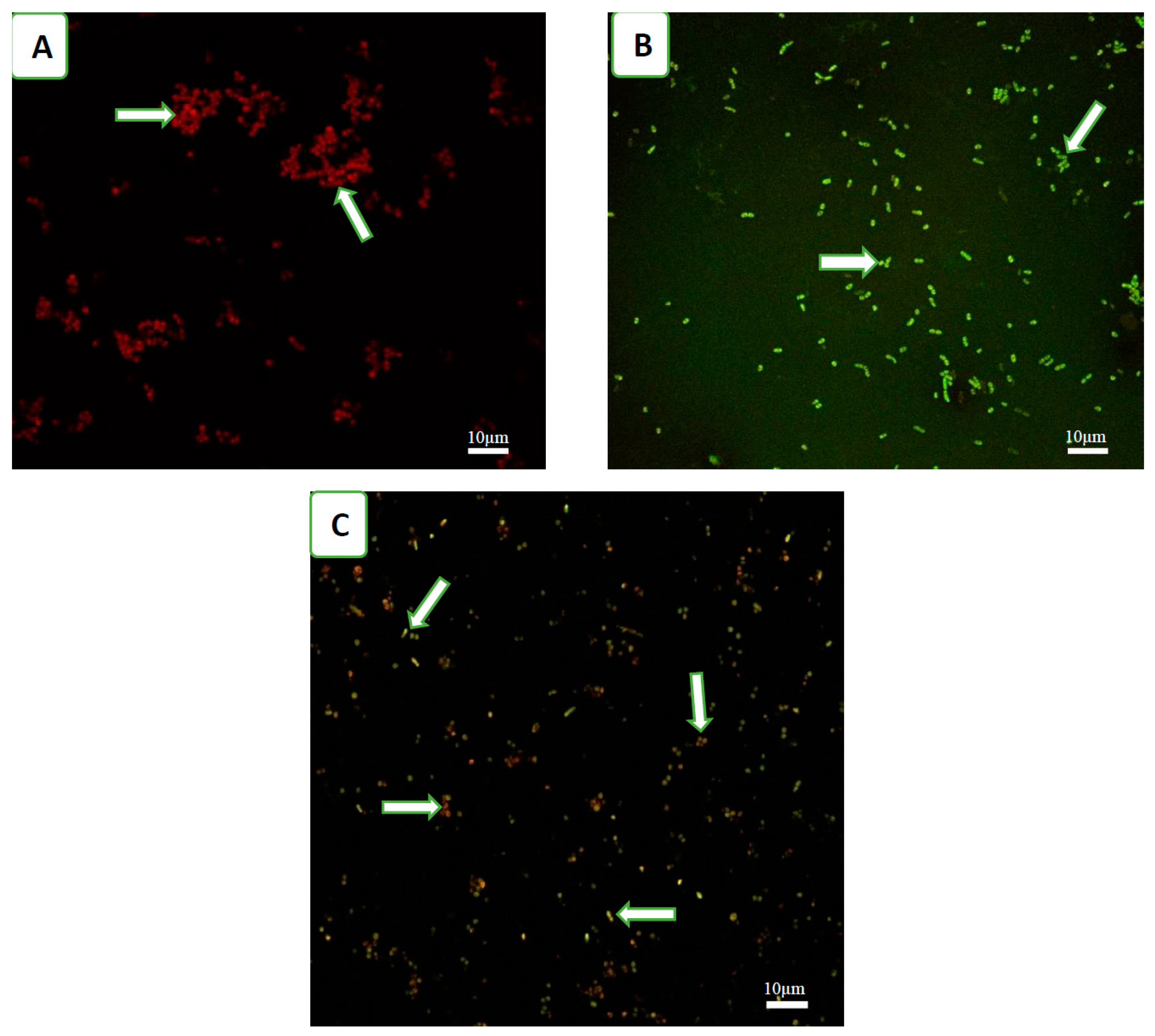
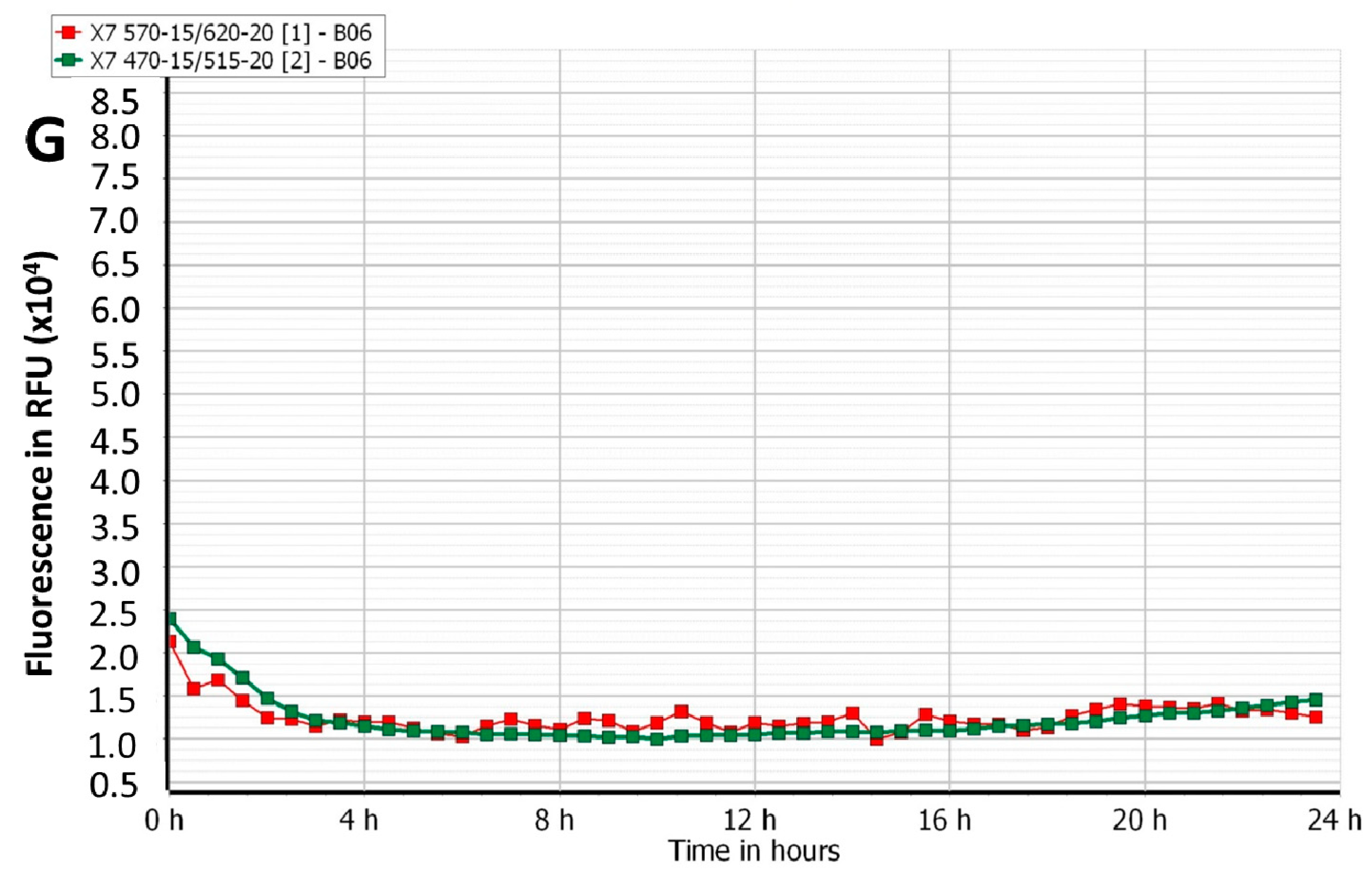
3.1. Effect of Phage Cocktails on Fluorescently Labelled Mixed-Species Planktonic Cultures
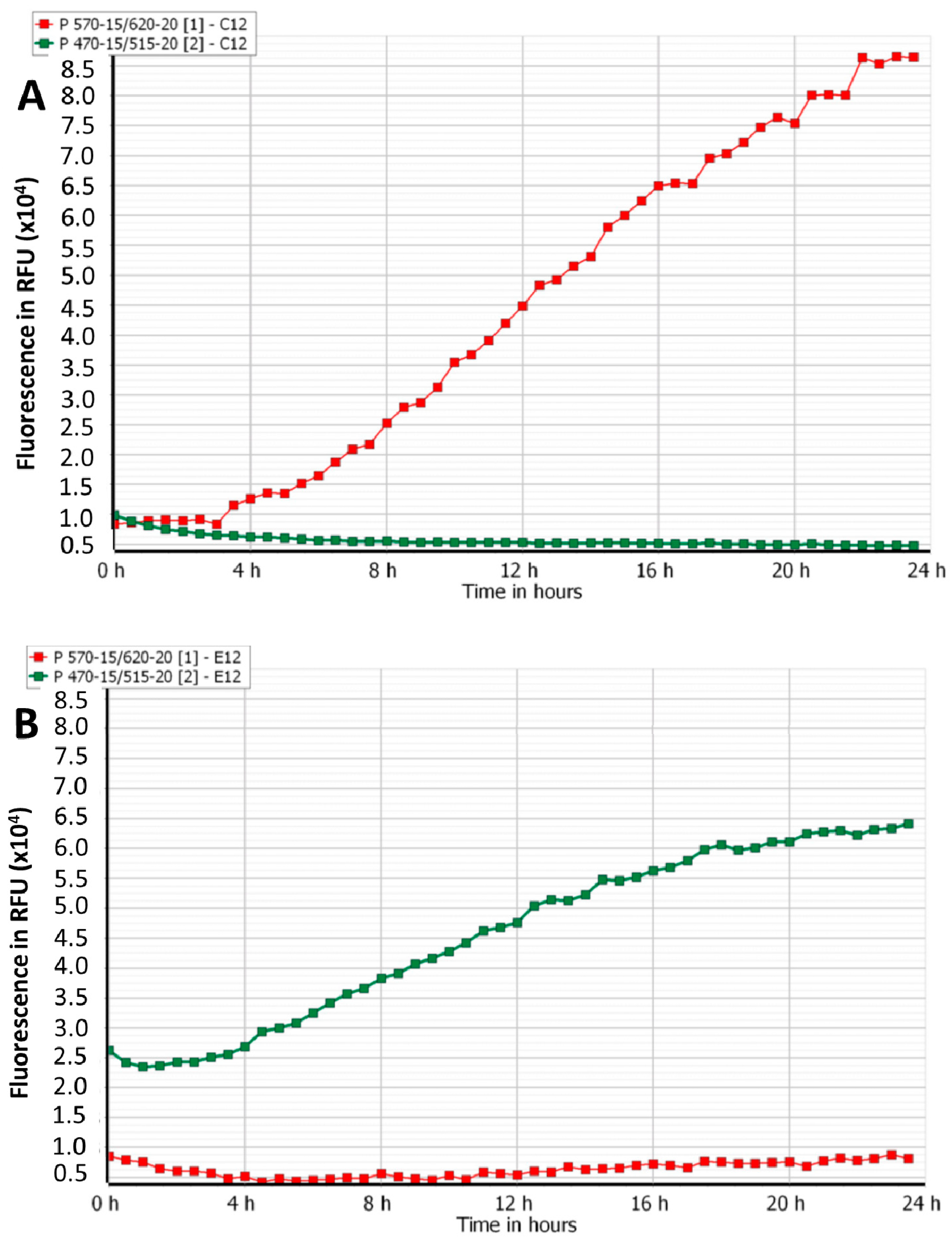
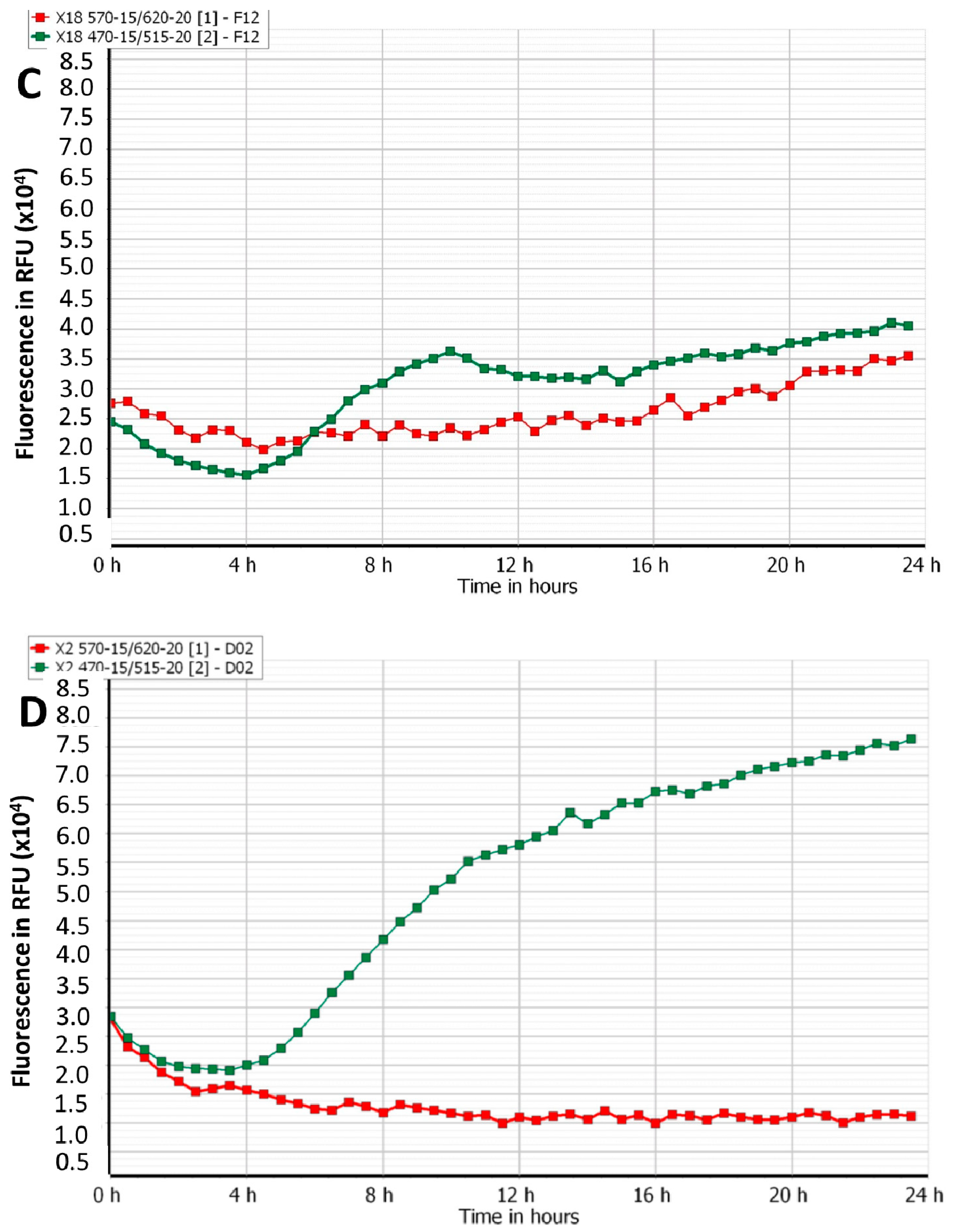
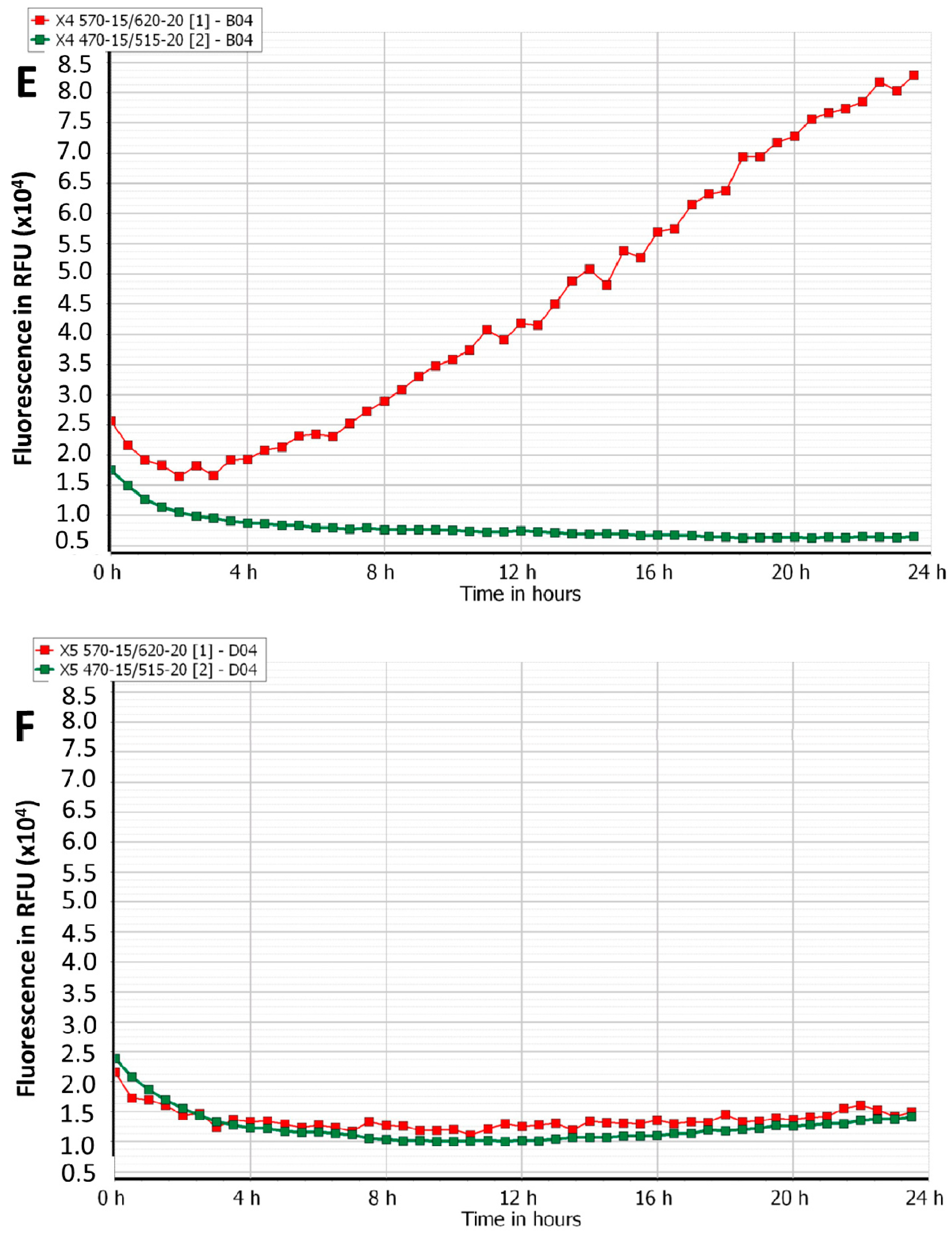
3.2. Efficacy of Phage Cocktails on Laboratory and Clinical Isolates Mixed-Species Planktonic Cultures
3.3. Effect of Phage Cocktails Treatment on Mixed-Species Biofilms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound microbiology and associated approaches to wound management. Clin. Microbiol. Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Citron, D.M.; Goldstein, E.J.; Merriam, C.V.; Lipsky, B.A.; Abramson, M.A. Bacteriology of moderate-to-severe diabetic foot infections and in vitro activity of antimicrobial agents. J. Clin. Microbiol. 2007, 45, 2819–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noor, S.; Borse, A.G.; Ozair, M.; Raghav, A.; Parwez, I.; Ahmad, J. Inflammatory markers as risk factors for infection with multidrug-resistant microbes in diabetic foot subjects. Foot 2017, 32, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Gjødsbøl, K.; Christensen, J.J.; Karlsmark, T.; Jørgensen, B.; Klein, B.M.; Krogfelt, K.A. Multiple bacterial species reside in chronic wounds: A longitudinal study. Int. Wound J. 2006, 3, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, U.; Parameswaran, S.; Armstrong, A.; Burgueno-Vega, D.; Griswold, J.; Dissanaike, S.; Rumbaugh, K.P. Prevalence of multiple antibiotic resistant infections in diabetic versus nondiabetic wounds. J. Pathog. 2014, 2014, 173053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, S.M.; Westh, H.; Danielsen, L.; Rosdahl, V.T. Bacterial colonization and healing of venous leg ulcers. Apmis 1996, 104, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Fazli, M.; Bjarnsholt, T.; Kirketerp-Moller, K.; Jorgensen, B.; Andersen, A.S.; Krogfelt, K.A.; Givskov, M.; Tolker-Nielsen, T. Nonrandom distribution of Pseudomonas aeruginosa and Staphylococcus aureus in chronic wounds. J. Clin. Microbiol. 2009, 47, 4084–4089. [Google Scholar] [CrossRef] [Green Version]
- DeLeon, S.; Clinton, A.; Fowler, H.; Everett, J.; Horswill, A.R.; Rumbaugh, K.P. Synergistic interactions of Pseudomonas aeruginosa and Staphylococcus aureus in an in vitro wound model. Infect. Immun. 2014, 82, 4718–4728. [Google Scholar] [CrossRef] [Green Version]
- Pastar, I.; Nusbaum, A.G.; Gil, J.; Patel, S.B.; Chen, J.; Valdes, J.; Stojadinovic, O.; Plano, L.R.; Tomic-Canic, M.; Davis, S.C. Interactions of methicillin resistant Staphylococcus aureus USA300 and Pseudomonas aeruginosa in polymicrobial wound infection. PLoS ONE 2013, 8, e56846. [Google Scholar] [CrossRef]
- Tay, W.H.; Chong, K.K.; Kline, K.A. Polymicrobial-Host Interactions during Infection. J. Mol. Biol. 2016, 428, 3355–3371. [Google Scholar] [CrossRef]
- Dalton, T.; Dowd, S.E.; Wolcott, R.D.; Sun, Y.; Watters, C.; Griswold, J.A.; Rumbaugh, K.P. An in vivo polymicrobial biofilm wound infection model to study interspecies interactions. PLoS ONE 2011, 6, e27317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S.T. Phage therapy in clinical practice: Treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Geredew Kifelew, L.; Mitchell, J.G.; Speck, P. Mini-review: Efficacy of lytic bacteriophages on multispecies biofilms. Biofouling 2019, 35, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Bacteriophage lysis: Mechanism and regulation. Microbiol. Rev. 1992, 56, 430–481. [Google Scholar] [CrossRef] [PubMed]
- Wittebole, X.; De Roock, S.; Opal, S.M. A historical overview of bacteriophage therapy as an alternative to antibiotics for the treatment of bacterial pathogens. Virulence 2014, 5, 226–235. [Google Scholar] [CrossRef]
- Donlan, R.M. Preventing biofilms of clinically relevant organisms using bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Torres-Barcelo, C.; Hochberg, M.E. Evolutionary Rationale for Phages as Complements of Antibiotics. Trends Microbiol. 2016, 24, 249–256. [Google Scholar] [CrossRef]
- Kirby, A.E. Synergistic action of gentamicin and bacteriophage in a continuous culture population of Staphylococcus aureus. PLoS ONE 2012, 7, e51017. [Google Scholar] [CrossRef] [Green Version]
- Woods, P.W.; Haynes, Z.M.; Mina, E.G.; Marques, C.N.H. Maintenance of S. aureus in Co-culture With P. aeruginosa While Growing as Biofilms. Front. Microbiol. 2018, 9, 3291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelsen, C.F.; Christensen, A.M.; Bojer, M.S.; Hoiby, N.; Ingmer, H.; Jelsbak, L. Staphylococcus aureus alters growth activity, autolysis, and antibiotic tolerance in a human host-adapted Pseudomonas aeruginosa lineage. J. Bacteriol. 2014, 196, 3903–3911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armbruster, C.R.; Wolter, D.J.; Mishra, M.; Hayden, H.S.; Radey, M.C.; Merrihew, G.; MacCoss, M.J.; Burns, J.; Wozniak, D.J.; Parsek, M.R. Staphylococcus aureus protein A mediates interspecies interactions at the cell surface of Pseudomonas aeruginosa. MBio 2016, 7, e00538-16. [Google Scholar] [CrossRef] [Green Version]
- Baldan, R.; Cigana, C.; Testa, F.; Bianconi, I.; De Simone, M.; Pellin, D.; Di Serio, C.; Bragonzi, A.; Cirillo, D.M. Adaptation of Pseudomonas aeruginosa in Cystic Fibrosis airways influences virulence of Staphylococcus aureus in vitro and murine models of co-infection. PLoS ONE 2014, 9, e89614. [Google Scholar] [CrossRef] [PubMed]
- Tavernier, S.; Crabbe, A.; Hacioglu, M.; Stuer, L.; Henry, S.; Rigole, P.; Dhondt, I.; Coenye, T. Community Composition Determines Activity of Antibiotics against Multispecies Biofilms. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radlinski, L.; Rowe, S.E.; Kartchner, L.B.; Maile, R.; Cairns, B.A.; Vitko, N.P.; Gode, C.J.; Lachiewicz, A.M.; Wolfgang, M.C.; Conlon, B.P. Pseudomonas aeruginosa exoproducts determine antibiotic efficacy against Staphylococcus aureus. PLoS Biol. 2017, 15, e2003981. [Google Scholar] [CrossRef]
- Webb, J.S.; Barratt, S.R.; Sabev, H.; Nixon, M.; Eastwood, I.M.; Greenhalgh, M.; Handley, P.S.; Robson, G.D. Green fluorescent protein as a novel indicator of antimicrobial susceptibility in Aureobasidium pullulans. Appl. Environ. Microbiol. 2001, 67, 5614–5620. [Google Scholar] [CrossRef] [Green Version]
- Steffy, K.; Shanthi, G.; Maroky, A.S.; Selvakumar, S. Synthesis and characterization of ZnO phytonanocomposite using Strychnos nux-vomica L. (Loganiaceae) and antimicrobial activity against multidrug-resistant bacterial strains from diabetic foot ulcer. J. Adv. Res. 2018, 9, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Burcham, Z.M.; Hood, J.A.; Pechal, J.L.; Krausz, K.L.; Bose, J.L.; Schmidt, C.J.; Benbow, M.E.; Jordan, H.R. Fluorescently labeled bacteria provide insight on post-mortem microbial transmigration. Forensic Sci. Int. 2016, 264, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Fong, S.A.; Drilling, A.; Morales, S.; Cornet, M.E.; Woodworth, B.A.; Fokkens, W.J.; Psaltis, A.J.; Vreugde, S.; Wormald, P.J. Activity of Bacteriophages in Removing Biofilms of Pseudomonas aeruginosa Isolates from Chronic Rhinosinusitis Patients. Front. Cell. Infect. Microbiol. 2017, 7, 418. [Google Scholar] [CrossRef] [Green Version]
- Lehman, S.M.; Mearns, G.; Rankin, D.; Cole, R.A.; Smrekar, F.; Branston, S.D.; Morales, S. Design and Preclinical Development of a Phage Product for the Treatment of Antibiotic-Resistant Staphylococcus aureus Infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Law, N.; Logan, C.; Yung, G.; Furr, C.L.; Lehman, S.M.; Morales, S.; Rosas, F.; Gaidamaka, A.; Bilinsky, I.; Grint, P.; et al. Successful adjunctive use of bacteriophage therapy for treatment of multidrug-resistant Pseudomonas aeruginosa infection in a cystic fibrosis patient. Infection 2019, 47, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ting, Y.P. Presence of Pseudomonas aeruginosa influences biofilm formation and surface protein expression of Staphylococcus aureus. Environ. Microbiol. 2015, 17, 4459–4468. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.R.; Perez-Esteban, P.; Kot, W.; Bean, J.E.; Arnot, T.; Hansen, L.H.; Enright, M.C.; Jenkins, A.T. A novel bacteriophage cocktail reduces and disperses Pseudomonas aeruginosa biofilms under static and flow conditions. Microb. Biotechnol. 2016, 9, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, S.; Fernandez, L.; Campelo, A.B.; Gutierrez, D.; Martinez, B.; Rodriguez, A.; Garcia, P. The Behavior of Staphylococcus aureus Dual-Species Biofilms Treated with Bacteriophage phiIPLA-RODI Depends on the Accompanying Microorganism. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Mendes, J.J.; Leandro, C.; Mottola, C.; Barbosa, R.; Silva, F.A.; Oliveira, M.; Vilela, C.L.; Melo-Cristino, J.; Gorski, A.; Pimentel, M.; et al. In vitro design of a novel lytic bacteriophage cocktail with therapeutic potential against organisms causing diabetic foot infections. J. Med Microbiol. 2014, 63, 1055–1065. [Google Scholar] [CrossRef]
- Ni, Y.; Deng, N.; Kokot, S. A simple kinetic spectrophotometric method for simultaneous determination of tetracyclines by use of chemometrics. Anal. Methods 2010, 2, 1302–1309. [Google Scholar] [CrossRef]
- Zheng, Z.; Tharmalingam, N.; Liu, Q.; Jayamani, E.; Kim, W.; Fuchs, B.B.; Zhang, R.; Vilcinskas, A.; Mylonakis, E. Synergistic efficacy of Aedes aegypti antimicrobial peptide cecropin A2 and tetracycline against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2017, 61, e00686-17. [Google Scholar] [CrossRef] [Green Version]
- Pai, H.; Kim, J.-W.; Kim, J.; Lee, J.H.; Choe, K.W.; Gotoh, N. Carbapenem resistance mechanisms in Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents Chemother. 2001, 45, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, D.; Vandenheuvel, D.; Martinez, B.; Rodriguez, A.; Lavigne, R.; Garcia, P. Two Phages, phiIPLA-RODI and phiIPLA-C1C, Lyse Mono- and Dual-Species Staphylococcal Biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [Green Version]
- Sillankorva, S.; Neubauer, P.; Azeredo, J. Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 2010, 26, 567–575. [Google Scholar] [CrossRef] [Green Version]
- McNerney, R.; Wilson, S.; Sidhu, A.; Harley, V.; Al Suwaidi, Z.; Nye, P.; Parish, T.; Stoker, N. Inactivation of mycobacteriophage D29 using ferrous ammonium sulphate as a tool for the detection of viable Mycobacterium smegmatis and M. tuberculosis. Res. Microbiol. 1998, 149, 487–495. [Google Scholar] [CrossRef]
- O’Toole, G.A. Classic Spotlight: Plate Counting You Can Count On. J. Bacteriol. 2016, 198, 3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, J.J.; Leandro, C.; Corte-Real, S.; Barbosa, R.; Cavaco-Silva, P.; Melo-Cristino, J.; Gorski, A.; Garcia, M. Wound healing potential of topical bacteriophage therapy on diabetic cutaneous wounds. Wound Repair Regen 2013, 21, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, R.; Costerton, J.W.; Raoult, D.; Cutler, S.J. The polymicrobial nature of biofilm infection. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013, 19, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Antonic, V.; Stojadinovic, A.; Zhang, B.; Izadjoo, M.J.; Alavi, M. Pseudomonas aeruginosa induces pigment production and enhances virulence in a white phenotypic variant of Staphylococcus aureus. Infect. Drug Resist. 2013, 6, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Lagendijk, E.L.; Validov, S.; Lamers, G.E.; de Weert, S.; Bloemberg, G.V. Genetic tools for tagging Gram-negative bacteria with mCherry for visualization in vitro and in natural habitats, biofilm and pathogenicity studies. FEMS Microbiol. Lett. 2010, 305, 81–90. [Google Scholar] [CrossRef]
- Chudakov, D.M.; Lukyanov, S.; Lukyanov, K.A. Fluorescent proteins as a toolkit for in vivo imaging. Trends Biotechnol. 2005, 23, 605–613. [Google Scholar] [CrossRef]
- Malone, C.L.; Boles, B.R.; Lauderdale, K.J.; Thoendel, M.; Kavanaugh, J.S.; Horswill, A.R. Fluorescent reporters for Staphylococcus aureus. J. Microbiol. Methods 2009, 77, 251–260. [Google Scholar] [CrossRef] [Green Version]
- Vickerman, M.M.; Mansfield, J.M.; Zhu, M.; Walters, K.S.; Banas, J.A. Codon-optimized fluorescent mTFP and mCherry for microscopic visualization and genetic counterselection of streptococci and enterococci. J. Microbiol. Methods 2015, 116, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Pereira, P.M.; Veiga, H.; Jorge, A.M.; Pinho, M.G. Fluorescent reporters for studies of cellular localization of proteins in Staphylococcus aureus. Appl. Environ. Microbiol. 2010, 76, 4346–4353. [Google Scholar] [CrossRef] [Green Version]
- Maksimow, M.; Hakkila, K.; Karp, M.; Virta, M. Simultaneous detection of bacteria expressing GFP and DsRed genes with a flow cytometer. Cytometry 2002, 47, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Steff, A.M.; Fortin, M.; Arguin, C.; Hugo, P. Detection of a decrease in green fluorescent protein fluorescence for the monitoring of cell death: An assay amenable to high-throughput screening technologies. Cytometry 2001, 45, 237–243. [Google Scholar] [CrossRef]
- Mashburn, L.M.; Jett, A.M.; Akins, D.R.; Whiteley, M. Staphylococcus aureus serves as an iron source for Pseudomonas aeruginosa during in vivo coculture. J. Bacteriol. 2005, 187, 554–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Yang, D.; Cirillo, S.L.; Li, S.; Akin, A.; Francis, K.P.; Maloney, T.; Cirillo, J.D. Application of Fluorescent Protein Expressing Strains to Evaluation of Anti-Tuberculosis Therapeutic Efficacy In Vitro and In Vivo. PLoS ONE 2016, 11, e0149972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, P.; Iturria, I.; Mohedano, M.L.; Caggianiello, G.; Rainieri, S.; Fiocco, D.; Angel Pardo, M.; Lopez, P.; Spano, G. Zebrafish gut colonization by mCherry-labelled lactic acid bacteria. Appl. Microbiol. Biotechnol. 2015, 99, 3479–3490. [Google Scholar] [CrossRef] [PubMed]
- Harcombe, W.; Bull, J. Impact of phages on two-species bacterial communities. Appl. Environ. Microbiol. 2005, 71, 5254–5259. [Google Scholar] [CrossRef] [Green Version]
- Kay, M.K.; Erwin, T.C.; McLean, R.J.; Aron, G.M. Bacteriophage ecology in Escherichia coli and Pseudomonas aeruginosa mixed-biofilm communities. Appl. Environ. Microbiol. 2011, 77, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Cerca, N.; Oliveira, R.; Azeredo, J. Susceptibility of Staphylococcus epidermidis planktonic cells and biofilms to the lytic action of staphylococcus bacteriophage K. Lett. Appl. Microbiol. 2007, 45, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Dvorackova, M.; Ruzicka, F.; Benesik, M.; Pantucek, R.; Dvorakova-Heroldova, M. Antimicrobial effect of commercial phage preparation Stafal(R) on biofilm and planktonic forms of methicillin-resistant Staphylococcus aureus. Folia Microbiol. 2019, 64, 121–126. [Google Scholar] [CrossRef]
- Harper, D.; Parracho, H.; Walker, J.; Sharp, R.; Hughes, G.; Werthén, M.; Lehman, S.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270–284. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.R.; Gaudion, A.; Bean, J.; Esteban, P.P.; Arnot, T.; Harper, D.; Kot, W.; Hansen, L.H.; Enright, M.; Jenkins, A.T.A. Combined use of bacteriophage K and a novel bacteriophage to reduce Staphylococcus aureus biofilm formation. Appl. Environ. Microbiol. 2014, 80, 6694–6703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, K.; Skillman, L.C.; Sutherland, I.W. The efficacy of bacteriophage as a method of biofilm eradication. Biofouling 2002, 18, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Ryu, J.H.; Beuchat, L.R. Inactivation of Escherichia coli O157:H7 in biofilm on stainless steel by treatment with an alkaline cleaner and a bacteriophage. J. Appl. Microbiol. 2005, 99, 449–459. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates Combination | Evaluated Isolate | Bacterial Cell Counts after Treatment | ||||
|---|---|---|---|---|---|---|
| PBS | AB-SA01 | AB-PA01 | AB-SA01+ AB-PA01 | Gentamicin | ||
| S. aureus KUB7 S and GFP PAO1 P | S. aureus KUB7 S | 5.5 | 3.6 | 8.7 | 3.0 | 0 |
| GFP PAO1 P | 7.9 | 8.9 | 0 | 3.7 | 0 | |
| 63-6538 S and 63-6598 P | 63-6538 S | 5.6 | 1.5 | 7.9 | 0 | 0 |
| 63-6598 P | 5.3 | 6.0 | 0 | 0 | 0 | |
| 63-2498 S and 63-5497 P | 63-2498 S | 6.2 | 0 | 6.6 | 0 | 0 |
| 63-5497 P | 7.5 | 8.0 | 3.5 | 4.0 | 0 | |
| 63-5656 S and 63-6036 P | 63-5656 S | 4.8 | 3.6 | 6.1 | 0 | 0 |
| 63-6036 P | 6.6 | 8.1 | 3.3 | 4.2 | 0 | |
| Summarized treatment effect | ||||||
| S. aureus mean | 5.5 | 2.2 | 7.3 | 0.8 | 0 | |
| S. aureus reduction | --- | 3.3 | +1.8 | 4.7 | 5.5 | |
| P. aeruginosa mean | 6.8 | 7.8 | 1.7 | 3.0 | 0 | |
| P. aeruginosa reduction | --- | +1.0 | 5.1 | 3.8 | 6.8 | |
| Isolates Combination | Evaluated Isolate | Bacterial Cell Counts after Treatment | ||||
|---|---|---|---|---|---|---|
| PBS | AB-SA01 | AB-PA01 | AB-SA01+ AB-PA01 | Tetracycline | ||
| S. aureus KUB7 S and PAO1 GFP P | S. aureus KUB7 S | 6.2 | 4.5 | 7.4 | 5.5 | 3.8 |
| PAO1 GFP P | 6.4 | 6.3 | 3.8 | 4.0 | 3.9 | |
| 63-6538 S and 63-6598 P | 63-6538 S | 5.2 | 4.4 | 4.7 | 3.6 | 3.0 |
| 63-6598 P | 5.5 | 5.6 | 3.6 | 3.7 | 0 | |
| 63-2498 S and 63-5497 P | 63-2498 S | 6.3 | 4.4 | 5.2 | 5.5 | 0 |
| 63-5497 P | 7.1 | 6.2 | 5.5 | 5.2 | 0 | |
| 63-5656S and 63-6036P | 63-5656 S | 7 | 4.9 | 5.9 | 5.3 | 0 |
| 63-6036 P | 8.5 | 8.9 | 4.7 | 6.3 | 0 | |
| Summarized treatment effect | ||||||
| S. aureus mean | 6.2 | 4.6 | 5.8 | 5.0 | 1.7 | |
| S. aureus reduction | ---- | 1.6 | 0.4 | 1.2 | 4.5 | |
| P. aeruginosa mean | 6.9 | 6.8 | 4.4 | 4.8 | 1.0 | |
| P. aeruginosa reduction | ---- | 0.1 | 2.5 | 2.1 | 5.9 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kifelew, L.G.; Warner, M.S.; Morales, S.; Thomas, N.; Gordon, D.L.; Mitchell, J.G.; Speck, P.G. Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses 2020, 12, 559. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050559
Kifelew LG, Warner MS, Morales S, Thomas N, Gordon DL, Mitchell JG, Speck PG. Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses. 2020; 12(5):559. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050559
Chicago/Turabian StyleKifelew, Legesse Garedew, Morgyn S. Warner, Sandra Morales, Nicky Thomas, David L. Gordon, James G. Mitchell, and Peter G. Speck. 2020. "Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms" Viruses 12, no. 5: 559. https://0-doi-org.brum.beds.ac.uk/10.3390/v12050559