Arboviruses and Muscle Disorders: From Disease to Cell Biology

 , ,
, ,
Abstract
:1. Introduction
2. Muscular Symptoms Associated with Arboviral Infection
3. Muscle Alterations and Arboviral Myotropism in Humans
4. Animal Models of Arboviral Myopathies
4.1. Ross River Virus
4.2. Chikungunya Virus

{kind=link}
{kind=link}
{kind=link}
| Virus (Family, Genus) | Virus (Species) | Animal Study (N) | Alteration, Histopathology/Viral Markers, In Vivo Tropism (Muscle) | References [n°] |
|---|---|---|---|---|
| Togaviridae Alphavirus | Ross River virus | mice C57BL/6N (3) | viral replication * | [72] |
| mice C57BL/6J mice B6-SJL mice Irf1−/− C57BL/6 (2–3) | viral antigens, infectious virus * | [76] | ||
| outbred mice mice macrophage(−/−) (3–4) | disruption, inflammatory infiltrate, pro-inflammatory cytokines/viral replication * | [66] | ||
| outbred mice CD-1 adult mice C57BL/6J mice RAG-1(−/−)(3–6) | inflammatory infiltrate/viral RNA, viral replication * | [67] | ||
| mice C57BL/6J mice C3−/− mice RAG-1(−/−) (3–6) | damage, inflammatory infiltrate/viral RNA, viral replication * | [69] | ||
| mice C57BL/6J mice MIF−/− (5) | disruption, inflammatory infiltrate/viral replication * | [71] | ||
| mice C57BL/6 mice MBL-DKO mice C1q−/− mice fB−/− (2–3) | disruption, inflammation/viral RNA, viral replication * | [77] | ||
| mice C57BL/6J mice CD74−/− mice MIF−/− (2–6) | disruption, inflammatory infiltrate/viral replication * | [78] | ||
| adult mice C57BL/6J mice TLR7−/− mice Myd88−/− (5–8) | disruption, inflammatory infiltrate/infectious virus * | [68] | ||
| mice C57BL/6J mice CD8α−/− mice Rag1−/− (2–6) | inflammatory infiltrate/viral replication * | [70] | ||
| mice C57BL/6J mice CCR2-DTR mice C57BL/6J Irf3−/−, Irf7−/− (3) | inflammation/viral replication * | [79] | ||
| Chikungunya virus | new-born mice (3–8) | structural degeneration, necrosis, inflammatory infiltrate, inflammatory cytokines/viral antigens, viral replication * | [74] | |
| young mice C57BL/6J (3–4) | necrosis, inflammatory infiltrate/viral replication * | [75] | ||
| new-born mice CD-1 young mice ICR (3–4) | degeneration, necrosis, fibrosis, dystrophic calcification, inflammation/viral antigens, viral replication * | [65] | ||
| mice C57BL/6J mice B6-SJL mice C57BL/6J Irf1−/− (2–3) | inflammation/viral antigens, viral RNA, infectious virus * | [76] | ||
| outbred mice OF1 new-born mice C57BL/6 mice 129s/v mice IFN-α/ßR−/− 129s/v (4) | myofiber necrosis, inflammatory infiltrate/viral antigens #, viral replication * | [64] | ||
| mice C57BL/6 mice IFNAR−/− (4–5) | disruption, necrosis, inflammatory infiltrate/viral burden ° * | [80] | ||
| mice C57BL/6J mice CD74−/− (2–6) | inflammation | [78] | ||
| Hamster (5–14) | necrosis, inflammation (myositis, tenosynovitis, myocarditis, fasciitis)/virus burden * | [81] | ||
| Zebrafish (3–5) | viral antigens | [82] | ||
| cynomolgus macaque (12) | focal necrosis, inflammatory infiltrate/viral antigen-RNA (persistent at 1.5 months post-inoculation §), viral replication * | [83] | ||
| rhesus macaque (2–6) | fiber necrosis, mononuclear infiltrate (perimysium)/viral RNA (persistent at 21 dpi) | [84] | ||
| Sindbis virus | neonatal mice CD-1 (3) | necrosis, inflammatory infiltrate/viral RNA | [85] | |
| Barmah Forest virus | Swiss outbred mice mice C57BL/6 (3–5) | inflammatory infiltrate/viral replication * | [86] | |
| Mayaro virus | mice Balb/c (3) | fiber degeneration, inflammation, MCP-1/viral replication * | [87] | |
| mice SV129 mice IFNAR−/− mice C57BL/6 mice RAG−/− (4–6) | damage, necrosis, inflammation/infectious virus * | [88] | ||
| FlaviviridaeFlavivirus | Zika virus | mice 129Sv mice AG129 (2–4) | multifocal myofiber degeneration, necrosis, nuclear rowing, attempted regeneration, infiltrate/viral replication * | [89] |
| rhesus macaque (7) | viral RNA (persistent at 35 dpi) | [90] |
4.3. Other Alphaviruses
4.4. Zika Virus
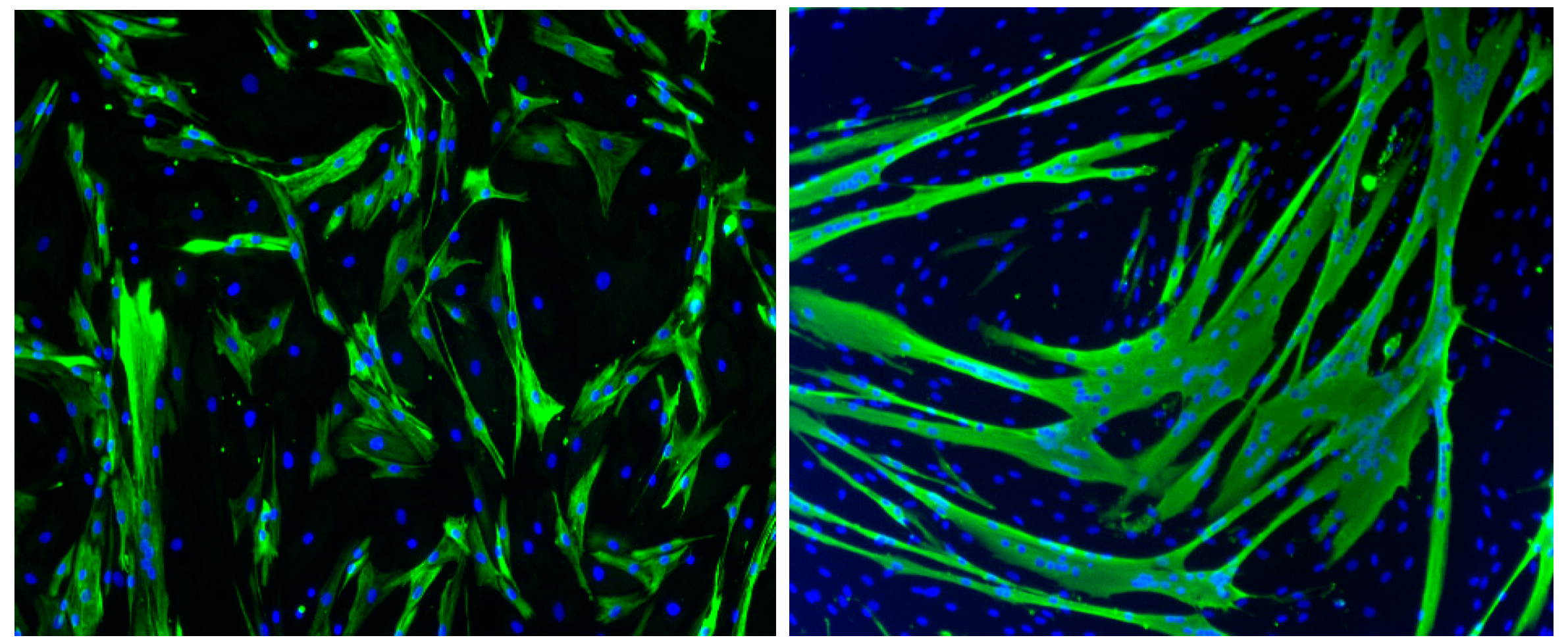
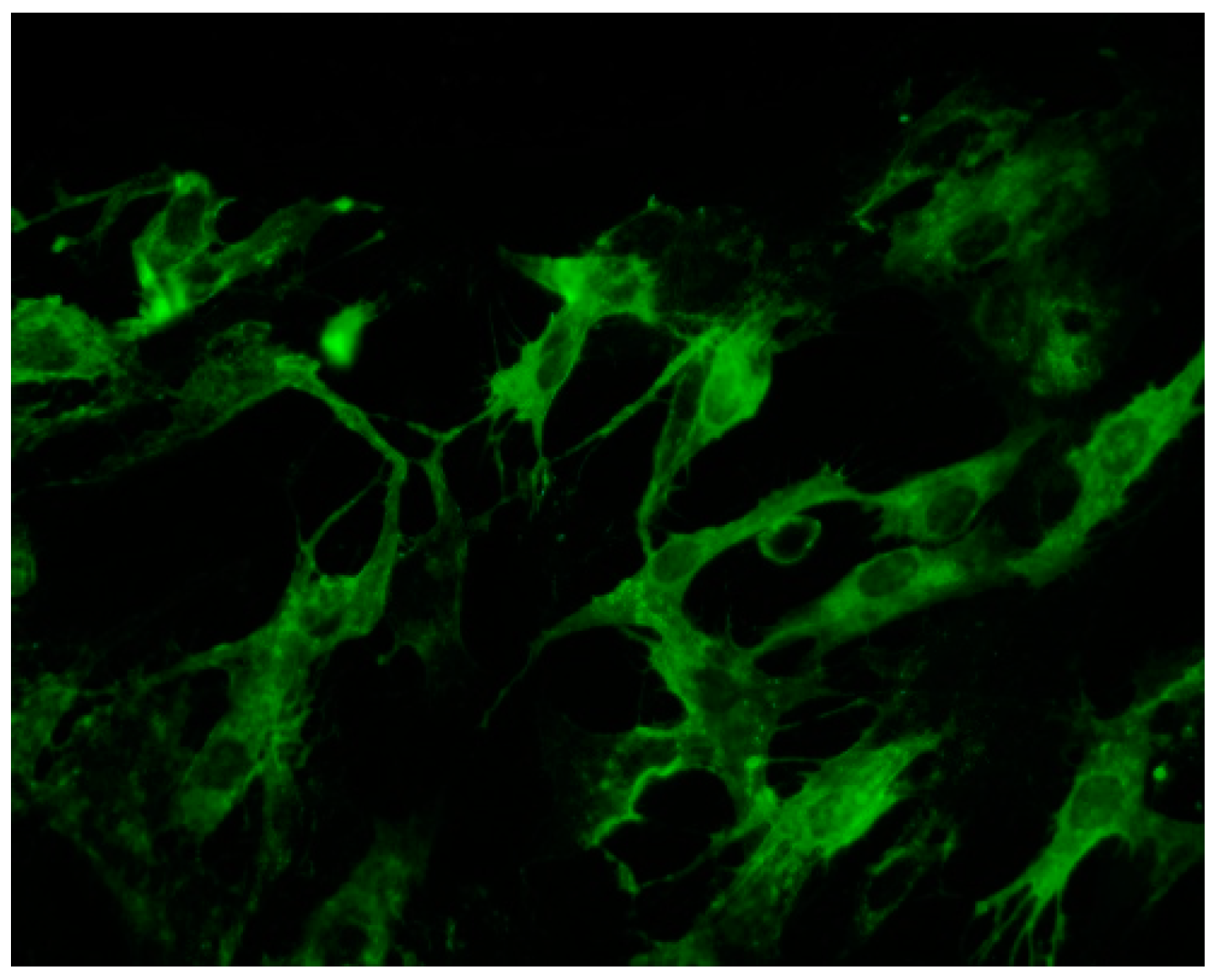
5. In Vitro Models of Muscle Cell Infection
5.1. Chikungunya Virus
5.2. Sindbis Virus
5.3. Zika Virus
5.4. Dengue Virus
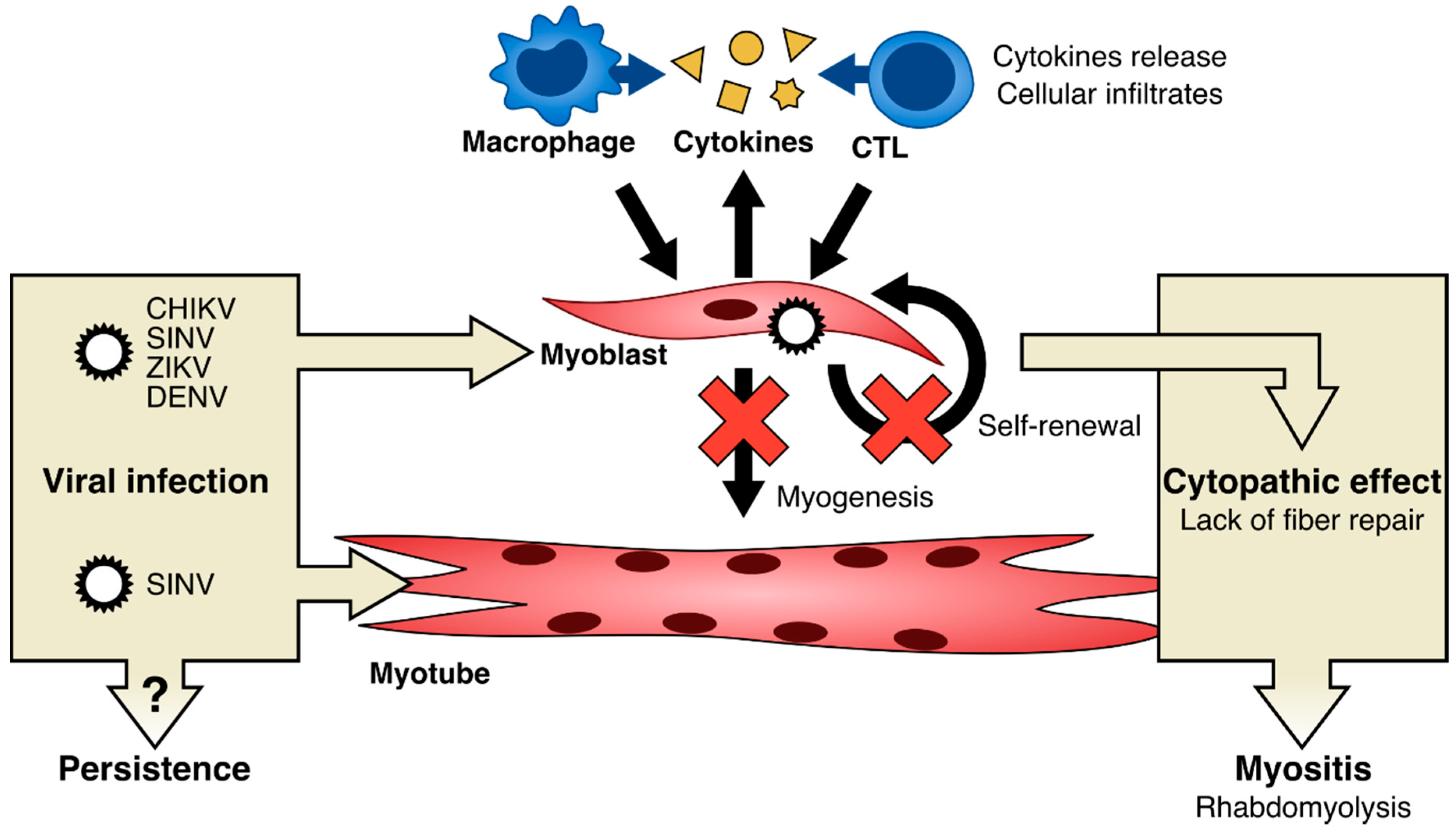
6. Possible Mechanisms of Pathophysiology of Arbovirus Associated Muscle Alterations
7. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.-C.; Lavenir, R.; Pardigon, N.; Reynes, J.-M.; Pettinelli, F.; et al. Genome Microevolution of Chikungunya Viruses Causing the Indian Ocean Outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef]
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Arboviruses: Molecular Biology, Evolution and Control; Vasilakis, N.; Gubler, D.J. (Eds.) Caister Academic Press: Norfolk, UK, 2016; ISBN 978-1-910190-22-7. [Google Scholar]
- Miller, F.W.; Lamb, J.A.; Schmidt, J.; Nagaraju, K. Risk factors and disease mechanisms in myositis. Nat. Rev. Rheumatol. 2018, 14, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Miller, F.W. State of the art: What we know about infectious agents and myositis. Curr. Opin. Rheumatol. 2011, 23, 585–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ytterberg, S.R. Infectious agents associated with myopathies. Curr. Opin. Rheumatol. 1996, 8, 507–513. [Google Scholar] [CrossRef]
- Singh, U.; Scheld, W.M. Infectious Etiologies of Rhabdomyolysis: Three Case Reports and Review. Clin. Infect. Dis. 1996, 22, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Crum-Cianflone, N.F. Bacterial, Fungal, Parasitic, and Viral Myositis. Clin. Microbiol. Rev. 2008, 21, 473–494. [Google Scholar] [CrossRef] [Green Version]
- Suhrbier, A.; Jaffar-Bandjee, M.-C.; Gasque, P. Arthritogenic alphaviruses—An overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.R.; Cunningham, A.L. Incubation time of epidemic polyarthritis. Med. J. Aust. 1980, 1, 550–551. [Google Scholar] [PubMed]
- Soden, M.; Vasudevan, H.; Roberts, B.; Coelen, R.; Hamlin, G.; Vasudevan, S.; La Brooy, J. Detection of viral ribonucleic acid and histologic analysis of inflamed synovium in Ross River virus infection. Arthritis Rheum. 2000, 43, 365–369. [Google Scholar] [CrossRef]
- Manimunda, S.P.; Vijayachari, P.; Uppoor, R.; Sugunan, A.P.; Singh, S.S.; Rai, S.K.; Sudeep, A.B.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R. Clinical progression of chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 392–399. [Google Scholar] [CrossRef]
- Hoarau, J.-J.; Jaffar Bandjee, M.-C.; Krejbich Trotot, P.; Das, T.; Li-Pat-Yuen, G.; Dassa, B.; Denizot, M.; Guichard, E.; Ribera, A.; Henni, T.; et al. Persistent Chronic Inflammation and Infection by Chikungunya Arthritogenic Alphavirus in Spite of a Robust Host Immune Response. J. Immunol. 2010, 184, 5914–5927. [Google Scholar] [CrossRef] [Green Version]
- Suhrbier, A.; La Linn, M. Clinical and pathologic aspects of arthritis due to Ross River virus and other alphaviruses. Curr. Opin. Rheumatol. 2004, 16, 374–379. [Google Scholar] [CrossRef]
- Sutaria, R.B.; Amaral, J.K.; Schoen, R.T. Emergence and treatment of chikungunya arthritis. Curr. Opin. Rheumatol. 2018, 30, 256–263. [Google Scholar] [CrossRef]
- Nakaya, H.I.; Gardner, J.; Poo, Y.-S.; Major, L.; Pulendran, B.; Suhrbier, A. Gene profiling of Chikungunya Virus arthritis in a mouse model reveals significant overlap with rheumatoid arthritis. Arthritis Rheum. 2012, 64, 3553–3563. [Google Scholar] [CrossRef]
- Chen, W.; Foo, S.-S.; Sims, N.A.; Herrero, L.J.; Walsh, N.C.; Mahalingam, S. Arthritogenic alphaviruses: New insights into arthritis and bone pathology. Trends Microbiol. 2015, 23, 35–43. [Google Scholar] [CrossRef]
- Krutikov, M.; Manson, J. Chikungunya Virus Infection: An Update on Joint Manifestations and Management. Rambam Maimonides Med. J. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Hazelton, R.A.; Hughes, C.; Aaskov, J.G. The inflammatory response in the synovium of a patient with Ross River arbovirus infection. Aust. N. Z. J. Med. 1985, 15, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Condon, R.J.; Rouse, I.L. Acute symptoms and sequelae of Ross River virus infection in South-Western Australia: A follow-up study. Clin. Diagn. Virol. 1995, 3, 273–284. [Google Scholar] [CrossRef]
- Dupuis-Maguiraga, L.; Noret, M.; Brun, S.; Le Grand, R.; Gras, G.; Roques, P. Chikungunya Disease: Infection-Associated Markers from the Acute to the Chronic Phase of Arbovirus-Induced Arthralgia. PLoS Negl. Trop. Dis. 2012, 6, e1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkoghe, D.; Kassa Kassa, R.F.; Bisvigou, U.; Caron, M.; Grard, G.; Leroy, E.M. No clinical or biological difference between Chikungunya and Dengue Fever during the 2010 Gabonese outbreak. Infect. Dis. Rep. 2012, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Bonifay, T.; Vesin, G.; Bidaud, B.; Bonnefoy, C.; Dueymes, M.; Nacher, M.; Djossou, F.; Epelboin, L. Clinical characteristics and predictive score of Dengue vs. Chikungunya Virus infections. Med. Mal. Infect. 2019, 49, 250–256. [Google Scholar] [CrossRef]
- Mallet, H.-P. Bilan de l’épidémie à virus Zika survenue en Polynésie française entre octobre 2013 et mars 2014. De la description de l’épidémie aux connaissances acquises après l’évènement. Bull. Epidémiologique Hebd. 2016, 367–373. [Google Scholar]
- Daudens-Vaysse, E.; Ledrans, M.; Gay, N.; Ardillon, V.; Cassadou, S.; Najioullah, F.; Leparc-Goffart, I.; Rousset, D.; Herrmann, C.; Cesaire, R.; et al. Zika emergence in the French Territories of America and description of first confirmed cases of Zika virus infection on Martinique, November 2015 to February 2016. Eurosurveillance 2016, 21, 30285. [Google Scholar] [CrossRef]
- Adouchief, S.; Smura, T.; Sane, J.; Vapalahti, O.; Kurkela, S. Sindbis virus as a human pathogen-epidemiology, clinical picture and pathogenesis. Rev. Med. Virol. 2016, 26, 221–241. [Google Scholar] [CrossRef]
- Seet, R.C.S.; Quek, A.M.L.; Lim, E.C.H. Post-infectious fatigue syndrome in dengue infection. J. Clin. Virol. 2007, 38, 1–6. [Google Scholar] [CrossRef]
- Sane, J.; Kurkela, S.; Desdouits, M.; Kalimo, H.; Mazalrey, S.; Lokki, M.-L.; Vaheri, A.; Helve, T.; Tornwall, J.; Huerre, M.; et al. Prolonged Myalgia in Sindbis Virus Infection: Case Description and In Vitro Infection of Myotubes and Myoblasts. J. Infect. Dis. 2012, 206, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Kurkela, S.; Helve, T.; Vaheri, A.; Vapalahti, O. Arthritis and arthralgia three years after Sindbis virus infection: Clinical follow-up of a cohort of 49 patients. Scand. J. Infect. Dis. 2008, 40, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Kiran, A.R.; Lau, R.A.; Wu, K.M.; Wong, A.L.; Clements, P.J.; Heinze, E.R. Fever, Myositis, and Paralysis: Is This Inflammatory Myopathy or Neuroinvasive Disease? Case Rep. Rheumatol. 2016, 2016, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, N. An Unusual Case of Myelitis with Myositis. J. Clin. Diagn. Res. 2016. [Google Scholar] [CrossRef]
- van den Berg, B.; van den Beukel, J.C.; Alsma, J.; van der Eijk, A.A.; Ruts, L.; van Doorn, P.A.; Jacobs, B.C.; Simon, C.C. Guillain-Barré-syndroom na zikavirus-infectie = Guillain-Barré syndrome following infection with the Zika virus. Ned. Tijdschr. Geneeskd. 2016, 160, D155. [Google Scholar]
- Martins, H.A. Chikungunya and Myositis: A Case Report in Brazil. J. Clin. Diagn. Res. 2016. [Google Scholar] [CrossRef]
- Ozden, S.; Huerre, M.; Riviere, J.-P.; Coffey, L.L.; Afonso, P.V.; Mouly, V.; de Monredon, J.; Roger, J.-C.; El Amrani, M.; Yvin, J.-L.; et al. Human Muscle Satellite Cells as Targets of Chikungunya Virus Infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef]
- Malheiros, S.M.; Oliveira, A.S.; Schmidt, B.; Lima, J.G.; Gabbai, A.A. Dengue. Muscle biopsy findings in 15 patients. Arq. Neuropsiquiatr. 1993, 51, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Paliwal, V.K.; Garg, R.K.; Juyal, R.; Husain, N.; Verma, R.; Sharma, P.K.; Verma, R.; Singh, M.K. Acute dengue virus myositis: A report of seven patients of varying clinical severity including two cases with severe fulminant myositis. J. Neurol. Sci. 2011, 300, 14–18. [Google Scholar] [CrossRef]
- Finsterer, J.; Kongchan, K. Severe, persisting, steroid-responsive Dengue myositis. J. Clin. Virol. 2006, 35, 426–428. [Google Scholar] [CrossRef]
- Arnež, M.; Avšič-Županc, T.; Ružić-Sabljić, E. Acute myositis associated with the initial phase of tick-borne encephalitis. J. Clin. Virol. 2011, 51, 276–278. [Google Scholar] [CrossRef]
- Acharya, S.; Shukla, S.; Mahajan, S.N.; Diwan, S.K. Acute dengue myositis with rhabdomyolysis and acute renal failure. Ann. Indian Acad. Neurol. 2010, 13, 221–222. [Google Scholar] [CrossRef]
- Davis, J.S.; Bourke, P. Rhabdomyolysis Associated with Dengue Virus Infection. Clin. Infect. Dis. 2004, 38, e109–e111. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.; Ghaffari, M.; Freire, A.X. Rhabdomyolysis in a patient with West Nile encephalitis and flaccid paralysis. Tenn. Med. J. Tenn. Med. Assoc. 2008, 101, 45–47. [Google Scholar]
- Sam, I.C.; Kamarulzaman, A.; Ong, G.S.Y.; Veriah, R.S.; Ponnampalavanar, S.; Chan, Y.F.; AbuBakar, S. Chikungunya Virus-associated death in Malaysia. Trop. Biomed. 2010, 27, 343–347. [Google Scholar]
- Ravanini, P.; Hasu, E.; Huhtamo, E.; Crobu, M.G.; Ilaria, V.; Brustia, D.; Salerno, A.M.; Vapalahti, O. Rhabdomyolysis and severe muscular weakness in a traveler diagnosed with Alkhurma hemorrhagic fever virus infection. J. Clin. Virol. 2011, 52, 254–256. [Google Scholar] [CrossRef]
- Bonifay, T.; Prince, C.; Neyra, C.; Demar, M.; Rousset, D.; Kallel, H.; Nacher, M.; Djossou, F.; Epelboin, L. Char Chik Working group Atypical and severe manifestations of Chikungunya Virus infection in French Guiana: A hospital-based study. PLoS ONE 2018, 13, e0207406. [Google Scholar] [CrossRef] [Green Version]
- Kulstad, E.B.; Wichter, M.D. West Nile encephalitis presenting as a stroke. Ann. Emerg. Med. 2003, 41, 283. [Google Scholar] [CrossRef]
- Samra, J.A.; Hagood, N.L.; Summer, A.; Medina, M.T.; Holden, K.R. Clinical Features and Neurologic Complications of Children Hospitalized With Chikungunya Virus in Honduras. J. Child Neurol. 2017, 32, 712–716. [Google Scholar] [CrossRef]
- Aletti, M.; Lecoules, S.; Kanczuga, V.; Soler, C.; Maquart, M.; Simon, F.; Leparc-Goffart, I. Transient myocarditis associated with acute Zika virus infection. Clin. Infect. Dis. 2016. [Google Scholar] [CrossRef]
- Obeyesekere, I.; Hermon, Y. Arbovirus heart disease: Myocarditis and cardiomyopathy following dengue and chikungunya fever--a follow-up study. Am. Heart J. 1973, 85, 186–194. [Google Scholar] [CrossRef]
- Lee, I.-K.; Lee, W.-H.; Liu, J.-W.; Yang, K.D. Acute myocarditis in dengue hemorrhagic fever: A case report and review of cardiac complications in dengue-affected patients. Int. J. Infect. Dis. 2010, 14, e919–e922. [Google Scholar] [CrossRef] [Green Version]
- Salgado, D.M.; Panqueba, C.A.; Castro, D.; R Vega, M.; Rodríguez, J.A. Miocarditis en niños con fiebre por dengue hemorrágico en un hospital universitario de Colombia = Myocarditis in children affected by dengue hemorrhagic fever in a teaching hospital in Colombia. Rev. Salud Publica Bogota Colomb. 2009, 11, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Hu, Z.; Huang, Y.; Li, J.; Hong, W.; Qin, Z.; Tong, Y.; Li, J.; Lv, M.; Li, M.; et al. Characterization of the Myocarditis during the worst outbreak of dengue infection in China. Medicine (Baltimore) 2016, 95, e4051. [Google Scholar] [CrossRef]
- Dev, N.; Kumar, R.; Gogna, A.; Sharma, S. Chikungunya-induced inflammatory myositis: A case report in India. Trop. Doct. 2019, 49, 241–243. [Google Scholar] [CrossRef]
- Misra, U.K.; Kalita, J.; Maurya, P.K.; Kumar, P.; Shankar, S.K.; Mahadevan, A. Dengue-associated transient muscle dysfunction: Clinical, electromyography and histopathological changes. Infection 2012, 40, 125–130. [Google Scholar] [CrossRef]
- Kalita, J.; Misra, U.K.; Maurya, P.K.; Shankar, S.K.; Mahadevan, A. Quantitative electromyography in dengue-associated muscle dysfunction. J. Clin. Neurophysiol. Off. Publ. Am. Electroencephalogr. Soc. 2012, 29, 468–471. [Google Scholar] [CrossRef]
- Verma, R.; Sharma, P.; Garg, R.K.; Atam, V.; Singh, M.K.; Malhotra, H.S. Neurological Complications of Dengue Fever: Experience From a Tertiary Center of North India. Ann. Indian Acad. Neurol. 2011, 14, 272–278. [Google Scholar] [CrossRef]
- Salgado, D.M.; Eltit, J.M.; Mansfield, K.; Panqueba, C.; Castro, D.; Vega, M.R.; Xhaja, K.; Schmidt, D.; Martin, K.J.; Allen, P.D.; et al. Heart and skeletal muscle are targets of dengue virus infection. Pediatr. Infect. Dis. J. 2010, 29, 238–242. [Google Scholar] [CrossRef] [Green Version]
- Póvoa, T.F.; Alves, A.M.B.; Oliveira, C.A.B.; Nuovo, G.J.; Chagas, V.L.A.; Paes, M.V. The pathology of severe dengue in multiple organs of human fatal cases: Histopathology, ultrastructure and virus replication. PLoS ONE 2014, 9, e83386. [Google Scholar] [CrossRef] [Green Version]
- Miranda, C.H.; Borges, M.D.C.; Matsuno, A.K.; Vilar, F.C.; Gali, L.G.; Volpe, G.J.; Schmidt, A.; Pazin-Filho, A.; da Silva, F.M.F.; de Castro-Jorge, L.A.; et al. Evaluation of cardiac involvement during dengue viral infection. Clin. Infect. Dis. Off. 2013, 57, 812–819. [Google Scholar] [CrossRef] [Green Version]
- Morrison, T.E.; Diamond, M.S. Animal Models of Zika Virus Infection, Pathogenesis, and Immunity. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Haese, N.N.; Broeckel, R.M.; Hawman, D.W.; Heise, M.T.; Morrison, T.E.; Streblow, D.N. Animal Models of Chikungunya Virus Infection and Disease. J. Infect. Dis. 2016, 214, S482–S487. [Google Scholar] [CrossRef] [Green Version]
- Couderc, T.; Chrétien, F.; Schilte, C.; Disson, O.; Brigitte, M.; Guivel-Benhassine, F.; Touret, Y.; Barau, G.; Cayet, N.; Schuffenecker, I.; et al. A Mouse Model for Chikungunya: Young Age and Inefficient Type-I Interferon Signaling Are Risk Factors for Severe Disease. PLoS Pathog. 2008, 4, e29. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.A.; Lu, L.; da Rosa, A.P.A.T.; Xiao, S.-Y.; Tesh, R.B. An animal model for studying the pathogenesis of Chikungunya Virus infection. Am. J. Trop. Med. Hyg. 2008, 79, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Lidbury, B.A.; Simeonovic, C.; Maxwell, G.E.; Marshall, I.D.; Hapel, A.J. Macrophage-Induced Muscle Pathology Results in Morbidity and Mortality for Ross River Virus–Infected Mice. J. Infect. Dis. 2000, 181, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Morrison, T.E.; Whitmore, A.C.; Shabman, R.S.; Lidbury, B.A.; Mahalingam, S.; Heise, M.T. Characterization of Ross River Virus Tropism and Virus-Induced Inflammation in a Mouse Model of Viral Arthritis and Myositis. J. Virol. 2006, 80, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Neighbours, L.M.; Long, K.; Whitmore, A.C.; Heise, M.T. Myd88-Dependent Toll-Like Receptor 7 Signaling Mediates Protection from Severe Ross River Virus-Induced Disease in Mice. J. Virol. 2012, 86, 10675–10685. [Google Scholar] [CrossRef] [Green Version]
- Morrison, T.E.; Fraser, R.J.; Smith, P.N.; Mahalingam, S.; Heise, M.T. Complement Contributes to Inflammatory Tissue Destruction in a Mouse Model of Ross River Virus-Induced Disease. J. Virol. 2007, 81, 5132–5143. [Google Scholar] [CrossRef] [Green Version]
- Burrack, K.S.; Montgomery, S.A.; Homann, D.; Morrison, T.E. CD8 + T Cells Control Ross River Virus Infection in Musculoskeletal Tissues of Infected Mice. J. Immunol. 2015, 194, 678–689. [Google Scholar] [CrossRef] [Green Version]
- Herrero, L.J.; Nelson, M.; Srikiatkhachorn, A.; Gu, R.; Anantapreecha, S.; Fingerle-Rowson, G.; Bucala, R.; Morand, E.; Santos, L.L.; Mahalingam, S. Critical role for macrophage migration inhibitory factor (MIF) in Ross River virus-induced arthritis and myositis. Proc. Natl. Acad. Sci. 2011, 108, 12048–12053. [Google Scholar] [CrossRef] [Green Version]
- Belarbi, E.; Legros, V.; Basset, J.; Desprès, P.; Roques, P.; Choumet, V. Bioluminescent Ross River Virus Allows Live Monitoring of Acute and Long-Term Alphaviral Infection by In Vivo Imaging. Viruses 2019, 11, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasque, P.; Couderc, T.; Lecuit, M.; Roques, P.; Ng, L.F.P. Chikungunya Virus pathogenesis and immunity. Vector Borne Zoonotic Dis. Larchmt. N 2015, 15, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Dhanwani, R.; Khan, M.; Lomash, V.; Rao, P.V.L.; Ly, H.; Parida, M. Characterization of Chikungunya Virus Induced Host Response in a Mouse Model of Viral Myositis. PLoS ONE 2014, 9, e92813. [Google Scholar] [CrossRef]
- Morrison, T.E.; Oko, L.; Montgomery, S.A.; Whitmore, A.C.; Lotstein, A.R.; Gunn, B.M.; Elmore, S.A.; Heise, M.T. A Mouse Model of Chikungunya Virus–Induced Musculoskeletal Inflammatory Disease. Am. J. Pathol. 2011, 178, 32–40. [Google Scholar] [CrossRef]
- Nair, S.; Poddar, S.; Shimak, R.M.; Diamond, M.S. Interferon Regulatory Factor 1 Protects against Chikungunya Virus-Induced Immunopathology by Restricting Infection in Muscle Cells. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Gunn, B.M.; Morrison, T.E.; Whitmore, A.C.; Blevins, L.K.; Hueston, L.; Fraser, R.J.; Herrero, L.J.; Ramirez, R.; Smith, P.N.; Mahalingam, S.; et al. Mannose Binding Lectin Is Required for Alphavirus-Induced Arthritis/Myositis. PLoS Pathog. 2012, 8, e1002586. [Google Scholar] [CrossRef] [Green Version]
- Herrero, L.J.; Sheng, K.-C.; Jian, P.; Taylor, A.; Her, Z.; Herring, B.L.; Chow, A.; Leo, Y.-S.; Hickey, M.J.; Morand, E.F.; et al. MIF receptor CD74 mediates alphavirus-induced arthritis and myositis: Requirement for CD74 in Alphavirus Arthritis and Myositis. Arthritis Rheum. 2013, 65, 2724–2736. [Google Scholar] [CrossRef] [Green Version]
- Haist, K.C.; Burrack, K.S.; Davenport, B.J.; Morrison, T.E. Inflammatory monocytes mediate control of acute alphavirus infection in mice. PLoS Pathog. 2017, 13, e1006748. [Google Scholar] [CrossRef]
- Rohatgi, A.; Corbo, J.C.; Monte, K.; Higgs, S.; Vanlandingham, D.L.; Kardon, G.; Lenschow, D.J.; Perlman, S. Infection of Myofibers Contributes to Increased Pathogenicity during Infection with an Epidemic Strain of Chikungunya Virus. J. Virol. 2014, 88, 2414–2425. [Google Scholar] [CrossRef] [Green Version]
- Bosco-Lauth, A.M.; Han, S.; Hartwig, A.; Bowen, R.A. Development of a Hamster Model for Chikungunya Virus Infection and Pathogenesis. PLoS ONE 2015, 10, e0130150. [Google Scholar] [CrossRef]
- Palha, N.; Guivel-Benhassine, F.; Briolat, V.; Lutfalla, G.; Sourisseau, M.; Ellett, F.; Wang, C.-H.; Lieschke, G.J.; Herbomel, P.; Schwartz, O.; et al. Real-Time Whole-Body Visualization of Chikungunya Virus Infection and Host Interferon Response in Zebrafish. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [Green Version]
- Labadie, K.; Larcher, T.; Joubert, C.; Mannioui, A.; Delache, B.; Brochard, P.; Guigand, L.; Dubreil, L.; Lebon, P.; Verrier, B.; et al. Chikungunya disease in nonhuman primates involves long-term viral persistence in macrophages. J. Clin. Investig. 2010, 120, 894–906. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-I.; Reisen, W.K.; Brault, A.C.; Pesavento, P.; Clark, D.C.; Luciw, P.A.; Lerche, N.W. Comparative Pathogenesis of Epidemic and Enzootic Chikungunya Viruses in a Pregnant Rhesus Macaque Model. Am. J. Trop. Med. Hyg. 2010, 83, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Trgovcich, J.; Aronson, J.F.; Johnston, R.E. Fatal Sindbis virus infection of neonatal mice in the absence of encephalitis. Virology 1996, 224, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Herrero, L.J.; Lidbury, B.A.; Bettadapura, J.; Jian, P.; Herring, B.L.; Hey-Cunningham, W.J.; Sheng, K.-C.; Zakhary, A.; Mahalingam, S. Characterization of Barmah Forest virus pathogenesis in a mouse model. J. Gen. Virol. 2014, 95, 2146–2154. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.M.; Dias, R.S.; de Oliveira, M.D.; Costa, I.C.T.A.; Fernandes, L.d.S.; Pessoa, C.R.; da Matta, S.L.P.; Costa, V.V.; Souza, D.G.; da Silva, C.C.; et al. Animal model of arthritis and myositis induced by the Mayaro virus. PLoS Negl. Trop. Dis. 2019, 13, e0007375. [Google Scholar] [CrossRef]
- Figueiredo, C.M.; Neris, R.L.d.S.; Gavino-Leopoldino, D.; da Silva, M.O.L.; Almeida, J.S.; Dos-Santos, J.S.; Figueiredo, C.P.; Bellio, M.; Bozza, M.T.; Assunção-Miranda, I. Mayaro Virus Replication Restriction and Induction of Muscular Inflammation in Mice Are Dependent on Age, Type-I Interferon Response, and Adaptive Immunity. Front. Microbiol. 2019, 10, 2246. [Google Scholar] [CrossRef]
- Aliota, M.T.; Caine, E.A.; Walker, E.C.; Larkin, K.E.; Camacho, E.; Osorio, J.E. Characterization of Lethal Zika Virus Infection in AG129 Mice. PLoS Negl. Trop. Dis. 2016, 10, e0004682. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, A.J.; Smith, J.L.; Haese, N.N.; Broeckel, R.M.; Parkins, C.J.; Kreklywich, C.; DeFilippis, V.R.; Denton, M.; Smith, P.P.; Messer, W.B.; et al. Zika Virus infection of rhesus macaques leads to viral persistence in multiple tissues. PLOS Pathog. 2017, 13, e1006219. [Google Scholar] [CrossRef]
- Lentscher, A.J.; McCarthy, M.K.; May, N.A.; Davenport, B.J.; Montgomery, S.A.; Raghunathan, K.; McAllister, N.; Silva, L.A.; Morrison, T.E.; Dermody, T.S. Chikungunya Virus replication in skeletal muscle cells is required for disease development. J. Clin. Investig. 2019. [Google Scholar] [CrossRef]
- Biacchesi, S.; Jouvion, G.; Mérour, E.; Boukadiri, A.; Desdouits, M.; Ozden, S.; Huerre, M.; Ceccaldi, P.-E.; Brémont, M. Rainbow trout (Oncorhynchus mykiss) muscle satellite cells are targets of salmonid alphavirus infection. Vet. Res. 2016, 47, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, J.J.; Diamond, M.S. Zika Virus Pathogenesis and Tissue Tropism. Cell Host Microbe 2017, 21, 134–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, K.M.; Lee, R.C.H.; Ng, M.M.-L.; Chu, J.J.H. Establishment of a Novel Primary Human Skeletal Myoblast Cellular Model for Chikungunya Virus Infection and Pathogenesis. Sci. Rep. 2016, 6, 21406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohachanakul, J.; Phuklia, W.; Thannagith, M.; Thongsakulprasert, T.; Smith, D.R.; Ubol, S. Differences in response of primary human myoblasts to infection with recent epidemic strains of Chikungunya Virus isolated from patients with and without myalgia. J. Med. Virol. 2015, 87, 733–739. [Google Scholar] [CrossRef]
- Issac, T.H.K.; Tan, E.L.; Chu, J.J.H. Proteomic profiling of Chikungunya Virus-infected human muscle cells: Reveal the role of cytoskeleton network in CHIKV replication. J. Proteomics 2014, 108, 445–464. [Google Scholar] [CrossRef]
- Thanapati, S.; Sudeep, A.B.; Kulkarni, S.P.; Tripathy, A.S. Regulation of the chikungunya-virus-induced innate inflammatory response by protein tyrosine phosphatase non-receptor 6 in muscle cells. Arch. Virol. 2018, 163, 243–248. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Yip, C.C.-Y.; Tsang, J.O.-L.; Tee, K.-M.; Cai, J.-P.; Chik, K.K.-H.; Zhu, Z.; Chan, C.C.-S.; Choi, G.K.-Y.; Sridhar, S.; et al. Differential cell line susceptibility to the emerging Zika virus: Implications for disease pathogenesis, non-vector-borne human transmission and animal reservoirs. Emerg. Microbes Infect. 2016, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Warke, R.V.; Becerra, A.; Zawadzka, A.; Schmidt, D.J.; Martin, K.J.; Giaya, K.; Dinsmore, J.H.; Woda, M.; Hendricks, G.; Levine, T.; et al. Efficient dengue virus (DENV) infection of human muscle satellite cells upregulates type I interferon response genes and differentially modulates MHC I expression on bystander and DENV-infected cells. J. Gen. Virol. 2008, 89, 1605–1615. [Google Scholar] [CrossRef]
- Bao, S.; King, N.J.; Dos Remedios, C.G. Flavivirus induces MHC antigen on human myoblasts: A model of autoimmune myositis? Muscle Nerve 1992, 15, 1271–1277. [Google Scholar] [CrossRef]
- Sourisseau, M.; Schilte, C.; Casartelli, N.; Trouillet, C.; Guivel-Benhassine, F.; Rudnicka, D.; Sol-Foulon, N.; Roux, K.L.; Prevost, M.-C.; Fsihi, H.; et al. Characterization of Reemerging Chikungunya Virus. PLoS Pathog. 2007, 3, e89. [Google Scholar] [CrossRef]
- Schwartz, O.; Albert, M.L. Biology and Pathogenesis of Chikungunya Virus. Nat. Rev. Microbiol. 2010, 8, 491–500. [Google Scholar] [CrossRef]
- Matusali, G.; Colavita, F.; Bordi, L.; Lalle, E.; Ippolito, G.; Capobianchi, M.; Castilletti, C. Tropism of the Chikungunya Virus. Viruses 2019, 11, 175. [Google Scholar] [CrossRef] [Green Version]
- Ozden, S.; Lucas-Hourani, M.; Ceccaldi, P.-E.; Basak, A.; Valentine, M.; Benjannet, S.; Hamelin, J.; Jacob, Y.; Mamchaoui, K.; Mouly, V.; et al. Inhibition of Chikungunya Virus infection in cultured human muscle cells by furin inhibitors: Impairment of the maturation of the E2 surface glycoprotein. J. Biol. Chem. 2008, 283, 21899–21908. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.R.; Mohamed Hussain, K.; Chu, J.J.H. Macropinocytosis dependent entry of Chikungunya Virus into human muscle cells. PLoS Negl. Trop. Dis. 2019, 13, e0007610. [Google Scholar] [CrossRef]
- Monel, B.; Compton, A.A.; Bruel, T.; Amraoui, S.; Burlaud-Gaillard, J.; Roy, N.; Guivel-Benhassine, F.; Porrot, F.; Génin, P.; Meertens, L.; et al. Zika virus induces massive cytoplasmic vacuolization and paraptosis-like death in infected cells. EMBO J. 2017, 36, 1653–1668. [Google Scholar] [CrossRef]
- McCarthy, M.K.; Morrison, T.E. Chronic Chikungunya Virus musculoskeletal disease: What are the underlying mechanisms? Future Microbiol. 2016, 11, 331–334. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.K.; Morrison, T.E. Persistent RNA virus infections: Do PAMPS drive chronic disease? Curr. Opin. Virol. 2017, 23, 8–15. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.K.; Davenport, B.J.J.; Morrison, T.E. Chronic Chikungunya Virus Disease. Curr. Top. Microbiol. Immunol. 2019. [Google Scholar] [CrossRef]
- Wintachai, P.; Wikan, N.; Kuadkitkan, A.; Jaimipuk, T.; Ubol, S.; Pulmanausahakul, R.; Auewarakul, P.; Kasinrerk, W.; Weng, W.-Y.; Panyasrivanit, M.; et al. Identification of prohibitin as a Chikungunya Virus receptor protein. J. Med. Virol. 2012, 84, 1757–1770. [Google Scholar] [CrossRef]
- Zhang, R.; Kim, A.S.; Fox, J.M.; Nair, S.; Basore, K.; Klimstra, W.B.; Rimkunas, R.; Fong, R.H.; Lin, H.; Poddar, S.; et al. Mxra8 is a receptor for multiple arthritogenic alphaviruses. Nature 2018, 557, 570–574. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef] [Green Version]
- Joubert, P.-E.; Werneke, S.W.; de la Calle, C.; Guivel-Benhassine, F.; Giodini, A.; Peduto, L.; Levine, B.; Schwartz, O.; Lenschow, D.J.; Albert, M.L. Chikungunya Virus–induced autophagy delays caspase-dependent cell death. J. Exp. Med. 2012, 209, 1029–1047. [Google Scholar] [CrossRef]



| Virus (Family, Genus) | Virus (Species) | Clinical Feature | Alteration, Histopathology/Viral Markers (Muscle) | Study | References [n°] |
|---|---|---|---|---|---|
| Togaviridae Alphavirus | Chikungunya virus | myositis | vacuolization, necrosis, infiltrating cells/viral antigens * | La Rèunion case reports (N = 2) | [37] |
| myositis | sarcoplasmic vacuolization, lymphocytic infiltrate | India case report (N = 1) | [55] | ||
| Sindbis virus | myalgia (chronic) | internal nuclei, MyHCn+ fibers | Finland case report (N = 1) | [31] | |
| Flaviviridae Flavivirus | Dengue virus | myalgia | perivascular mononuclear infiltrate, lipid accumulation | Brazil case reports (N = 12) | [38] |
| myositis | inflammatory cells, fibrotic areas | endemic areas case report (N = 1) | [39] | ||
| myositis | perifascicular inflammatory infiltrates, myonecrosis | India case report (N = 1) | [42] | ||
| myalgia | myonecrosis, myophagocitosis, inflammation | India case reports (N = 2) | [56] | ||
| myopathy | myonecrosis, myophagocytosis | India case reports (N = 2) | [57] | ||
| myositis | inflammatory infiltrate | India case reports (N = 2) | [58] | ||
| myocarditis † (hemorrhagic fever) | inflammatory markers (MCP-1)/viral antigens # | Colombia case report (N = 1) | [59] | ||
| myalgia (hemorrhagic fever) myocarditis † | degeneration of muscle fibers, mononuclear infiltrates/viral antigens-RNA ° | Brazil case reports (N = 4) | [60] | ||
| myocarditis † | inflammatory infiltrates/viral antigens § | Brazil case reports (N = 2) | [61] |
| Virus (Family, Genus) | Virus (Species) | Cell Study (N) | Alteration/Viral Markers, In Vitro Tropism (Muscle Cells) | References [n°] |
|---|---|---|---|---|
| TogaviridaeAlphavirus | Chikungunya virus | primary myoblasts/satellite cells (human) (2) | cytopathic effect/viral antigens | [37] |
| primary myoblasts (human) (3) | cytopathic effect/viral antigens, infectious virions | [94] | ||
| primary myoblasts (human) (4) | cytopathic effect, cytokines production/viral replication | [95] | ||
| myoblasts (murine) myotubes (murine) (2) | viral replication | [80] | ||
| rhabdomyosarcoma (human) (3) | cytopathic effect, cell death/viral replication | [96] | ||
| rhabdomyosarcoma (human) (3) | viral replication | [97] | ||
| Sindbis virus | primary myoblasts (human) primary myotubes (human) (2) | cytopathic effect/viral antigens, infectious virions | [31] | |
| FlaviviridaeFlavivirus | Zika virus | rhabdomyosarcoma (human) (3) | cytopathic effect/viral antigens, viral replication | [98] |
| primary myoblasts (human) (3) | cytopathic effect/viral antigens, viral replication | Legros V et al. (accepted) | ||
| Dengue virus | primary satellite cells (human) (2) | virus-like particles in cytoplasmic vacuoles/ viral antigens-RNA, viral replication | [99] | |
| primary myotubes (human) | disregulation of Ca2+/homeostasis/viral antigens | [59] | ||
| rhabdomyosarcoma (human) (3) | cytopathic effect/viral antigens, viral replication | [98] | ||
| West Nile virus | myoblasts (human embryonic) | viral antigens | [100] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippone, C.; Legros, V.; Jeannin, P.; Choumet, V.; Butler-Browne, G.; Zoladek, J.; Mouly, V.; Gessain, A.; Ceccaldi, P.-E. Arboviruses and Muscle Disorders: From Disease to Cell Biology. Viruses 2020, 12, 616. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060616
Filippone C, Legros V, Jeannin P, Choumet V, Butler-Browne G, Zoladek J, Mouly V, Gessain A, Ceccaldi P-E. Arboviruses and Muscle Disorders: From Disease to Cell Biology. Viruses. 2020; 12(6):616. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060616
Chicago/Turabian StyleFilippone, Claudia, Vincent Legros, Patricia Jeannin, Valérie Choumet, Gillian Butler-Browne, Jim Zoladek, Vincent Mouly, Antoine Gessain, and Pierre-Emmanuel Ceccaldi. 2020. "Arboviruses and Muscle Disorders: From Disease to Cell Biology" Viruses 12, no. 6: 616. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060616