mRNA Profile in Milk Extracellular Vesicles from Bovine Leukemia Virus-Infected Cattle

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Hematology
2.1.1. Detection of Serum Antibodies against BLV
2.1.2. Detection of BLV Provirus
2.1.3. Measurement of BLV Proviral Load
2.1.4. Measurement of LDH Isozymes
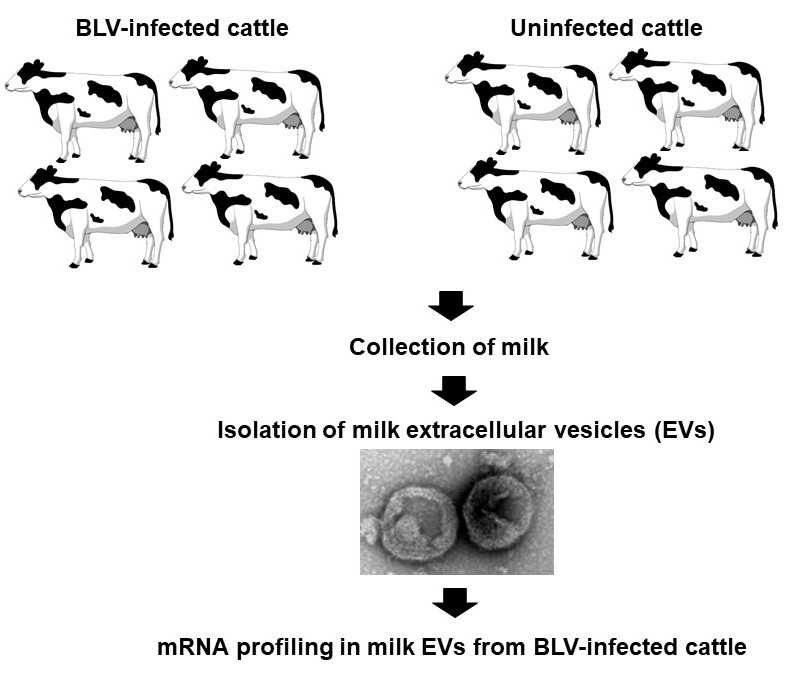
2.2. Classification of Milk Samples
2.2.1. EV Isolation
2.2.2. Transmission Electron Microscopy (TEM) Analysis of Milk EVs
2.2.3. RNA Extraction and cDNA Synthesis
2.2.4. Microarray Analysis
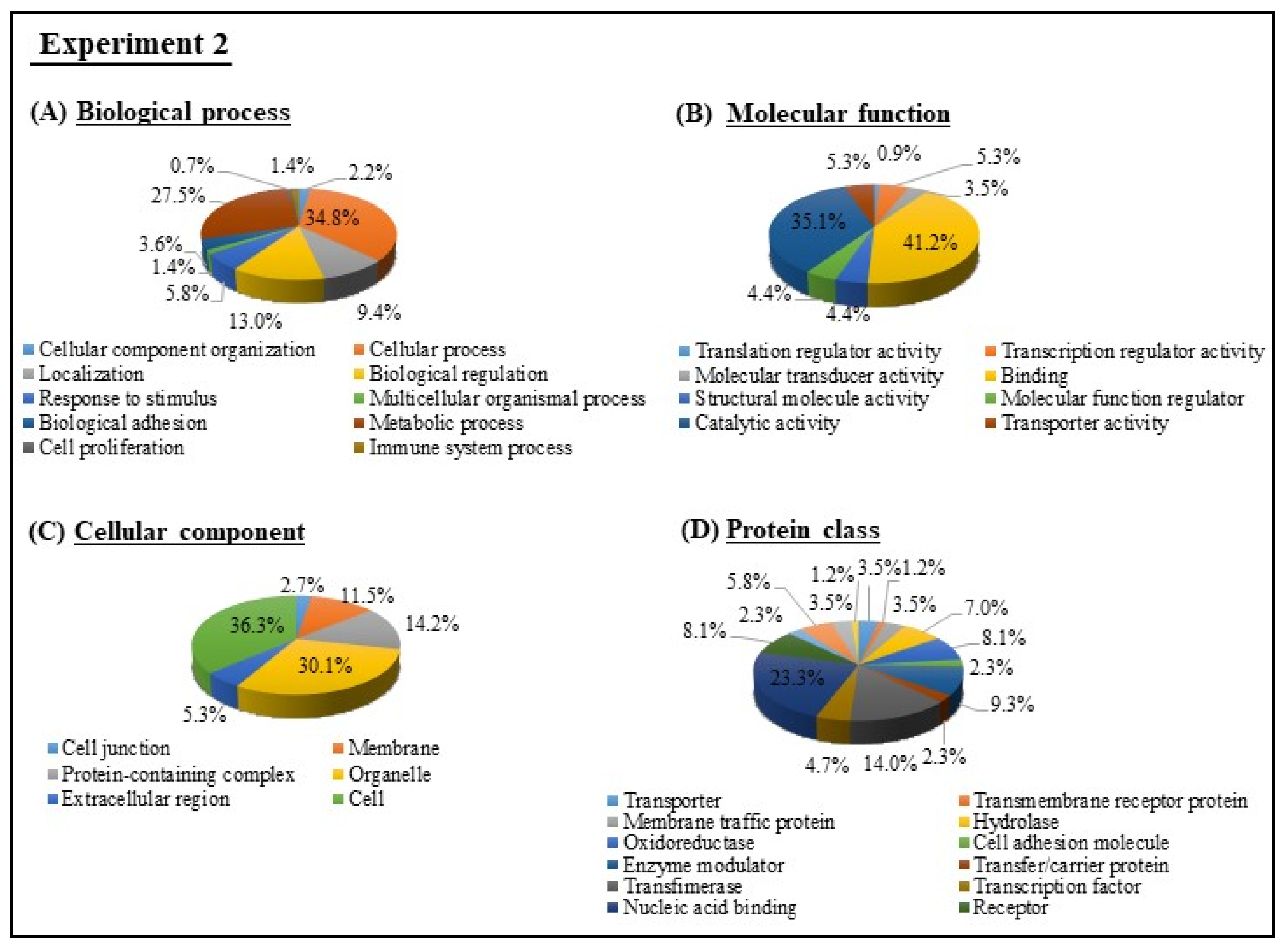
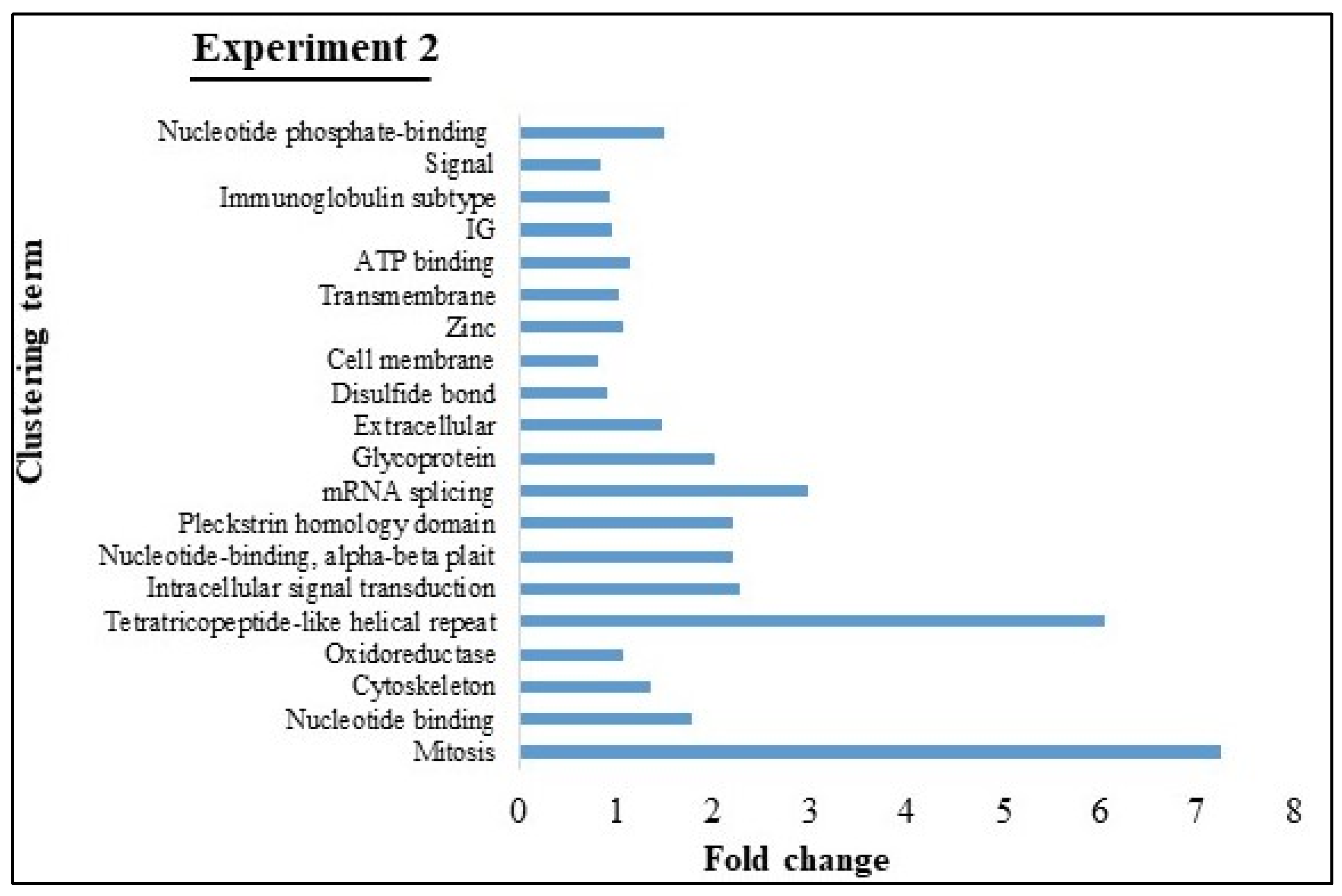
2.2.5. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analyses
2.3. Ethical Approval
3. Results
3.1. BLV Infection and Clinical Status
3.2. TEM Analysis of Milk EVs
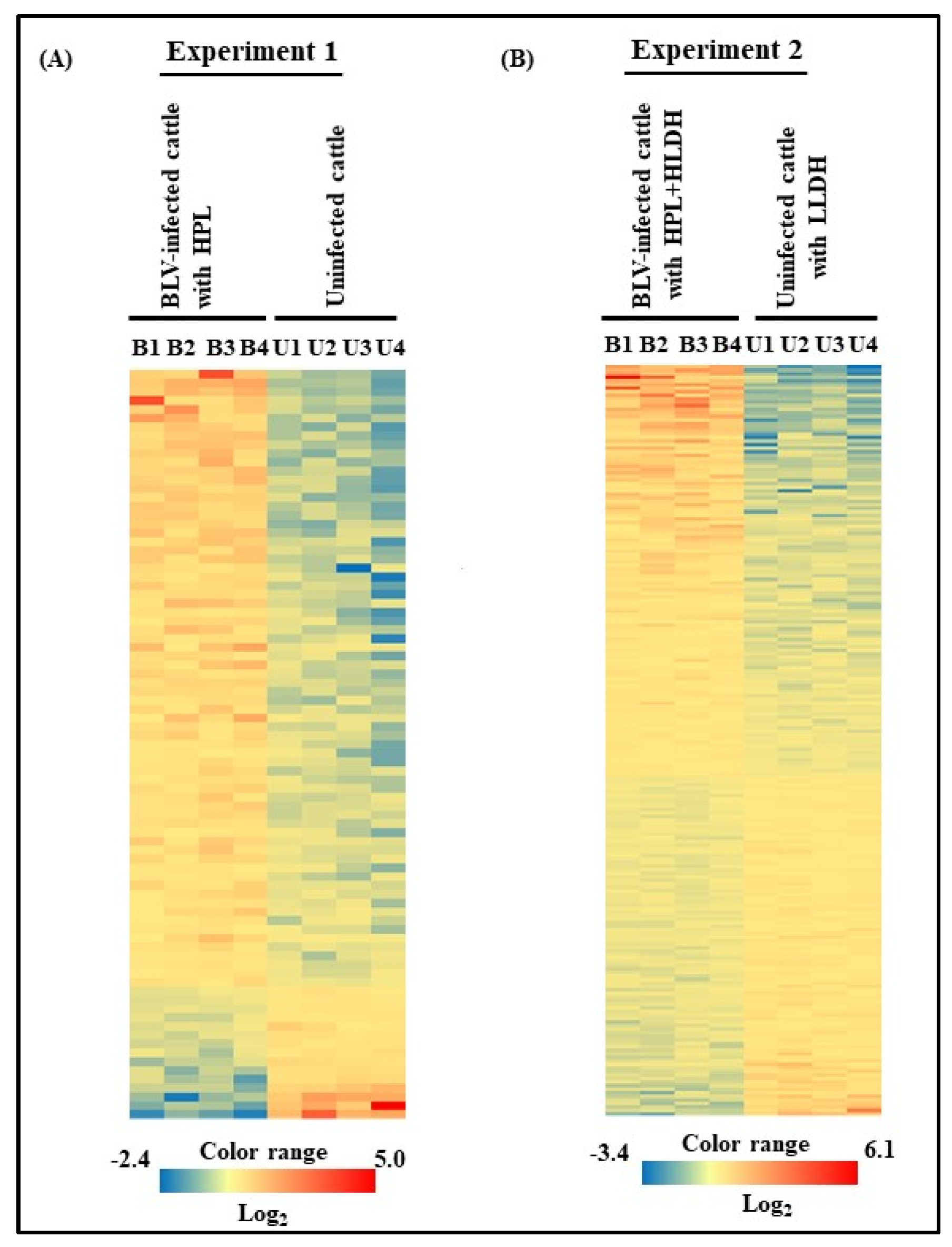
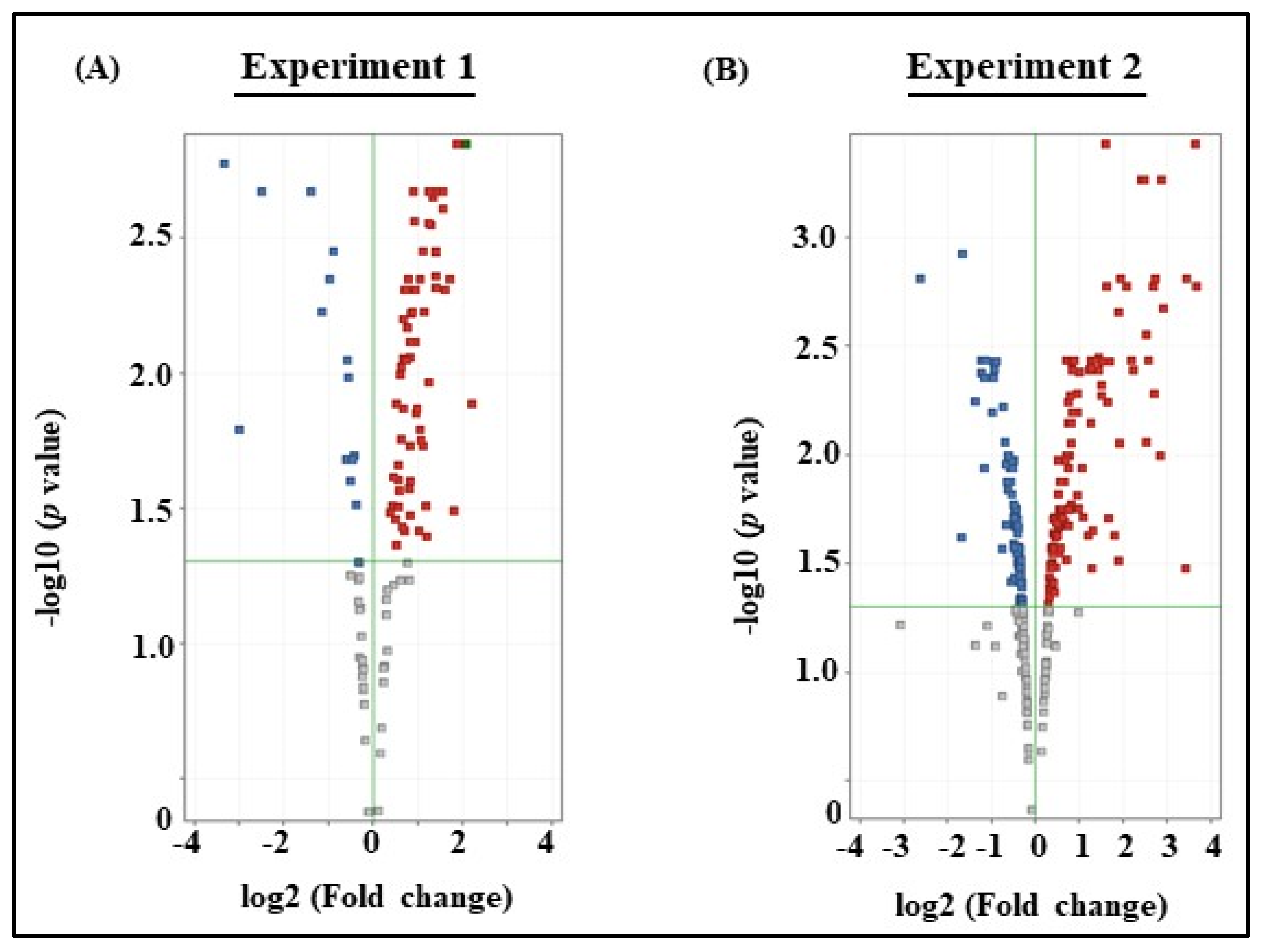
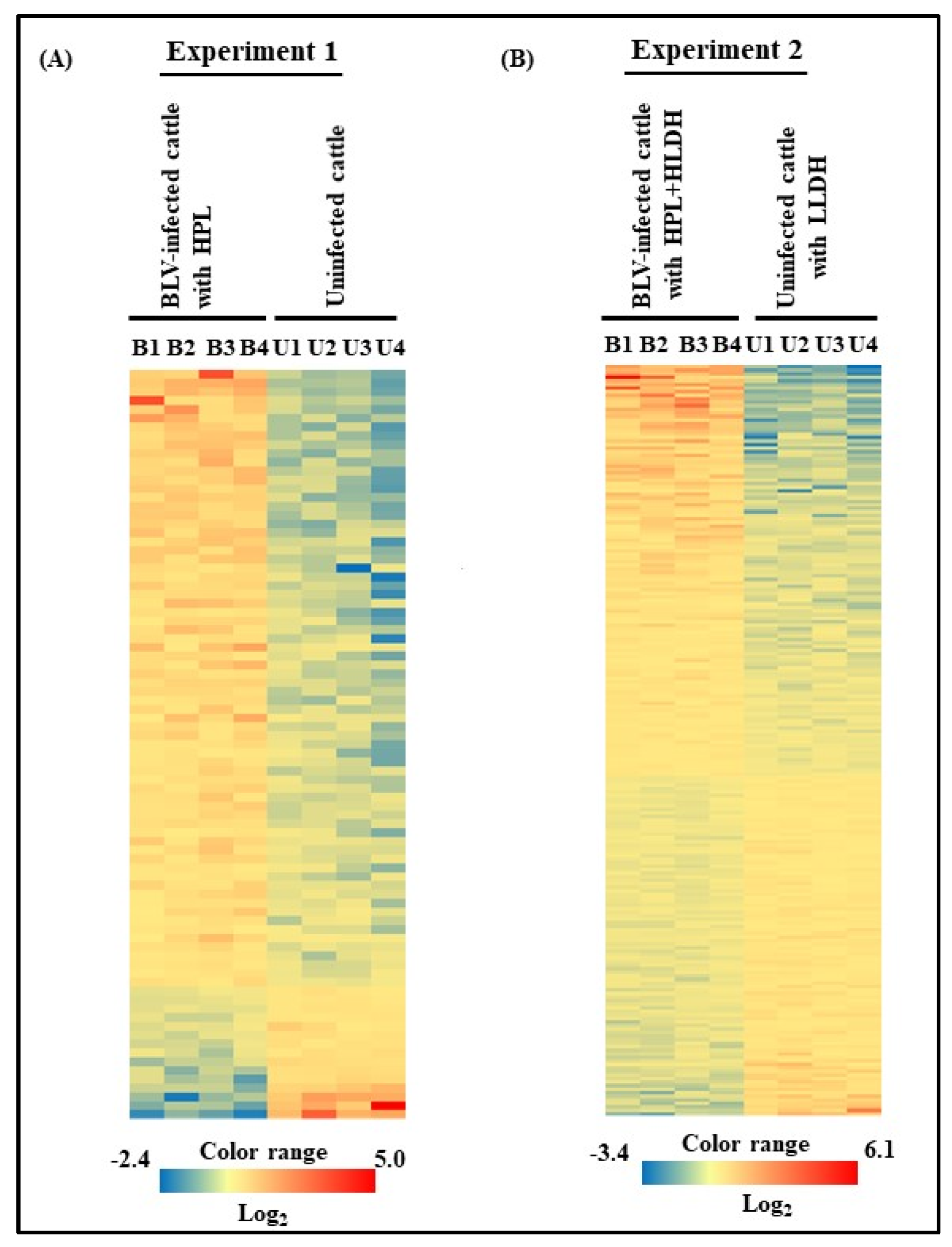
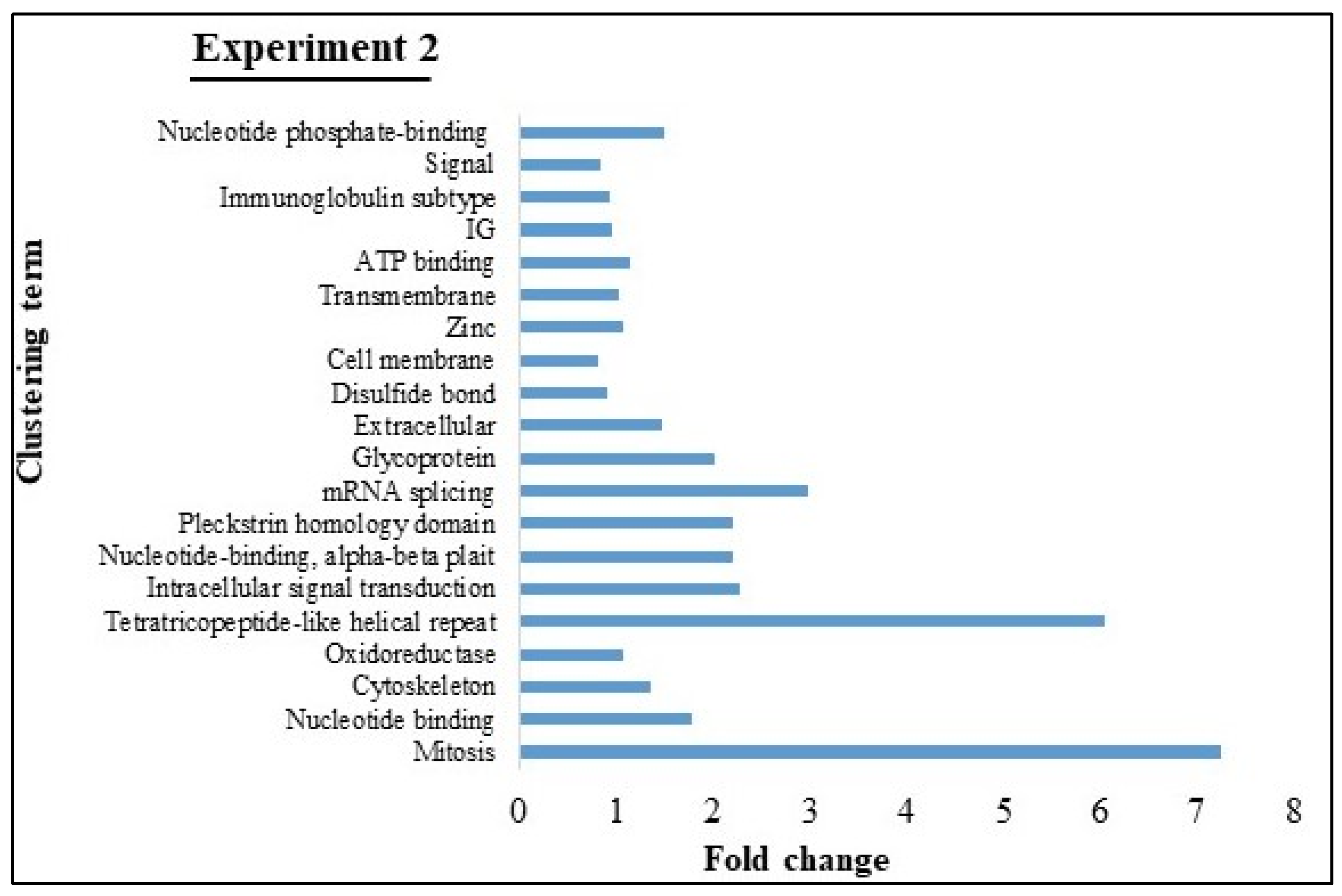
3.3. Microarray Analysis of Experiments 1 and 2
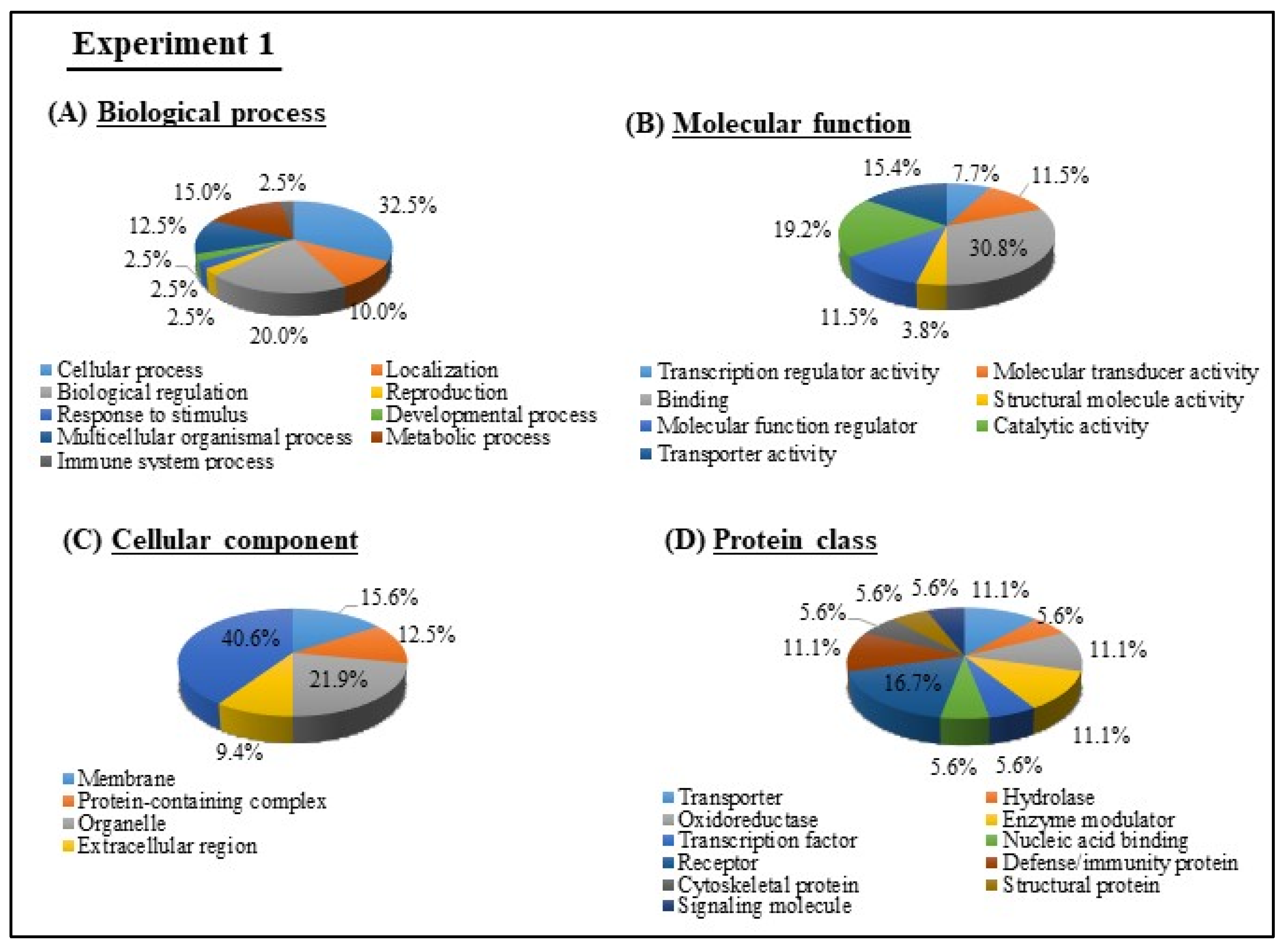
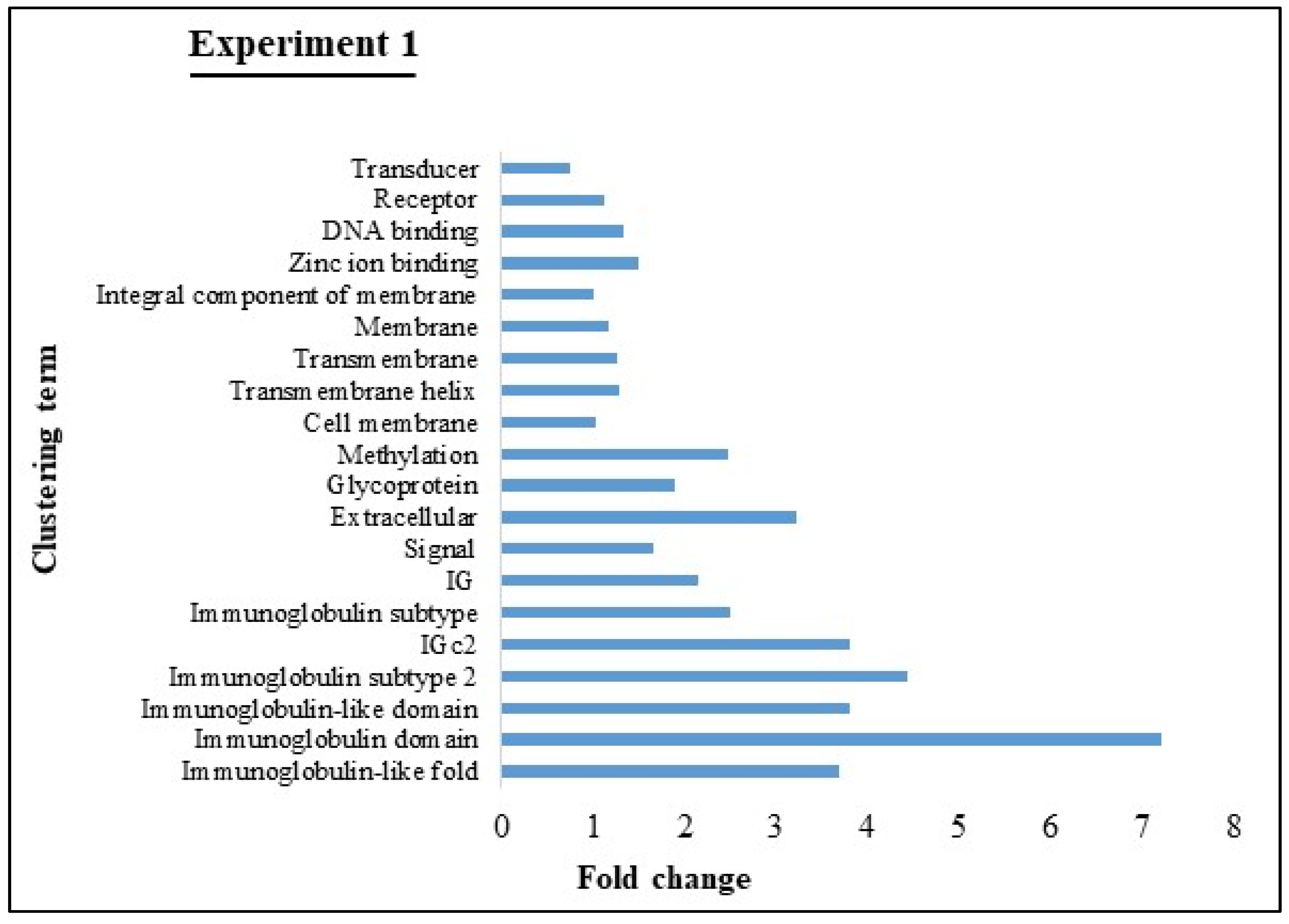
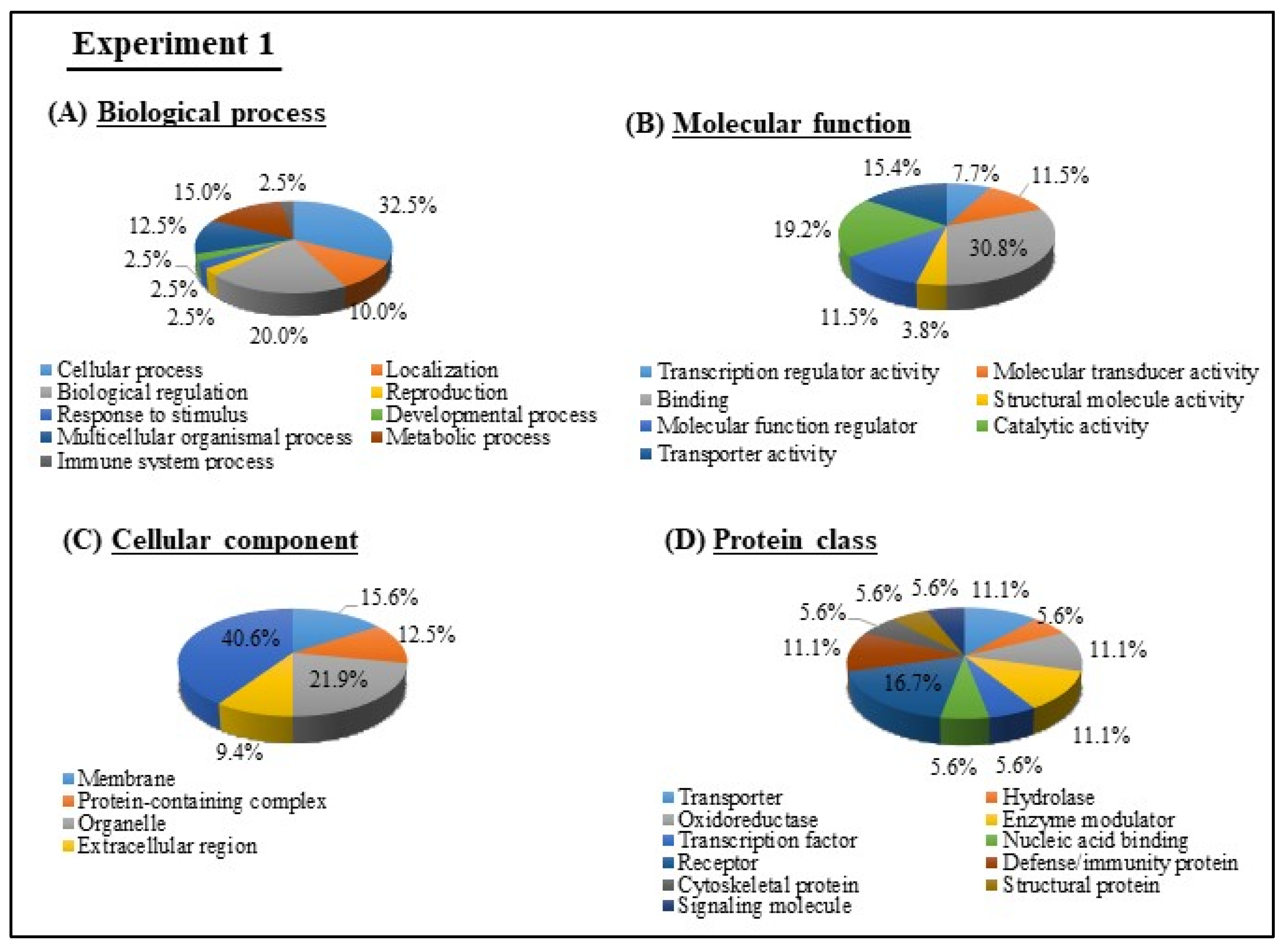
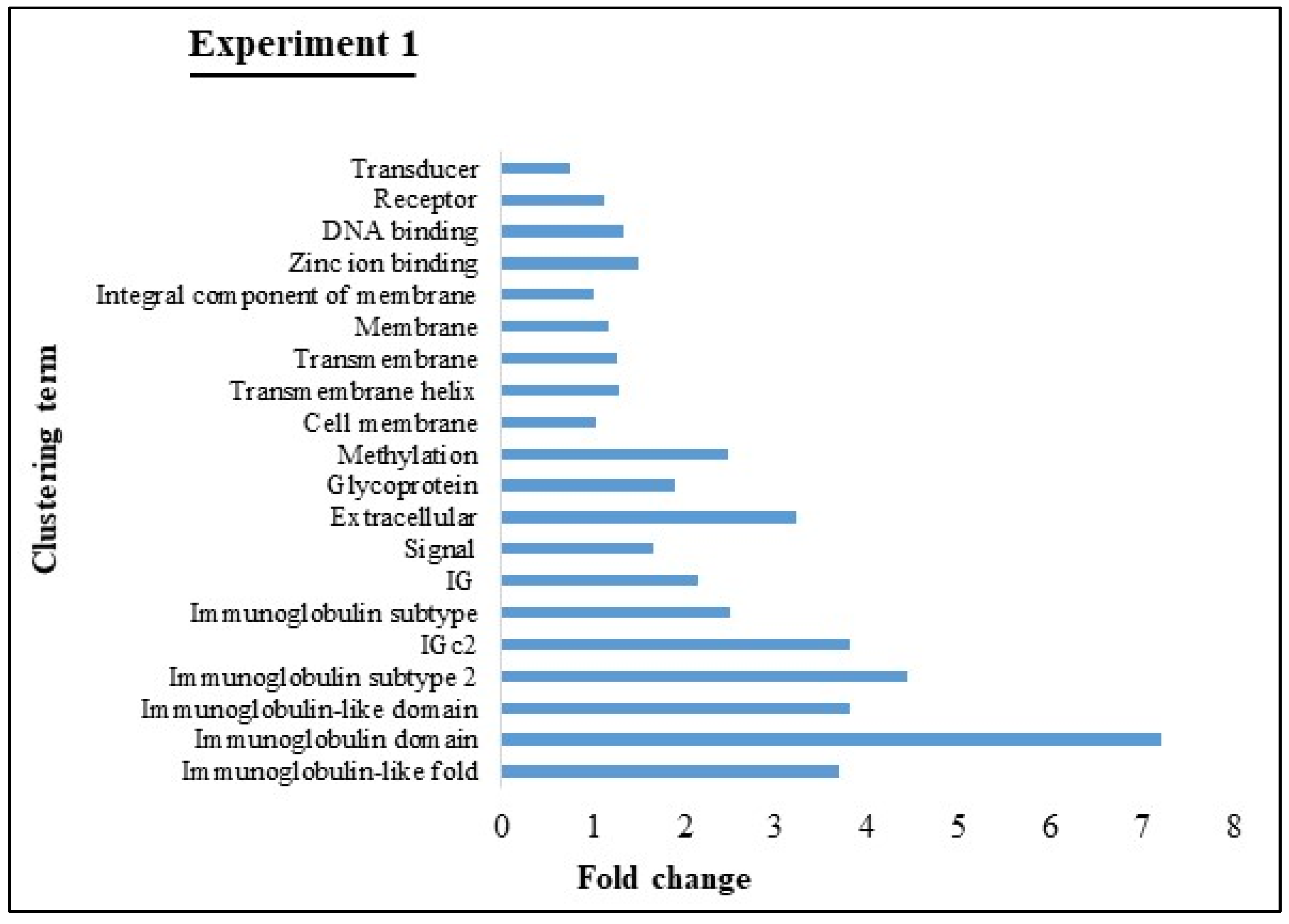
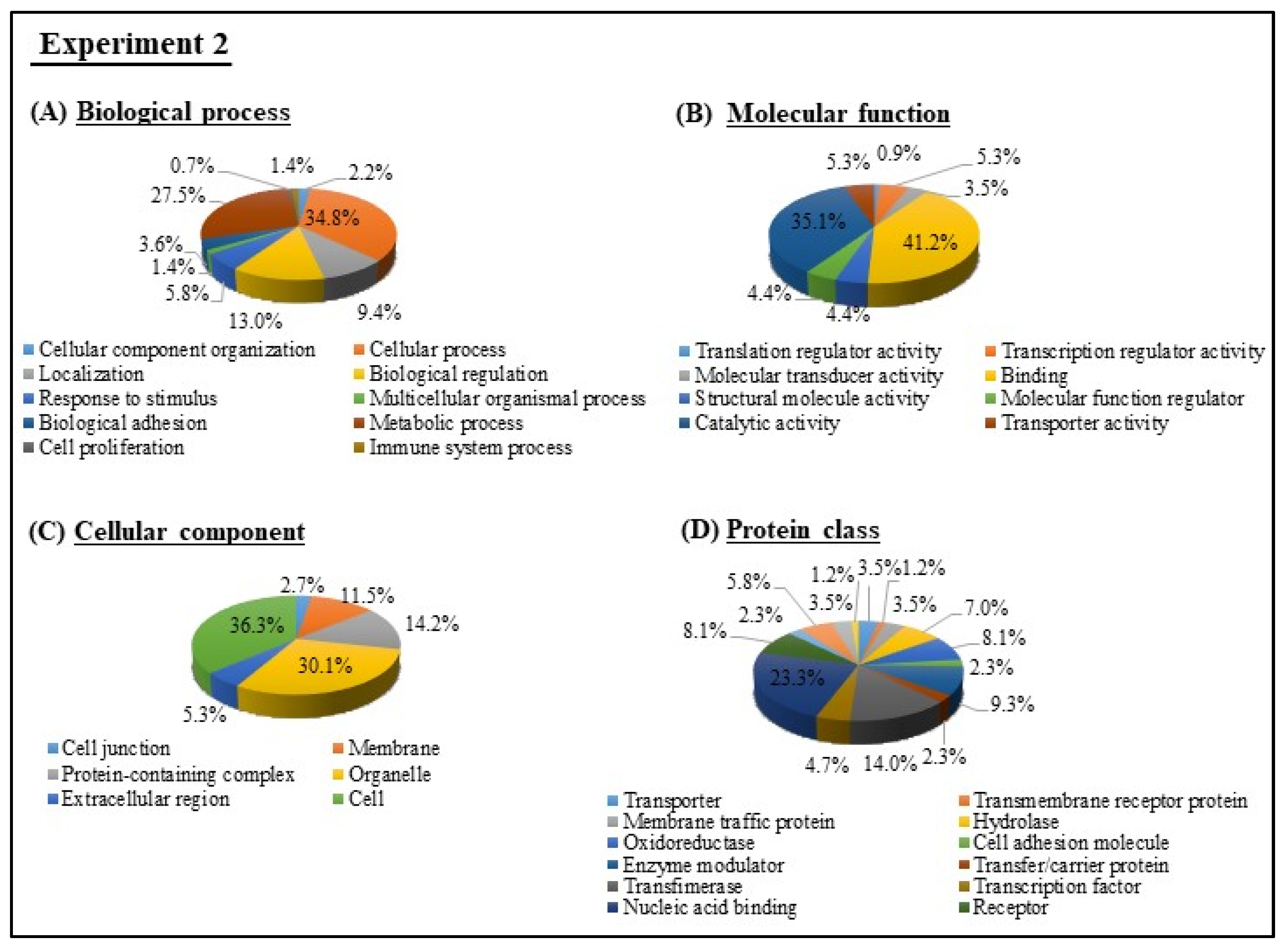
3.4. GO and KEGG Pathway analyses of Experiments 1 and 2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Cufaro, M.C.; Pieragostino, D.; Lanuti, P.; Rossi, C.; Cicalini, I.; Federici, L.; De Laurenzi, V.; Del Boccio, P. Extracellular Vesicles and Their Potential Use in Monitoring Cancer Progression and Therapy: The Contribution of Proteomics. J. Oncol. 2019, 2019, 1639854. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefirld, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumor growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Chen, T.; Xi, Q.-Y.; Sun, J.; Ye, R.-S.; Cheng, X.; Sun, R.-P.; Wang, S.; Shu, G.; Wang, L.-N.; Zhu, X.; et al. Revelation of mRNAs and proteins in porcine milk exosomes by transcriptomic and proteomic analysis. BMC Vet. Res. 2017, 13, 101. [Google Scholar] [CrossRef]
- Izumi, H.; Tsuda, M.; Sato, Y.; Kosaka, N.; Ochiya, T.; Iwamoto, H.; Namba, K.; Takeda, Y. Bovine milk exosomes contain microRNA and mRNA and are taken up by human macrophages. J. Dairy Sci. 2015, 98, 2920–2933. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.-J.; Aswath, K.; Schroeder, S.G.; Lippolis, J.; Reinhardt, T.; Sonstegard, T.S. MicroRNA expression profiles of bovine milk exosomes in response to Staphylococcus aureus infection. BMC Genom. 2015, 16, 806. [Google Scholar] [CrossRef] [Green Version]
- Gillet, N.A.; Florins, A.; Boxus, M.; Burteau, C.; Nigro, A.; Vandermeers, F.; Balon, H.; Bouzar, A.-B.; Defoiche, J.; Burny, A.; et al. Mechanisms of leukemogenesis induced by bovine leukemia virus: Prospects for novel anti-retroviral therapies in human. Retrovirology 2007, 4, 18. [Google Scholar] [CrossRef] [Green Version]
- Burny, A.; Cleuter, Y.; Kettmann, R.; Mammerickx, M.; Marbaix, G.; Portetelle, D.; Broeke, A.V.D.; Willems, L.; Thomas, R. Bovine Leukemia: Facts and Hypotheses Derived from the Study of an Infectious Cancer. Adv. Vet. Sci. Comp. Med. 1988, 32, 149–170. [Google Scholar] [CrossRef] [PubMed]
- Manet, G.; Guilbert, X.; Roux, A.; Vuillaume, A.; Parodi, A. Natural mode of horizontal transmission of bovine leukemia virus (BLV): The potential role of tabanids (Tabanus spp.). Vet. Immunol. Immunopathol. 1989, 22, 255–263. [Google Scholar] [CrossRef]
- Lassauzet, M.L.G.; Thurmond, M.C.; Johnson, W.O.; Stevens, F.; Picanso, J.P. Effect of brucellosis vaccination and dehorning on transmission of bovine leukemia virus in heifers on a Califolnia dairy. Can. J. Vet. Res. 1990, 54, 184–189. [Google Scholar] [PubMed]
- Kohara, J.; Konnai, S.; Onuma, M. Experimental transmission of Bovine leukemia virus in cattle via rectal palpation. Jpn. J. Vet. Res. 2006, 54, 25–30. [Google Scholar]
- Murakami, K.; Kobayashi, S.; Konishi, M.; Kameyama, K.-I.; Tsutsui, T. Nationwide survey of bovine leukemia virus infection among dairy and beef breeding cattle in Japan from 2009-2011. J. Vet. Med. Sci. 2013, 75, 1123–1126. [Google Scholar] [CrossRef] [Green Version]
- Nuotio, L.; Rusanen, H.; Sihvonen, L.; Neuvonen, E. Eradication of enzootic bovine leukosis from Finland. Prev. Vet. Med. 2003, 59, 43–49. [Google Scholar] [CrossRef]
- Mammerickx, M.; Lorenz, R.J.; Straub, O.C.; Donnelly, W.J.; Flensburg, J.C.; Gentile, G.; Markson, L.M.; Ressang, A.A.; Taylor, S.M. Bovine hematology. III. Comparative breed studies on the leukocyte parameters of several European cattle breeds as determined in the common reference laboratory. Vet. Med. B 1978, 25, 257–267. [Google Scholar]
- Fechner, H.; Blankenstein, P.; Looman, A.C.; Elwert, J.; Geue, L.; Albrecht, C.; Kurg, A.; Beier, D.; Marquardt, O.; Ebner, D. Provirus Variants of the Bovine Leukemia Virus and Their Relation to the Serological Status of Naturally Infected Cattle. Virology 1997, 237, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Murakami, K.; Okada, K.; Ikawa, Y.; Aida, Y. Bovine Leukemia Virus Induces CD5- B Cell Lymphoma in Sheep Despite Temporarily Increasing CD5+ B Cells in Asymptomatic Stage. Virology 1994, 202, 458–465. [Google Scholar] [CrossRef]
- Mammerickx, M.; Portelle, D.; de Clercq, K.; Burny, A. Experimental transmission of enzootic bovine leucosis to cattle, sheep and goats: Infectious doses of blood and incubation period of the disease. Leuk. Res. 1987, 11, 353–358. [Google Scholar] [CrossRef]
- Miura, S.; Inokuma, H. Evaluation of lactate dehydrogenase activity as an onset marker for enzootic bovine leukemia. Jpn. J. Large Anim. Clin. 2016, 6, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, M.; Shimizu, K.; Rahman, M.; Ishikawa, H.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Efficient method for isolation of exosomes from raw bovine milk. Drug Dev. Ind. Pharm. 2018, 45, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Shimizu, K.; Yamauchi, M.; Takase, H.; Ugawa, S.; Okada, A.; Inoshima, Y. Acidification effects on isolation of extracellular vesicles from bovine milk. PLoS ONE 2019, 14, e0222613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Li, M.; Aliotta, J.M.; Asara, J.M.; Tucker, L.; Quesenberry, P.; Lally, M.; Ramratnam, B. Quantitative proteomic analysis of exosomes from HIV-1-infected lymphocytic cells. Proteomics 2012, 12, 2203–2211. [Google Scholar] [CrossRef]
- Raab-Traub, N.; Dittmer, D.P. Viral effects on the content and function of extracellular vesicles. Nat. Rev. Genet. 2017, 15, 559–572. [Google Scholar] [CrossRef]
- Garg, M.; Braunstein, G.D.; Koeffler, H.P. LAMC2 as a therapeutic target for cancers. Expert Opin. Ther. Targets 2014, 18, 979–982. [Google Scholar] [CrossRef] [Green Version]
- Moon, Y.W.; Rao, G.; Kim, J.J.; Shim, H.S.; Park, K.-S.; An, S.S.; Kim, B.; Steeg, P.S.; Sarfaraz, S.; Lee, L.C.; et al. LAMC2 enhances the metastatic potential of lung adenocarcinoma. Cell Death Differ. 2015, 22, 1341–1352. [Google Scholar] [CrossRef] [Green Version]
- Hicklin, D.J.; Marincola, F.M.; Ferrone, S. HLA class I antigen downregulation in human cancers: T-cell immunotherapy revives an old story. Mol. Med. Today 1999, 5, 178–186. [Google Scholar] [CrossRef]
- Mahamdallie, S.; Yost, S.; Poyastro-Pearson, E.; Holt, E.; Zachariou, A.; Seal, S.; Elliott, A.; Clarke, M.; Warren-Perry, M.; Hanks, S.; et al. Identification of new Wilms tumour predisposition genes: An exome sequencing study. Lancet Child Adolesc. Health 2019, 3, 322–331. [Google Scholar] [CrossRef] [Green Version]
- Song, D.H.; Ko, G.H.; Lee, J.H.; Lee, J.S.; Lee, G.-W.; Kim, H.C.; Yang, J.W.; Heo, R.W.; Roh, G.S.; Han, S.-Y.; et al. Myoferlin expression in non-small cell lung cancer: Prognostic role and correlation with VEGFR-2 expression. Oncol. Lett. 2015, 11, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-S.; Liu, X.-H.; Liu, L.; Lou, W.-H.; Jin, D.; Yang, P.-Y.; Wang, X.-L. iTRAQ-based quantitative proteomics reveals myoferlin as a novel prognostic predictor in pancreatic adenocarcinoma. J. Proteom. 2013, 91, 453–465. [Google Scholar] [CrossRef] [PubMed]
- An, H.J.; Song, D.H.; Koh, H.M.; Kim, Y.-M.; Ko, G.H.; Lee, J.-H.; Lee, J.S.; Yang, J.W.; Kim, M.H.; Seo, D.H.; et al. Myoferlin silencing inhibits VEGFR2-mediated proliferation of metastatic clear cell renal cell carcinoma. Sci. Rep. 2019, 9, 12656–12658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheishvili, D.; Stefanska, B.; Yi, C.; Li, C.C.; Yu, P.; Arakelian, A.; Tanvir, I.; Khan, H.A.; Rabbani, S.; Szyf, M. A common promoter hypomethylation signature in invasive breast, liver and prostate cancer cell lines reveals novel targets involved in cancer invasiveness. Oncotarget 2015, 6, 33253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, C.; Qin, L.; Li, D.; Zhou, G.; Dang, D.; Chen, S.; Sun, T.; Zhang, R.; Wu, W.; et al. Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq. Viruses 2018, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, N.A.; Calabrese, C.; Malapelle, U.; Pellino, G.; De Stefano, A.; Sepe, R.; Sgariglia, R.; Quintavalle, C.; Federico, A.; Bianco, A.; et al. UbcH10 expression can predict prognosis and sensitivity to the antineoplastic treatment for colorectal cancer patients. Mol. Carcinog. 2015, 55, 793–807. [Google Scholar] [CrossRef]
- Matsumoto, A.; Ishibashi, Y.; Urashima, M.; Omura, N.; Nakada, K.; Nishikawa, K.; Shida, A.; Takada, K.; Kashiwagi, H.; Yanaga, K. High UBCH10 protein expression as a marker of poor prognosis in esophageal squamous cell carcinoma. Anticancer Res. 2014, 34, 955–962. [Google Scholar]
- Mo, C.; Gao, L.; Zhu, X.-F.; Wei, K.-L.; Zeng, J.-J.; Chen, G.; Feng, Z.-B. The clinicopathological significance of UBE2C in breast cancer: A study based on immunohistochemistry, microarray and RNA-sequencing data. Cancer Cell Int. 2017, 17, 83. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.-K.; Wu, W.-G.; Chen, L.; Dong, P.; Gu, J.; Mu, J.-S.; Yang, J.-H.; Liu, Y.-B. Expression of UbcH10 in pancreatic ductal adenocarcinoma and its correlation with prognosis. Tumor Biol. 2013, 34, 1473–1477. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, S.; Li, X.; Ji, Y.; Wang, Z.; Liu, C. UBE2C promotes rectal carcinoma via miR-381. Cancer Ther. Biol. 2018, 19, 230–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zou, J.; Yin, Y.; Zhang, B.; Hu, Y.; Wang, J.; Mu, H. Bioinformatic analysis identifies potentially key differentially expressed genes in oncogenesis and progression of clear cell renal cell carcinoma. PeerJ 2019, 7, e8096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Yu, G.; Liu, L.; Wang, J.; Chen, X.; Bian, Y.; Ji, Y.; Zhou, X.; Chen, Y.; et al. UBE2C Is a Potential Biomarker of Intestinal-Type Gastric Cancer With Chromosomal Instability. Front. Pharmacol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Canales, S.; De Rodas, M.L.; Nuncia, M.; Páez, R.; Amir, E.; Győrffy, B.; Pandiella, A.; Galán-Moya, E.M.; Ocaña, A. Functional transcriptomic annotation and protein–protein interaction analysis identify EZH2 and UBE2C as key upregulated proteins in ovarian cancer. Cancer Med. 2018, 7, 1896–1907. [Google Scholar] [CrossRef]
- Ieta, K.; Ojima, E.; Tanaka, F.; Nakamura, Y.; Haraguchi, N.; Mimori, K.; Inoue, H.; Kuwano, H.; Mori, M. Identification of overexpressed genes in hepatocellular carcinoma, with special reference toubiquitin-conjugating enzyme E2Cgene expression. Int. J. Cancer 2007, 121, 33–38. [Google Scholar] [CrossRef]
- Armogida, S.A.; Yannaras, N.M.; Melton, A.L.; Srivastava, M.D. Identification and quantification of innate immune system mediators in human breast milk. Allergy Asthma Proc. 2004, 25, 297–304. [Google Scholar]
- Zhang, N.; Zhang, S. Identification of differentially expressed genes between primary lung cancer and lymph node metastasis via bioinformatic analysis. Oncol. Lett. 2019, 18, 3754–3768. [Google Scholar] [CrossRef] [Green Version]
- Kuang, M.; Tao, X.; Peng, Y.; Zhang, W.; Pan, Y.; Cheng, L.; Yuan, C.; Zhao, Y.; Mao, H.; Zhuge, L.; et al. Proteomic analysis of plasma exosomes to differentiate malignant from benign pulmonary nodules. Clin. Proteom. 2019, 16, 5–11. [Google Scholar] [CrossRef]
- Brym, P.; Kaminski, S. Microarray analysis of differential gene expression profiles in blood cells of naturally BLV-infected and uninfected Holstein-Friesian cows. Mol. Biol. Rep. 2016, 44, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, K.; Nagata, Y.; Kitanaka, A.; Shiraishi, Y.; Shimamura, T.; Yasunaga, J.-I.; Totoki, Y.; Chiba, K.; Sato-Otsubo, A.; Nagae, G.; et al. Integrated molecular analysis of adult T cell leukemia/lymphoma. Nat. Genet. 2015, 47, 1304–1315. [Google Scholar] [CrossRef]
- Rosewick, N.; Durkin, K.; Artesi, M.; Marçais, A.; Hahaut, V.; Griebel, P.; Arsic, N.; Avettand-Fenoel, V.; Burny, A.; Charlier, C.; et al. Cis-perturbation of cancer drivers by the HTLV-1/BLV proviruses is an early determinant of leukemogenesis. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cattle no. | Age ※2 (Month) | ELISA ※3 (Antibody) | Nested PCR | Proviral Load ※4 | WBC ※5 (/µL) | Lymphocyte (/µL) | Key of EC ※6 | LDH ※7 Total (IU/l) | LDH Isozyme (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 2 + 3 | 4 | 5 | ||||||||||
| Experiment 1 | Uninfected cattle | ||||||||||||||
| U1 | 59.0 | - | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | |
| U2 | 57.7 | - | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | |
| U3 | 94.8 | - | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | |
| U4 | 49.8 | - | - | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | NT | |
| BLV-infected cattle with HPL※8 | |||||||||||||||
| B1 | 74.0 | + | NT | 15,405 | 15,300 | 12,393 | + | NT | NT | NT | NT | NT | NT | NT | |
| B2 | 78.5 | + | NT | 14,571 | 12,900 | 8127 | + | NT | NT | NT | NT | NT | NT | NT | |
| B3 | 107.6 | + | NT | 13,918 | 11,200 | 6720 | ± | NT | NT | NT | NT | NT | NT | NT | |
| B4 | 53.3 | + | NT | 31,093 | 16,500 | 11,880 | + | NT | NT | NT | NT | NT | NT | NT | |
| Experiment 2 | Uninfected cattle with LLDH※9 | ||||||||||||||
| U1 | 33.6 | - | - | NT | 7100 | 2592 | - | 963 | 70.6 | 15.0 | 9.4 | 24.4 | 3.3 | 1.7 | |
| U2 | 43.2 | - | - | NT | 8800 | 3142 | - | 729 | 72.3 | 14.5 | 8.0 | 22.5 | 3.2 | 2.0 | |
| U3 | 37.1 | - | - | NT | 9700 | 4113 | - | 1062 | 75.6 | 15.0 | 6.1 | 21.1 | 2.0 | 1.3 | |
| U4 | 33.5 | - | - | NT | 7800 | 4547 | - | 821 | 69.7 | 12.9 | 11.0 | 23.9 | 4.1 | 2.3 | |
| BLV-infected cattle with HPL + HLDH※10 | |||||||||||||||
| B1 | 24.8 | + | + | 10,264 | 11,900 | 6593 | - | 909 | 48.1 | 29.4 | 15.0 | 44.4 | 5.1 | 2.4 | |
| B2 | 38.5 | + | + | 5883 | 8000 | 4592 | - | 901 | 51.9 | 25.6 | 15.4 | 41.0 | 5.0 | 2.1 | |
| B3 | 23.5 | + | + | 6550 | 16,200 | 8699 | - | 918 | 52.4 | 27.5 | 14.2 | 41.7 | 4.0 | 1.9 | |
| B4 | 22.6 | + | + | 10,781 | 15,000 | 8940 | - | 819 | 51.4 | 25.3 | 15.4 | 40.7 | 5.5 | 2.4 | |
| KEGG Pathway Name | Gene List | Gene No. |
|---|---|---|
| Pathways in cancer | AXIN1, LAMC2, PIK3CB, FGF22 | 4 |
| PI3K-Akt signaling pathway | MYB, LAMC2, PIK3CB, FGF22 | 4 |
| Metabolic pathways | COX2, PGAM2, PIK3CB, HSD17B6 | 4 |
| MAPK signaling pathway | CACNA1A, FGF22, PTPN5 | 3 |
| Cell adhesion molecules | PVRL1, BLA-DQB, IGSF11 | 3 |
| Ras signaling pathway | PIK3CB, FGF22, RASAL3 | 3 |
| Phagosome | DYNC1H1, BLA-DQB | 2 |
| B cell receptor signaling pathway | CD79A, PIK3CB | 2 |
| HTLV-1 infection ※ | PIK3CB, BLA-DQB | 2 |
| KEGG Pathway Name | Gene List | Gene No. |
|---|---|---|
| Metabolic pathways | NNT, CKMT1B, SCP2, ISYNA1, GNS, NADSYN1, ALDH8A1, SLC33A1, AMPD3, ENTPD4, ELOVL6, DHCR24, MGAT1, ALAS1, ELOVL5 | 15 |
| Pathways in cancer | BAX, LPAR1, CTBP2, GNAS, TGFBR1, JUP, BAK1, DDB2 | 8 |
| Human papillomavirus infection | BAX, GNAS, PPP2R5B, BAK1, TBPL1 | 5 |
| Protein processing in endoplasmic reticulum | BAX, HSPA1A, PRKCSH, EDEM2; ER, BAK1 | 5 |
| Influenza A | BAX, TLR4, TRIM25, TMPRSS2, BAK1 | 5 |
| Chronic myeloid leukemia | BAX, CTBP2, TGFBR1, BAK1, DDB2 | 5 |
| HTLV-1 infection | BAX, ITGB2, TGFBR1, CDC20, TBPL1; | 5 |
| Viral carcinogenesis | BAX, BAK1, CDC20, TBPL1 | 4 |
| HIV-1 infection ※ | BAX, TLR4, CCNB1, BAK1 | 4 |
| Rap1 signaling pathway | LPAR1, GNAS, ITGB2 | 3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, H.; Rahman, M.M.; Yamauchi, M.; Takashima, S.; Wakihara, Y.; Kamatari, Y.O.; Shimizu, K.; Okada, A.; Inoshima, Y. mRNA Profile in Milk Extracellular Vesicles from Bovine Leukemia Virus-Infected Cattle. Viruses 2020, 12, 669. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060669
Ishikawa H, Rahman MM, Yamauchi M, Takashima S, Wakihara Y, Kamatari YO, Shimizu K, Okada A, Inoshima Y. mRNA Profile in Milk Extracellular Vesicles from Bovine Leukemia Virus-Infected Cattle. Viruses. 2020; 12(6):669. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060669
Chicago/Turabian StyleIshikawa, Hinata, Md. Matiur Rahman, Marika Yamauchi, Shigeo Takashima, Yoshiko Wakihara, Yuji O. Kamatari, Kaori Shimizu, Ayaka Okada, and Yasuo Inoshima. 2020. "mRNA Profile in Milk Extracellular Vesicles from Bovine Leukemia Virus-Infected Cattle" Viruses 12, no. 6: 669. https://0-doi-org.brum.beds.ac.uk/10.3390/v12060669