Temperature Sensitivity: A Potential Method for the Generation of Vaccines against the Avian Coronavirus Infectious Bronchitis Virus

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. In Vivo Characterisation
2.3. Virus Re-Isolation in Embryonated Hens’ Eggs
2.4. In Vitro Growth Kinetic Experiments
2.5. Analysis of sgmRNA Synthesis by Quantitative Real-Time PCR (qRT-PCR)
2.6. Temperature Swap Assays
2.7. Co-Infection Assay
2.8. Analysis of Protein Expression by Western Blot
2.9. Statistics
3. Results
3.1. Replication of rIBV Beau-R is Restricted In Vivo
3.2. Replication of rIBV Beau-R In Vitro Is Sensitive to Temperature
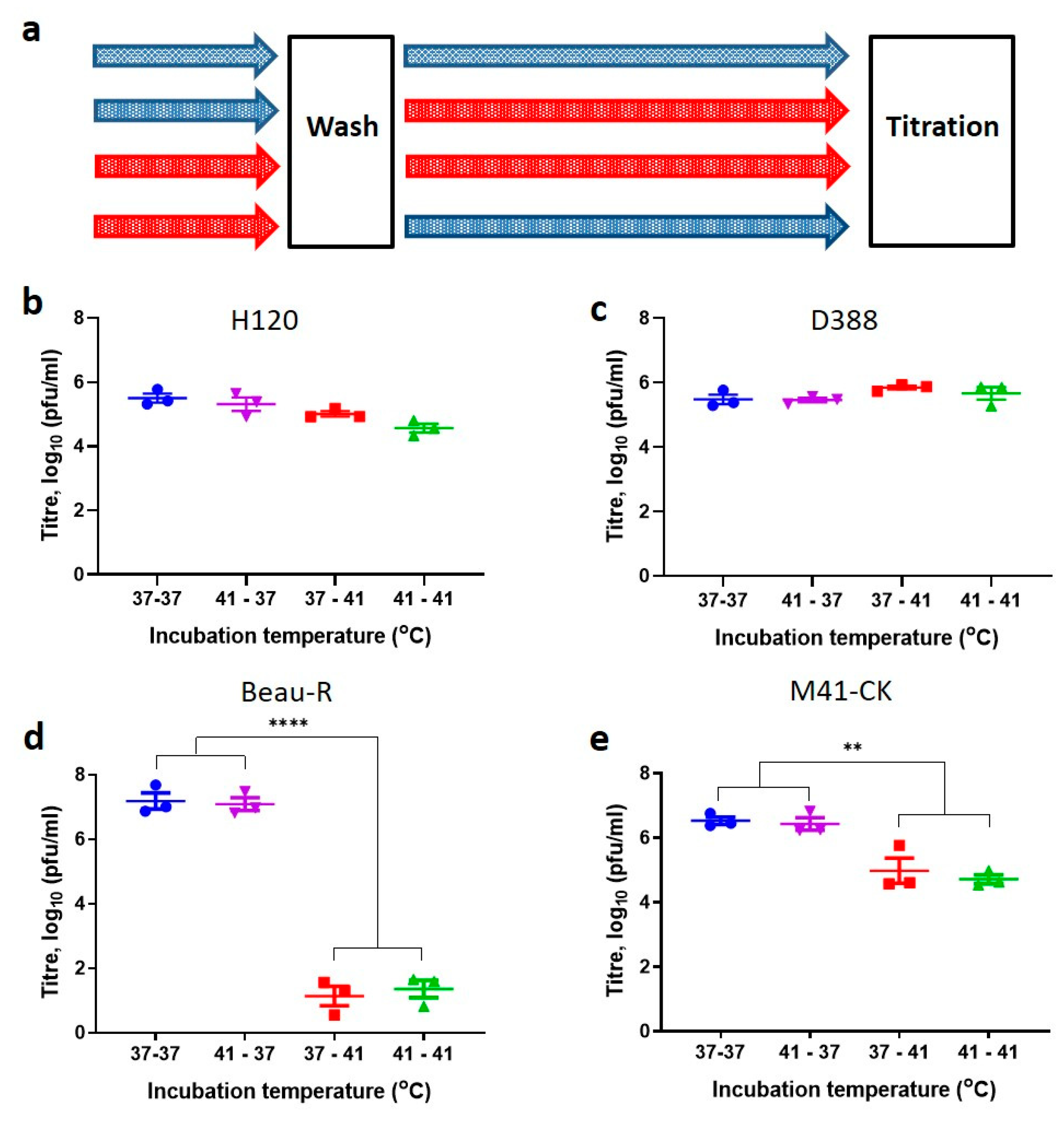
3.3. Temperature Sensitivity Is Not a Shared Characteristic of Attenuated Strains
3.4. The Nephropathogenic IBV Strain D388(QX) Is Able to Replicate at 41 °C
3.5. Entry at Permissive Temperatures Cannot Recover Replication at Non-Permissive Temperatures
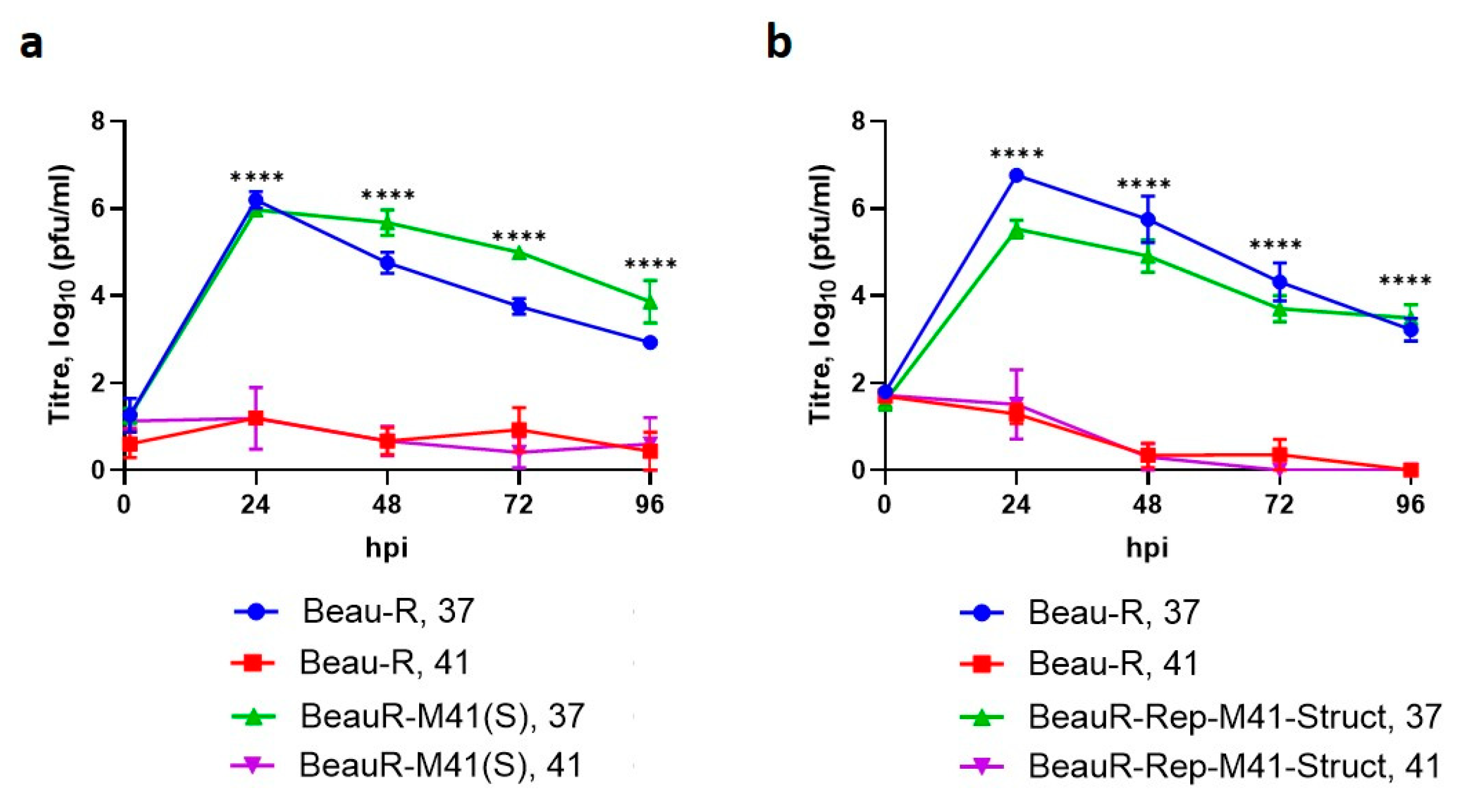
3.6. The Spike Glycoprotein from M41-CK Cannot Rescue Beau-R Replication at 41 °C
3.7. The Replicase Gene Is a Determinant of Temperature Sensitivity
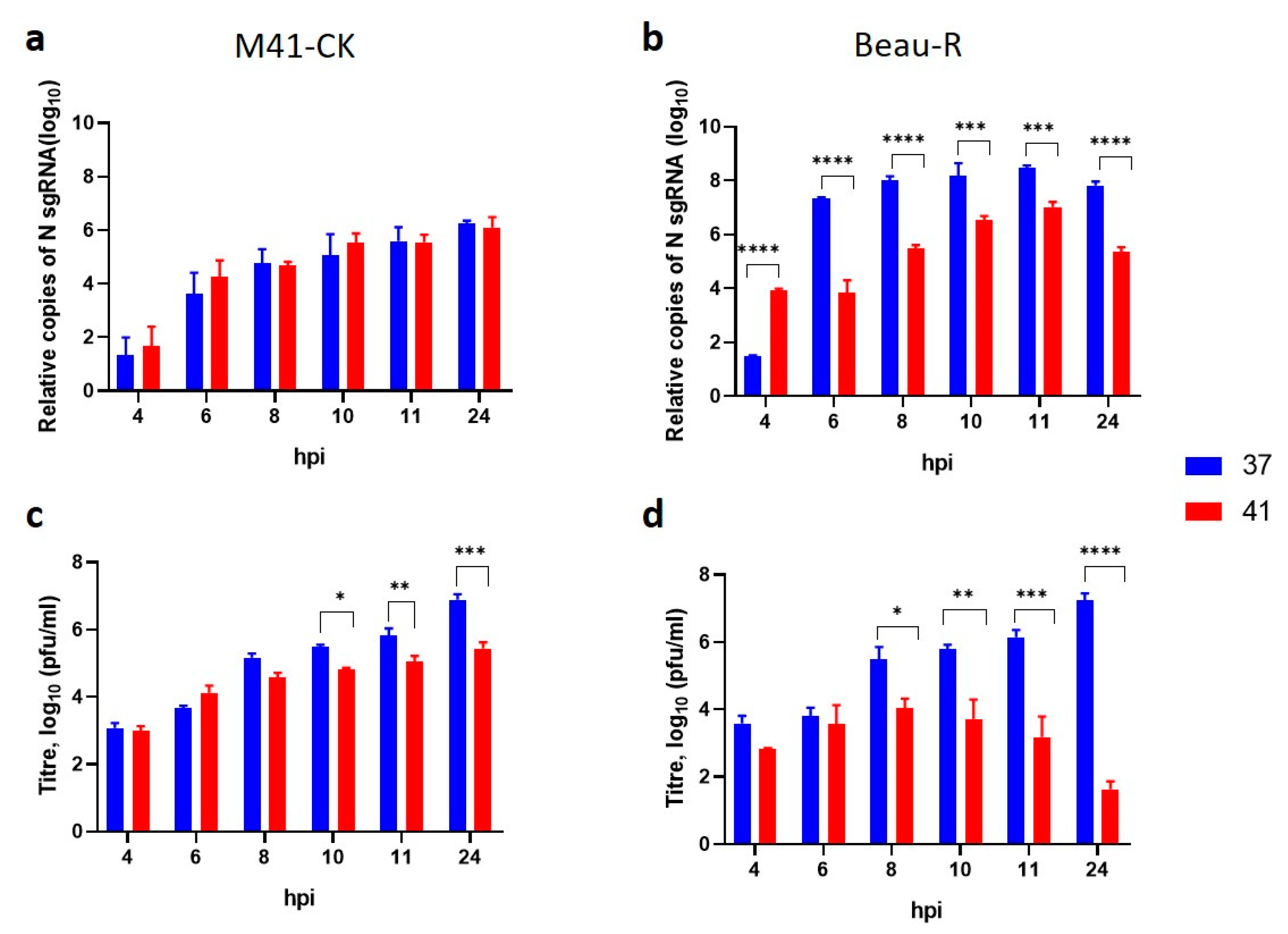
3.8. Temperature Affects sgRNA Levels during Beau-R Infection
3.9. Beau-R Replication at 41 °C Can Be Rescued by M41-CK
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rota, P.A.; Oberste, M.S.; Monroe, S.S.; Nix, W.A.; Campagnoli, R.; Icenogle, J.P.; Peñaranda, S.; Bankamp, B.; Maher, K.; Chen, M.-H.; et al. Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 2003, 300, 1394–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. MMBR 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.X.; Ng, Y.L.; Tam, J.P.; Liu, D.X. Human Coronaviruses: A Review of Virus-Host Interactions. Diseases 2016, 4, 26. [Google Scholar] [CrossRef]
- Laude, H.; Van Reeth, K.; Pensaert, M. Porcine respiratory coronavirus: Molecular features and virus-host interactions. Vet. Res. 1993, 24, 125–150. [Google Scholar]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Bejo, M.H.; Abubakar, M.S.; Abba, Y. Pathogenesis and Diagnostic Approaches of Avian Infectious Bronchitis. Adv. Virol. 2016, 2016, 4621659. [Google Scholar] [CrossRef] [Green Version]
- de Wit, J.J.; Cook, J.K.; van der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef]
- Cook, J.K.; Chesher, J.; Baxendale, W.; Greenwood, N.; Huggins, M.B.; Orbell, S.J. Protection of chickens against renal damage caused by a nephropathogenic infectious bronchitis virus. Avian Pathol. 2001, 30, 423–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayry, J.; Goudar, M.S.; Nighot, P.K.; Kshirsagar, S.G.; Ladman, B.S.; Gelb, J., Jr.; Ghalsasi, G.R.; Kolte, G.N. Emergence of a nephropathogenic avian infectious bronchitis virus with a novel genotype in India. J. Clin. Microbiol. 2005, 43, 916–918. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Kong, X. A new genotype of nephropathogenic infectious bronchitis virus circulating in vaccinated and non-vaccinated flocks in China. Avian Pathol. 2004, 33, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Bijlenga, G.; Cook, J.K.A.; Gelb, J.; de Wit, J.J., Jr. Development and use of the H strain of avian infectious bronchitis virus from the Netherlands as a vaccine: A review. Avian Pathol. 2004, 33, 550–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerligs, H.J.; Boelm, G.J.; Meinders, C.A.; Stuurman, B.G.; Symons, J.; Tarres-Call, J.; Bru, T.; Vila, R.; Mombarg, M.; Karaca, K.; et al. Efficacy and safety of an attenuated live QX-like infectious bronchitis virus strain as a vaccine for chickens. Avian Pathol. 2011, 40, 93–102. [Google Scholar] [CrossRef]
- Oade, M.S.; Keep, S.; Freimanis, G.L.; Orton, R.J.; Britton, P.; Hammond, J.A.; Bickerton, E. Attenuation of Infectious Bronchitis Virus in Eggs Results in Different Patterns of Genomic Variation across Multiple Replicates. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, T.; Casais, R.; Dove, B.; Britton, P.; Cavanagh, D. Recombinant infectious bronchitis coronavirus Beaudette with the spike protein gene of the pathogenic M41 strain remains attenuated but induces protective immunity. J. Virol. 2004, 78, 13804–13811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armesto, M.; Evans, S.; Cavanagh, D.; Abu-Median, A.B.; Keep, S.; Britton, P. A recombinant avian infectious bronchitis virus expressing a heterologous spike gene belonging to the 4/91 serotype. PLoS ONE 2011, 6, e24352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Beurden, S.J.; Berends, A.J.; Kramer-Kuhl, A.; Spekreijse, D.; Chenard, G.; Philipp, H.C.; Mundt, E.; Rottier, P.J.M.; Verheije, M.H. Recombinant live attenuated avian coronavirus vaccines with deletions in the accessory genes 3ab and/or 5ab protect against infectious bronchitis in chickens. Vaccine 2018, 36, 1085–1092. [Google Scholar] [CrossRef]
- Jiang, Y.; Cheng, X.; Zhao, X.; Yu, Y.; Gao, M.; Zhou, S. Recombinant infectious bronchitis coronavirus H120 with the spike protein S1 gene of the nephropathogenic IBYZ strain remains attenuated but induces protective immunity. Vaccine 2020, 38, 3157–3168. [Google Scholar] [CrossRef]
- Casais, R.; Thiel, V.; Siddell, S.G.; Cavanagh, D.; Britton, P. Reverse genetics system for the avian coronavirus infectious bronchitis virus. J. Virol. 2001, 75, 12359–12369. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.; Keep, S.; Britton, P.; de Wit, S.; Bickerton, E.; Vervelde, L. Recombinant Infectious Bronchitis Viruses Expressing Chimeric Spike Glycoproteins Induce Partial Protective Immunity against Homologous Challenge despite Limited Replication In Vivo. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Geilhausen, H.E.; Ligon, F.B.; Lukert, P.D. The pathogenesis of virulent and avirulent avian infectious bronchitis virus. Arch. Gesamte Virusforsch. 1973, 40, 285–290. [Google Scholar] [CrossRef]
- Ammayappan, A.; Upadhyay, C.; Gelb, J., Jr.; Vakharia, V.N. Identification of sequence changes responsible for the attenuation of avian infectious bronchitis virus strain Arkansas DPI. Arch. Virol. 2009, 154, 495–499. [Google Scholar] [CrossRef]
- Armesto, M.; Cavanagh, D.; Britton, P. The replicase gene of avian coronavirus infectious bronchitis virus is a determinant of pathogenicity. PLoS ONE 2009, 4, e7384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.-F.; Huang, Q.-H.; Lu, M.; Wu, J.-Q.; Lin, S.-Q.; Zhu, F.; Zhang, X.-M.; Huang, Y.Y.; Yang, S.-H.; Xu, C.-T. Attenuation mechanism of virulent infectious bronchitis virus strain with QX genotype by continuous passage in chicken embryos. Vaccine 2016, 34, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, K.K.; Cervantes-Barragan, L.; Ludewig, B.; Thiel, V. Mouse hepatitis virus liver pathology is dependent on ADP-ribose-1”-phosphatase, a viral function conserved in the alpha-like supergroup. J. Virol. 2008, 82, 12325–12334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, R.L.; Becker, M.M.; Eckerle, L.D.; Bolles, M.; Denison, M.R.; Baric, R.S. A live, impaired-fidelity coronavirus vaccine protects in an aged, immunocompromised mouse model of lethal disease. Nat. Med. 2012, 18, 1820–1826. [Google Scholar] [CrossRef] [Green Version]
- Fehr, A.R.; Channappanavar, R.; Jankevicius, G.; Fett, C.; Zhao, J.; Athmer, J.; Meyerholz, D.K.; Ahel, I.; Perlman, S. The Conserved Coronavirus Macrodomain Promotes Virulence and Suppresses the Innate Immune Response during Severe Acute Respiratory Syndrome Coronavirus Infection. mBio 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Menachery, V.D.; Gralinski, L.E.; Mitchell, H.D.; Dinnon, K.H., 3rd; Leist, S.R.; Yount, B.L., Jr.; McAnarney, E.T.; Graham, R.L.; Waters, K.M.; Baric, R.S.; et al. Combination Attenuation Offers Strategy for Live Attenuated Coronavirus Vaccines. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Belshe, R.B.; Mendelman, P.M.; Treanor, J.; King, J.; Gruber, W.C.; Piedra, P.; Bernstein, D.I.; Hayden, F.G.; Kotloff, K.; Zangwill, K.; et al. The efficacy of live attenuated, cold-adapted, trivalent, intranasal influenzavirus vaccine in children. N. Engl. J. Med. 1998, 338, 1405–1412. [Google Scholar] [CrossRef]
- Juhasz, K.; Whitehead, S.S.; Boulanger, C.A.; Firestone, C.-Y.; Collins, P.L.; Murphy, B.R. The two amino acid substitutions in the L protein of cpts530/1009, a live-attenuated respiratory syncytial virus candidate vaccine, are independent temperature-sensitive and attenuation mutations. Vaccine 1999, 17, 1416–1424. [Google Scholar] [CrossRef]
- Foxman, E.F.; Storer, J.A.; Fitzgerald, M.E.; Wasik, B.R.; Hou, L.; Zhao, H.; Turner, P.E.; Pyle, A.M.; Iwasaki, A. Temperature-dependent innate defense against the common cold virus limits viral replication at warm temperature in mouse airway cells. Proc. Natl. Acad. Sci. USA 2015, 112, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Nogales, A.; Rodriguez, L.; Chauché, C.; Huang, K.; Reilly, E.C.; Topham, D.J.; Murcia, P.R.; Parrish, C.R.; Martínez-Sobrido, L. Temperature-Sensitive Live-Attenuated Canine Influenza Virus H3N8 Vaccine. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennion, R.; Hill, G. The Preparation of Chicken Kidney Cell Cultures for Virus Propagation. In Coronaviruses: Methods and Protocols 1282; Maier, H.J., Bickerton, E., Britton, P., Eds.; Springer: New York, NY, USA, 2015; pp. 57–62. [Google Scholar]
- Himly, M.; Foster, D.N.; Bottoli, I.; Iacovoni, J.S.; Vogt, P.K. The DF-1 chicken fibroblast cell line: Transformation induced by diverse oncogenes and cell death resulting from infection by avian leukosis viruses. Virology 1998, 248, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casais, R.; Dove, B.; Cavanagh, D.; Britton, P. Recombinant avian infectious bronchitis virus expressing a heterologous spike gene demonstrates that the spike protein is a determinant of cell tropism. J. Virol. 2003, 77, 9084–9089. [Google Scholar] [CrossRef] [Green Version]
- de Wit, J.J.; Nieuwenhuisen-van Wilgen, J.; Hoogkamer, A.; van de Sande, H.; Zuidam, G.J.; Fabri, T.H.F. Induction of cystic oviducts and protection against early challenge with infectious bronchitis virus serotype D388 (genotype QX) by maternally derived antibodies and by early vaccination. Avian Pathol. 2011, 40, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, D.; Ellis, M.M.; Cook, J.K.A. Relationship between sequence variation in the S1 spike protein of infectious bronchitis virus and the extent of cross-protection in vivo. Avian Pathol. 1997, 26, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.K.A.; Orbell, S.J.; Woods, M.A.; Huggins, M.B. Breadth of protection of the respiratory tract provided by different live-attenuated infectious bronchitis vaccines against challenge with infectious bronchitis viruses of heterologous serotypes. Avian Pathol. 1999, 28, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Keep, S.M.; Bickerton, E.; Britton, P. Transient dominant selection for the modification and generation of recombinant infectious bronchitis coronaviruses. Methods Mol. Biol. 2015, 1282, 115–133. [Google Scholar]
- Maier, H.J.; Hawes, P.C.; Cottam, E.M.; Mantell, J.; Verkade, P.; Monaghan, P.; Wileman, T.; Britton, P. Infectious bronchitis virus generates spherules from zippered endoplasmic reticulum membranes. mBio 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Pharmacopeia, E. Avian Infectious Bronchitis Vaccine (Live); European Directorate for the Quality of Medicines and HealthCare (EDQM), Council of Europe: Strasbourg, France, 2010. [Google Scholar]
- Keep, S.; Bickerton, E.; Armesto, M.; Britton, P. The ADRP domain from a virulent strain of infectious bronchitis virus is not sufficient to confer a pathogenic phenotype to the attenuated Beaudette strain. J. Gen. Virol. 2018, 99, 1097–1102. [Google Scholar] [CrossRef]
- Alford, R.H.; Kasel, J.A.; Gerone, P.J.; Knight, V. Human influenza resulting from aerosol inhalation. Proc. Soc. Exp. Biol. Med. 1966, 122, 800–804. [Google Scholar] [CrossRef]
- Nogales, A.; Martinez-Sobrido, L. Reverse Genetics Approaches for the Development of Influenza Vaccines. Int. J. Mol. Sci. 2016, 18, 20. [Google Scholar] [CrossRef]
- Crowe, J.E.; Bui, P.T.; Davis, A.R.; Chanock, R.M.; Murphy, B.R. A further attenuated derivative of a cold-passaged temperature-sensitive mutant of human respiratory syncytial virus retains immunogenicity and protective efficacy against wild-type challenge in seronegative chimpanzees. Vaccine 1994, 12, 783–790. [Google Scholar] [CrossRef]
- Buchholz, U.J.; Cunningham, C.K.; Muresan, P.; Gnanashanmugam, D.; Sato, P.; Siberry, G.K.; Rexroad, V.; Valentine, M.; Perlowski, C.; Schappell, E.; et al. Live Respiratory Syncytial Virus (RSV) Vaccine Candidate Containing Stabilized Temperature-Sensitivity Mutations Is Highly Attenuated in RSV-Seronegative Infants and Children. J. Infect. Dis. 2018, 217, 1338–1346. [Google Scholar] [CrossRef] [PubMed]
- McFarland, E.J.; Karron, R.A.; Muresan, P.; Cunningham, C.K.; Perlowski, C.; Libous, J.; Oliva, J.; Jean-Philippe, P.; Moye, J.; Schappell, E.; et al. Live-attenuated respiratory syncytial virus vaccine with M2-2 deletion and with SH non-coding region is highly immunogenic in children. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Dingle, J.G. Module 4: Gas exchange and thermoregulation. In Study Book: Poultry Husbandry 1; University of Southern Queensland: Toowoomba, Australia, 1990. [Google Scholar]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, E.R., Jr.; Pichurko, B.M.; Bowman, H.F.; Ingenito, E.; Burns, S.; Dowling, N.; Solway, J. Thermal mapping of the airways in humans. J. Appl. Physiol. 1985, 58, 564–570. [Google Scholar] [CrossRef]
- Lindemann, J.; Leiacker, R.; Rettinger, G.; Keck, T. Nasal mucosal temperature during respiration. Clin. Otolaryngol. Allied Sci. 2002, 27, 135–139. [Google Scholar] [CrossRef]
- Bradel-Tretheway, B.G.; Kelley, Z.; Chakraborty-Sett, S.; Takimoto, T.; Kim, B.; Dewhurst, S. The human H5N1 influenza A virus polymerase complex is active in vitro over a broad range of temperatures, in contrast to the WSN complex, and this property can be attributed to the PB2 subunit. J. Gen. Virol. 2008, 89, 2923–2932. [Google Scholar] [CrossRef]
- Massin, P.; Kuntz-Simon, G.; Barbezange, C.; Deblanc, C.; Oger, A.; Marquet-Blouin, E.; Marquet-Blouin, E.; Bougeard, S.; van der Werf, S.; Jestin, V. Temperature sensitivity on growth and/or replication of H1N1, H1N2 and H3N2 influenza A viruses isolated from pigs and birds in mammalian cells. Vet. Microbiol. 2010, 142, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Broadbent, A.J.; Santos, C.P.; Godbout, R.A.; Subbarao, K. The temperature-sensitive and attenuation phenotypes conferred by mutations in the influenza virus PB2, PB1, and NP genes are influenced by the species of origin of the PB2 gene in reassortant viruses derived from influenza A/California/07/2009 and A/WSN/33 viruses. J. Virol. 2014, 88, 12339–12347. [Google Scholar] [PubMed] [Green Version]
- Deng, X.; Mettelman, R.C.; O’Brien, A.; Thompson, J.A.; O’Brien, T.E.; Baker, S.C. Analysis of Coronavirus Temperature-Sensitive Mutants Reveals an Interplay between the Macrodomain and Papain-Like Protease Impacting Replication and Pathogenesis. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maassab, H.F. Adaptation and growth characteristics of influenza virus at 25 degrees c. Nature 1967, 213, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Maassab, H.F. Plaque Formation of Influenza Virus at 25 °C. Nature 1968, 219, 645–646. [Google Scholar] [CrossRef]
- Hall, S.L.; Stokes, A.; Tierney, E.L.; London, W.T.; Belshe, R.B.; Newman, F.C.; Murphy, B.R. Cold-passaged human parainfluenza type 3 viruses contain ts and non-ts mutations leading to attenuation in rhesus monkeys. Virus Res. 1992, 22, 173–184. [Google Scholar] [CrossRef]
- Ray, R.; Galinski, M.S.; Heminway, B.R.; Meyer, K.; Newman, F.K.; Belshe, R.B. Temperature-sensitive phenotype of the human parainfluenza virus type 3 candidate vaccine strain (cp45) correlates with a defect in the L gene. J. Virol. 1996, 70, 580–584. [Google Scholar] [CrossRef] [Green Version]
- Maassab, H.F.; Bryant, M.L. The development of live attenuated cold-adapted influenza virus vaccine for humans. Rev. Med. Virol. 1999, 9, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Lu, B.; Zhou, H.; Ma, C.; Zhao, J.; Yang, C.F.; Kemble, G.; Greenberg, H. Multiple amino acid residues confer temperature sensitivity to human influenza virus vaccine strains (FluMist) derived from cold-adapted A/Ann Arbor/6/60. Virology 2003, 306, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Sawicki, S.G.; Sawicki, D.L.; Younker, D.; Meyer, Y.; Thiel, V.; Stokes, H.; Siddel, S.G. Functional and Genetic Analysis of Coronavirus Replicase-Transcriptase Proteins. PLoS Pathog. 2005, 1, e39. [Google Scholar] [CrossRef]
- Schinkel, S.C.B.; Rubin, S.; Wright, K.E. Mechanisms of temperature sensitivity of attenuated Urabe mumps virus. Virus Res. 2017, 227, 104–109. [Google Scholar] [CrossRef]
- Yachida, S.; Aoyama, S.; Takahashi, N.; Iritani, Y.; Katagiri, K. Influence of temperature of incubation on chicken embryo tracheal organ cultures and chick embryos infected with strains of avian infectious bronchitis virus. Res. Vet. Sci. 1981, 31, 14–18. [Google Scholar] [CrossRef]
- Gelb, J.; Lunt, R.L.; Metz, A.L.; Fries, P.A. Attenuation of Avian Infectious Bronchitis Virus by Cold-Adaptation. Avian Dis. 1991, 35, 847–853. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.Y.; Lu, C.Y.; Shao, P.L.; Lee, P.I.; Lin, M.T.; Fan, T.Y.; Cheng, A.L.; Lee, W.L.; Hu, J.J.; Yeh, S.J.; et al. Viral infections associated with Kawasaki disease. J. Formos. Med. Assoc. Taiwan Yi Zhi 2014, 113, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielech, A.M.; Deng, X.; Chen, Y.; Kindler, E.; Wheeler, D.L.; Mesecar, A.D.; Theil, V.; Perlman, S.; Baker, S.C. Murine coronavirus ubiquitin-like domain is important for papain-like protease stability and viral pathogenesis. J. Virol. 2015, 89, 4907–4917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stobart, C.C.; Lee, A.S.; Lu, X.; Denison, M.R. Temperature-sensitive mutants and revertants in the coronavirus nonstructural protein 5 protease (3CLpro) define residues involved in long-distance communication and regulation of protease activity. J. Virol. 2012, 86, 4801–4810. [Google Scholar] [CrossRef] [Green Version]
- Sparks, J.S.; Donaldson, E.F.; Lu, X.; Baric, R.S.; Denison, M.R. A novel mutation in murine hepatitis virus nsp5, the viral 3C-like proteinase, causes temperature-sensitive defects in viral growth and protein processing. J. Virol. 2008, 82, 5999–6008. [Google Scholar] [CrossRef] [Green Version]
- V’kovski, P.; Gultom, M.; Steiner, S.; Kelly, J.; Russeil, J.; Mangeat, B.; Cora, E.; Pezoldt, J.; Holwerda, M.; Kratzel, A.; et al. Disparate temperature-dependent virus—Host dynamics for SARS-CoV-2 and SARS-CoV in the human respiratory epithelium. bioRxiv 2020. [Google Scholar] [CrossRef]
- Clementz, M.A.; Kanjanahaluethai, A.; O’Brien, T.E.; Baker, S.C. Mutation in murine coronavirus replication protein nsp4 alters assembly of double membrane vesicles. Virology 2008, 375, 118–129. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dpi | Beak | Eyelid | Trachea |
|---|---|---|---|---|
| Mock | 1 | 0/6 | 0/6 | 0/6 |
| Beau-R | 1 | 6/6 | 0/6 | 0/6 |
| M41-CK | 1 | 6/6 | 4/6 | 0/6 |
| Mock | 4 | 0/6 | 0/6 | 0/6 |
| Beau-R | 4 | 2/6 | 0/6 | 0/6 |
| M41-CK | 4 | 6/6 | 6/6 | 5/6 |
| Mock | 6 | 0/6 | 0/6 | 0/6 |
| Beau-R | 6 | 0/6 | 0/6 | 1/6 |
| M41-CK | 6 | 0/6 | 6/6 | 5/6 |
| Mock | 7 | 0/6 | 0/6 | 0/6 |
| Beau-R | 7 | 2/4 | 1/4 | 0/4 |
| M41-CK | 7 | 2/4 * | 5/6 | 2/5 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keep, S.; Stevenson-Leggett, P.; Steyn, A.; Oade, M.S.; Webb, I.; Stuart, J.; Vervelde, L.; Britton, P.; Maier, H.J.; Bickerton, E. Temperature Sensitivity: A Potential Method for the Generation of Vaccines against the Avian Coronavirus Infectious Bronchitis Virus. Viruses 2020, 12, 754. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070754
Keep S, Stevenson-Leggett P, Steyn A, Oade MS, Webb I, Stuart J, Vervelde L, Britton P, Maier HJ, Bickerton E. Temperature Sensitivity: A Potential Method for the Generation of Vaccines against the Avian Coronavirus Infectious Bronchitis Virus. Viruses. 2020; 12(7):754. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070754
Chicago/Turabian StyleKeep, Sarah, Phoebe Stevenson-Leggett, Angela Steyn, Michael S. Oade, Isobel Webb, Jamie Stuart, Lonneke Vervelde, Paul Britton, Helena J. Maier, and Erica Bickerton. 2020. "Temperature Sensitivity: A Potential Method for the Generation of Vaccines against the Avian Coronavirus Infectious Bronchitis Virus" Viruses 12, no. 7: 754. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070754