MAVS Genetic Variation Is Associated with Decreased HIV-1 Replication In Vitro and Reduced CD4+ T Cell Infection in HIV-1-Infected Individuals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Ethics Statement
2.3. Quantification of Proviral DNA
2.4. Multiplex Assay
2.5. T Cell Phenotyping
2.6. Intracellular Cytokine Production of HIV-1-Specific T Cells
2.7. Cell Isolation and Infection
2.8. Statistical Analysis
3. Results
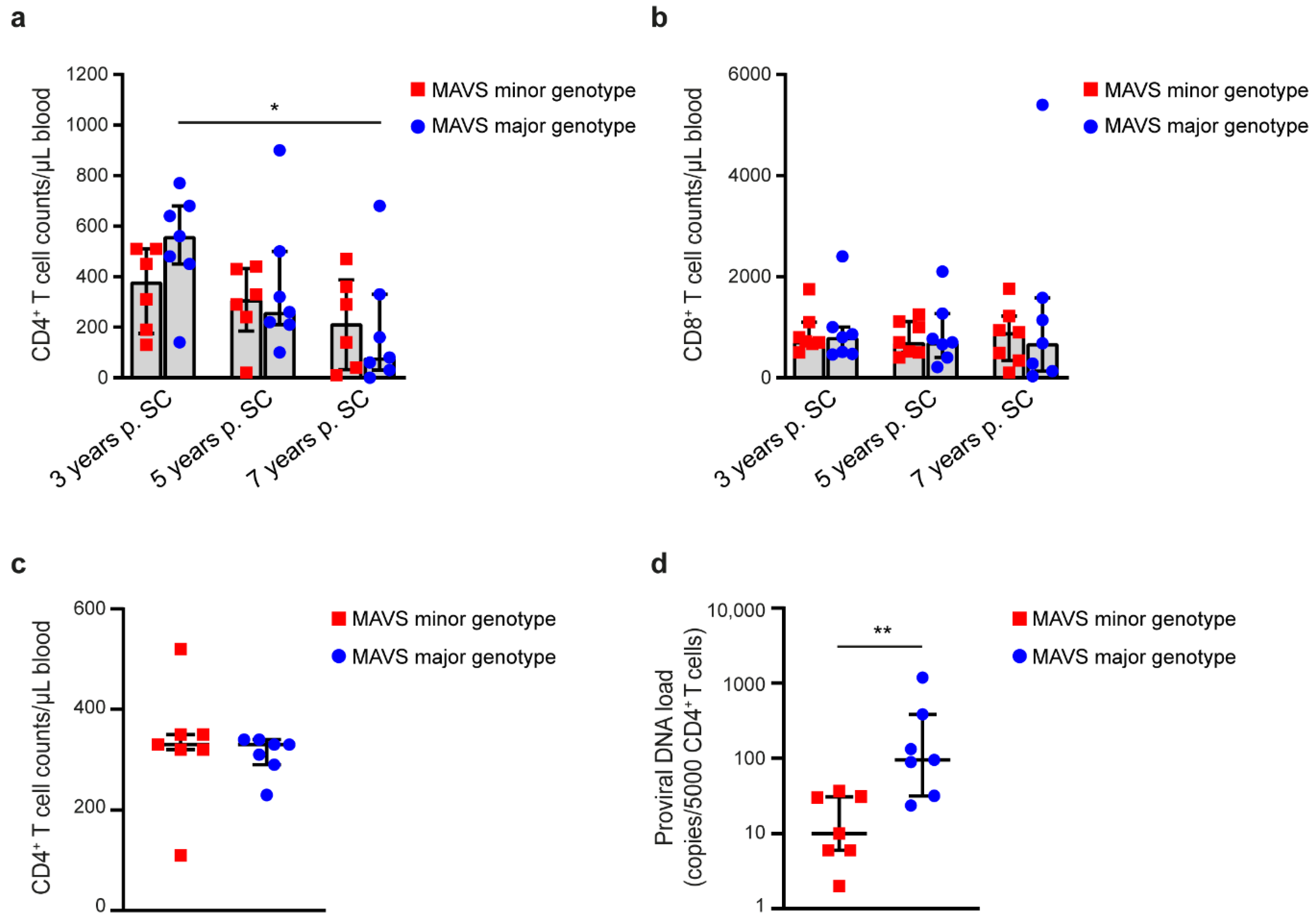
3.1. MAVS Minor Genotype Is Associated with Sustained CD4+ T Cell Counts and a Reduced Proviral DNA Load during HIV-1 Infection
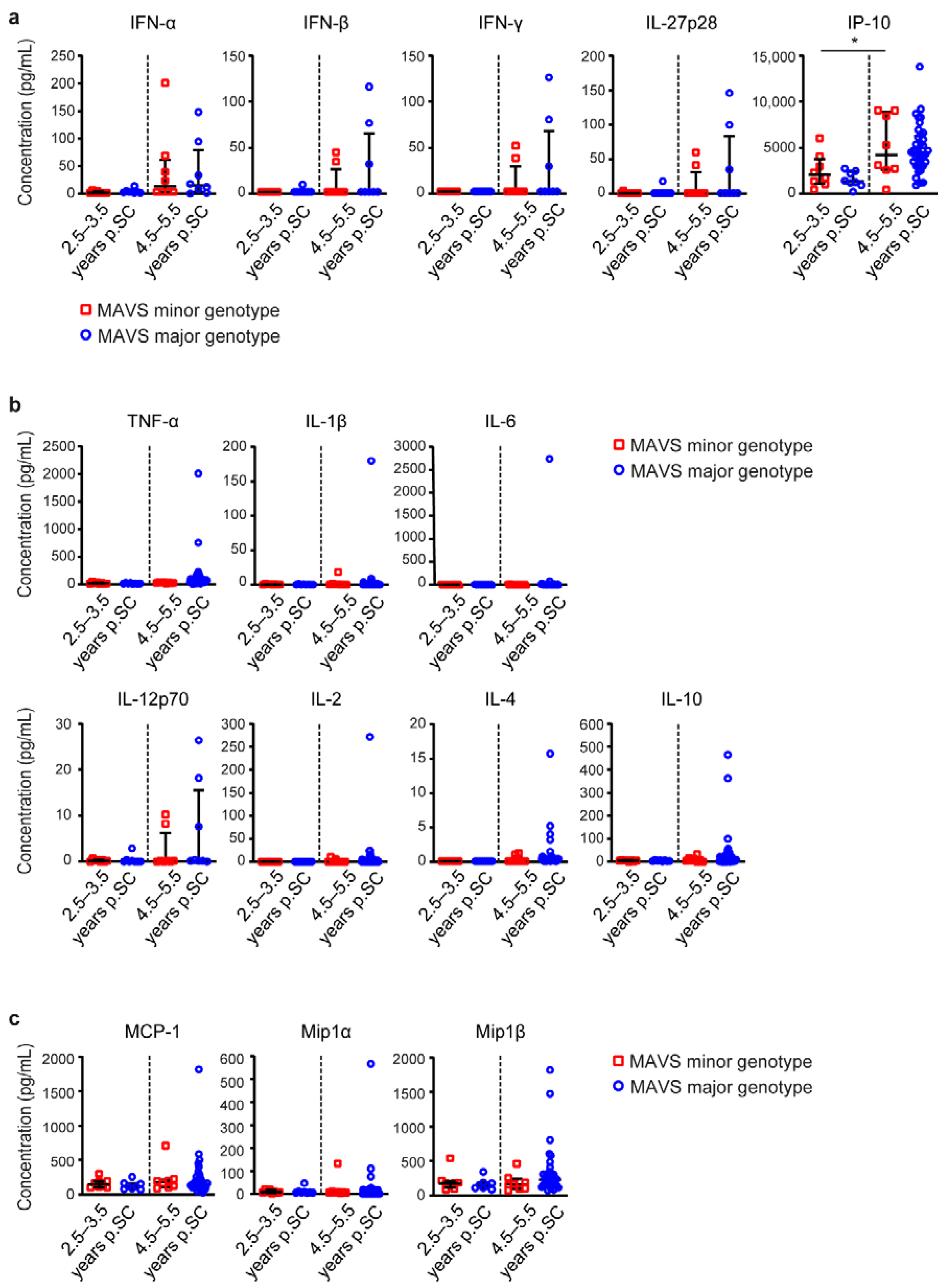
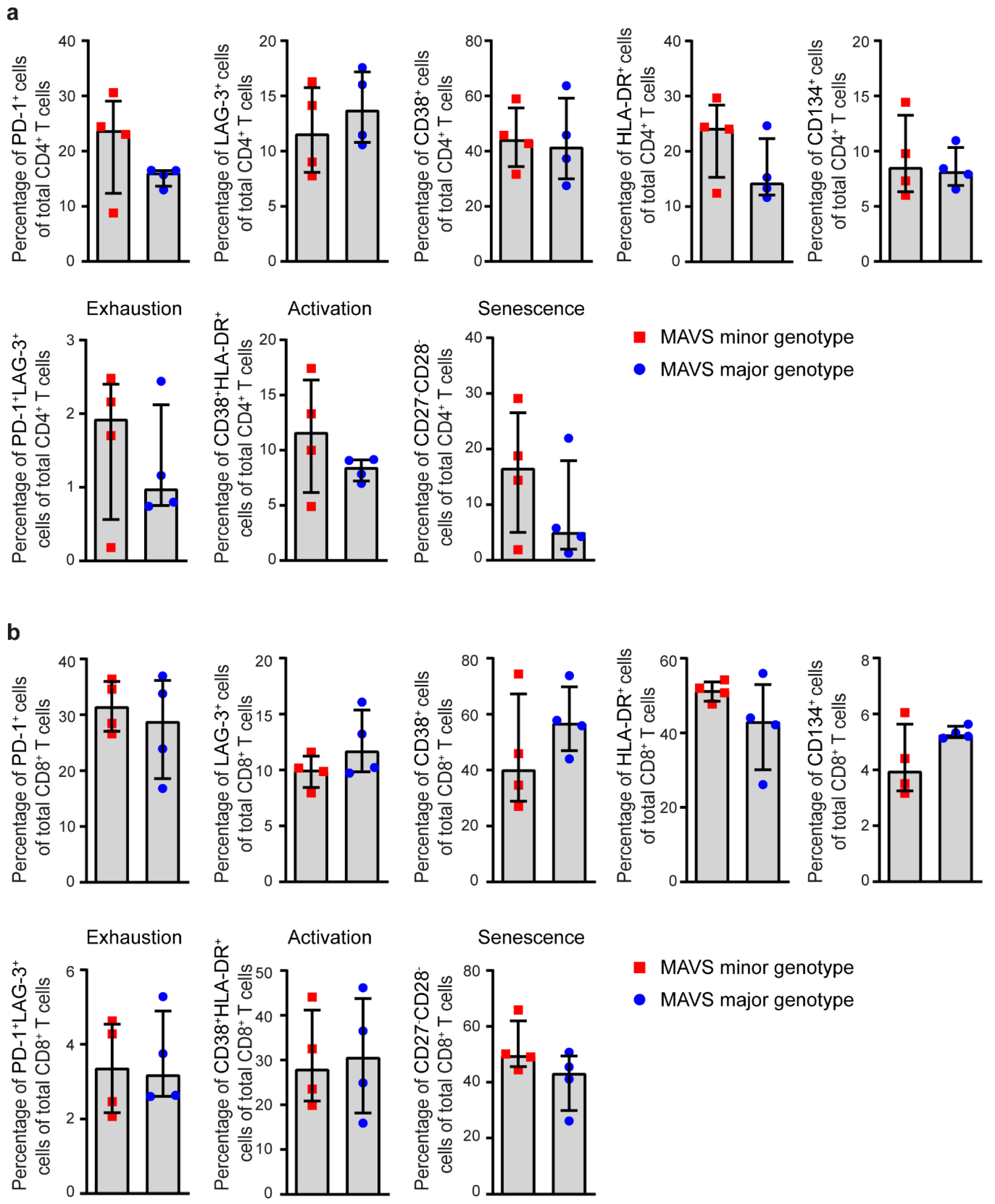
3.2. MAVS Genetic Variation Does Not Affect Immune Activation Levels
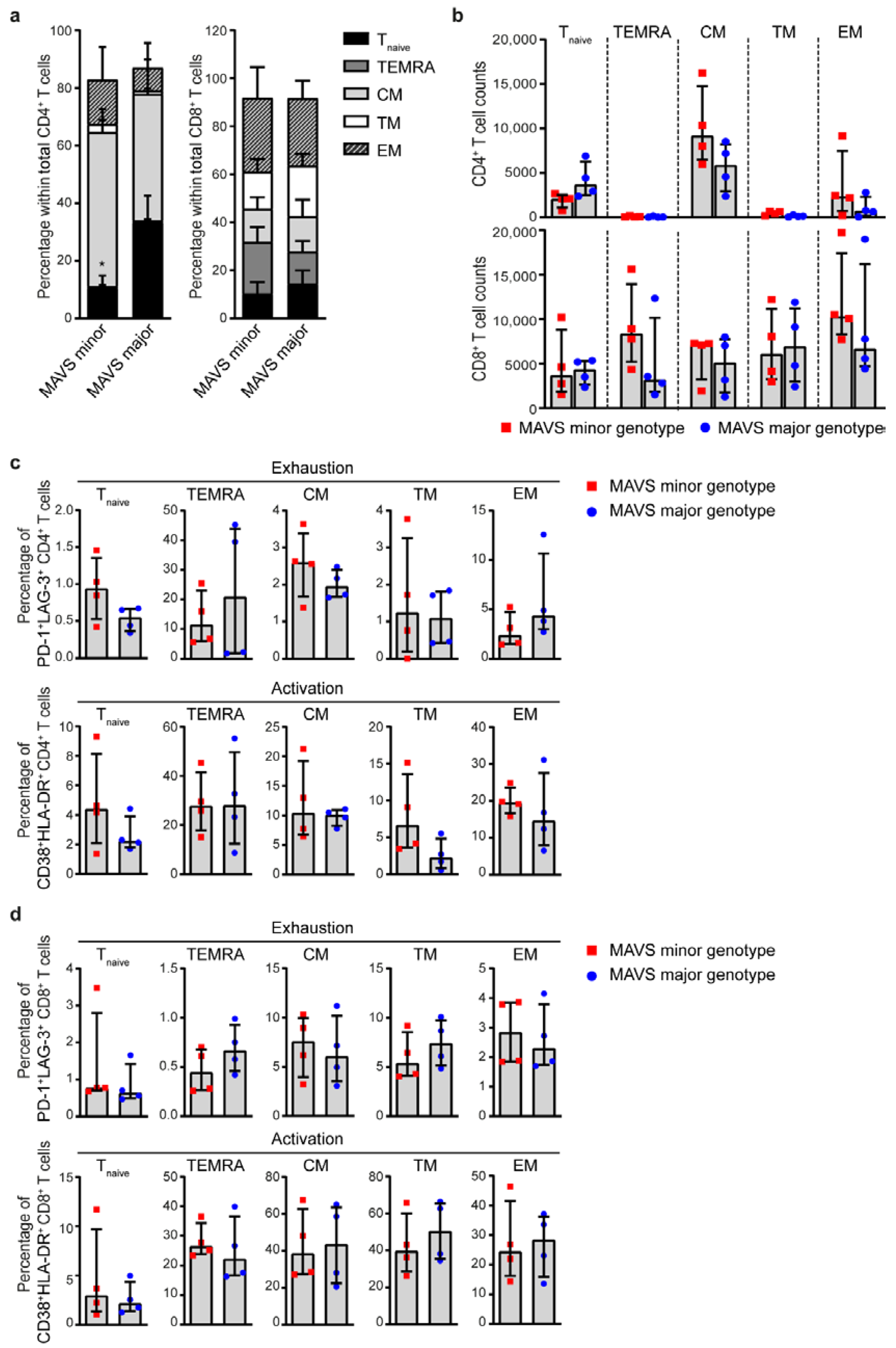
3.3. MAVS Minor Genotype Contains a Decreased Percentage of Naïve CD4+ T Cells
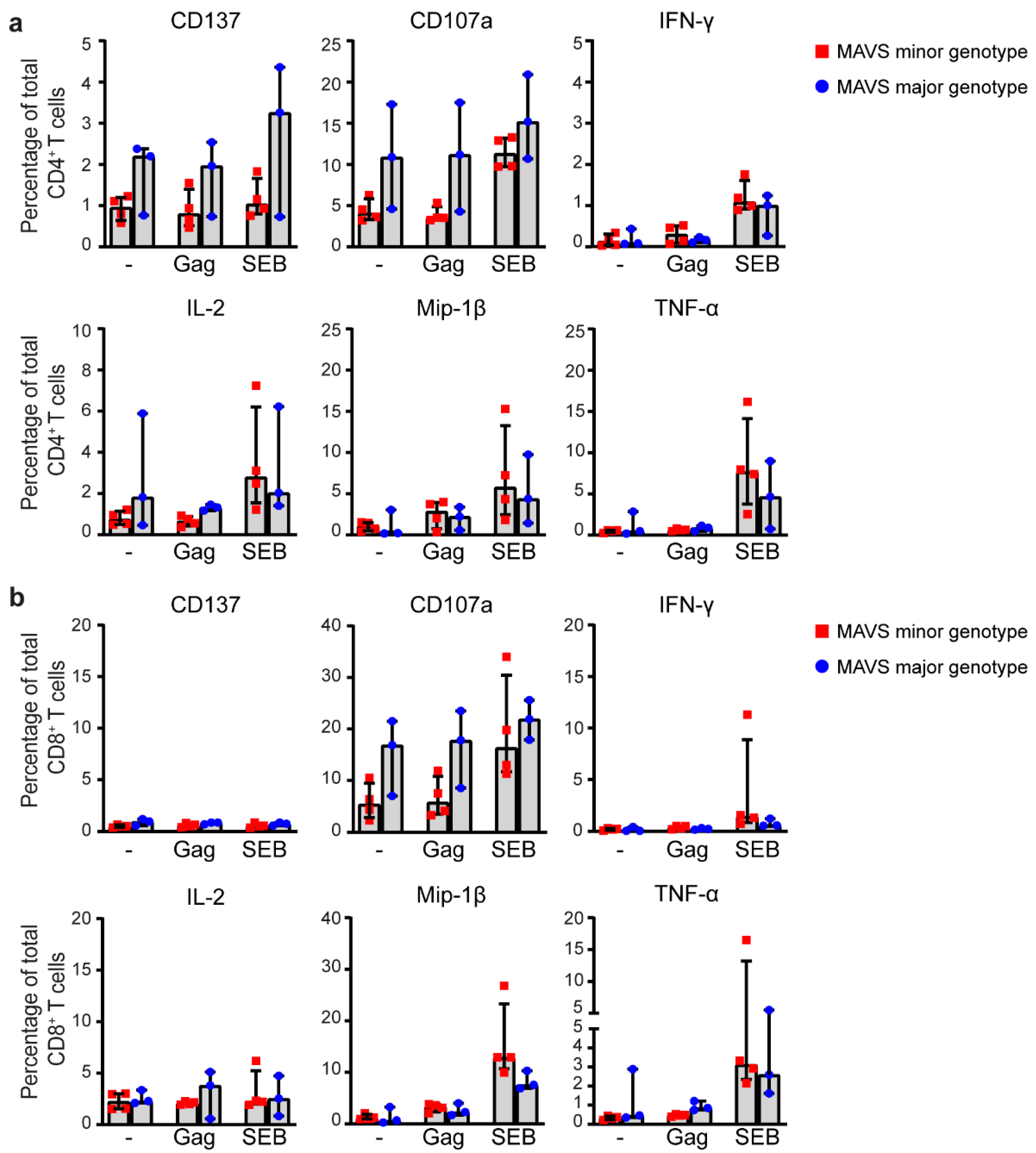
3.4. MAVS Genetic Variation Does Not Affect Intracellular Cytokine Responses of HIV-1-Specific T Cells
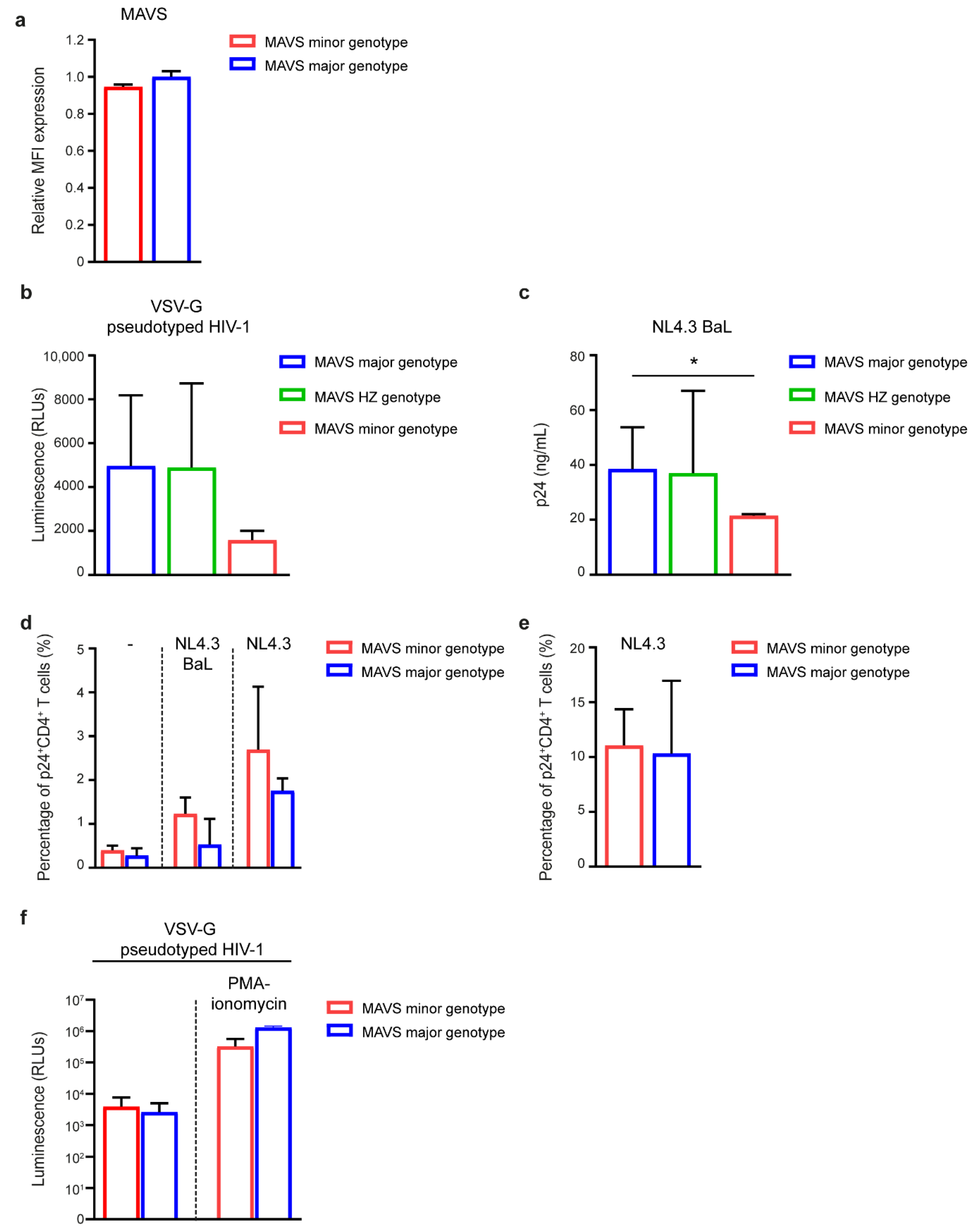
3.5. MAVS Minor Genotype Affects HIV-1 Replication In Vitro
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Che, K.F.; Sabado, R.L.; Shankar, E.M.; Tjomsland, V.; Messmer, D.; Bhardwaj, N.; Lifson, J.D.; Larsson, M. HIV-1 impairs in vitro priming of naïve T cells and gives rise to contact-dependent suppressor T cells. Eur. J. Immunol. 2010, 40, 2248–2258. [Google Scholar] [CrossRef] [PubMed]
- Luban, J. Innate immune sensing of HIV-1 by dendritic cells. Cell Host Microbe 2012, 12, 408–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautmann, L.; Janbazian, L.; Chomont, N.; Said, E.A.; Gimmig, S.; Bessette, B.; Boulassel, M.-R.; Delwart, E.; Sepulveda, H.; Balderas, R.S.; et al. Upregulation of PD-1 expression on HIV-specific CD8+ T cells leads to reversible immune dysfunction. Nat. Med. 2006, 12, 1198–1202. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J.; Ha, S.-J.; Kaech, S.M.; Haining, W.N.; Sarkar, S.; Kalia, V.; Subramaniam, S.; Blattman, J.N.; Barber, D.L.; Ahmed, R. Molecular Signature of CD8+ T Cell Exhaustion during Chronic Viral Infection. Immunity 2007, 27, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smed-Sörensen, A.; Loré, K.; Walther-Jallow, L.; Andersson, J.; Spetz, A.L. HIV-1-infected dendritic cells up-regulate cell surface markers but fail to produce IL-12 p70 in response to CD40 ligand stimulation. Blood 2004, 104, 2810–2817. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.; Bhardwaj, N. Dendritic cell dysregulation during HIV-1 infection. Immunol. Rev. 2013, 254, 170–189. [Google Scholar] [CrossRef] [Green Version]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef]
- Gringhuis, S.I.; van der Vlist, M.; van den Berg, L.M.; den Dunnen, J.; Litjens, M.; Geijtenbeek, T.B.H. HIV-1 exploits innate signaling by TLR8 and DC-SIGN for productive infection of dendritic cells. Nat. Immunol. 2010, 11, 419–426. [Google Scholar] [CrossRef]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a dendritic cell-specific HIV-1-binding protein that enhances trans-infection of T cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, M.R.; Bak, R.O.; Andersen, A.; Berg, R.K.; Jensen, S.B.; Jin, T.; Laustsen, A.; Hansen, K.; Østergaard, L.; Fitzgerald, K.A.; et al. IFI16 senses DNA forms of the lentiviral replication cycle and controls HIV-1 replication. Proc. Natl. Acad. Sci. USA 2013, 110, 19651. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, X.; Satoh, T.; Gentili, M.; Cerboni, S.; Conrad, C.; Hurbain, I.; ElMarjou, A.; Lacabaratz, C.; Lelièvre, J.D.; Manel, N. The Capsids of HIV-1 and HIV-2 Determine Immune Detection of the Viral cDNA by the Innate Sensor cGAS in Dendritic Cells. Immunity 2013, 39, 1132–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, R.K.; Melchjorsen, J.; Rintahaka, J.; Diget, E.; Søby, S.; Horan, K.A.; Gorelick, R.J.; Matikainen, S.; Larsen, C.S.; Ostergaard, L.; et al. Genomic HIV RNA induces innate immune responses through RIG-I-dependent sensing of secondary-structured RNA. PLoS ONE 2012, 7, e29291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yedavalli, V.S.R.K.; Neuveut, C.; Chi, Y.H.; Kleiman, L.; Jeang, K.T. Requirement of DDX3 DEAD box RNA helicase for HIV-1 Rev-RRE export function. Cell 2004, 119, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Rifo, R.; Rubilar, P.S.; Limousin, T.; de Breyne, S.; Décimo, D.; Ohlmann, T. DEAD-box protein DDX3 associates with eIF4F to promote translation of selected mRNAs. EMBO J. 2012, 31, 3745–3756. [Google Scholar] [CrossRef] [Green Version]
- Gringhuis, S.I.; Hertoghs, N.; Kaptein, T.M.; Zijlstra-Willems, E.M.; Sarrami-Fooroshani, R.; Sprokholt, J.K.; van Teijlingen, N.H.; Kootstra, N.A.; Booiman, T.; van Dort, K.A.; et al. HIV-1 blocks the signaling adaptor MAVS to evade antiviral host defense after sensing of abortive HIV-1 RNA by the host helicase DDX3. Nat. Immunol. 2017, 18, 225–235. [Google Scholar] [CrossRef]
- Spörri, R.; e Sousa, C.R. Inflammatory mediators are insufficient for full dendritic cell activation and promote expansion of CD4+ T cell populations lacking helper function. Nat. Immunol. 2005, 6, 163–170. [Google Scholar] [CrossRef]
- e Sousa, C.R. Dendritic cells in a mature age. Nat. Rev. Immunol. 2006, 6, 476–483. [Google Scholar] [CrossRef]
- Gringhuis, S.I.; den Dunnen, J.; Litjens, M.; van het Hof, B.; van Kooyk, Y.; Geijtenbeek, T.B.H. C-Type Lectin DC-SIGN Modulates Toll-like Receptor Signaling via Raf-1 Kinase-Dependent Acetylation of Transcription Factor NF-κB. Immunity 2007, 26, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Hou, B.; Reizis, B.; DeFranco, A.L. Toll-like Receptors Activate Innate and Adaptive Immunity by using Dendritic Cell-Intrinsic and-Extrinsic Mechanisms. Immunity 2008, 29, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Gringhuis, S.I.; den Dunnen, J.; Litjens, M.; van der Vlist, M.; Geijtenbeek, T.B.H. Carbohydrate-specific signaling through the DC-SIGN signalosome tailors immunity to Mycobacterium tuberculosis, HIV-1 and Helicobacter pylori. Nat. Immunol. 2009, 10, 1081–1088. [Google Scholar] [CrossRef]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.-M.; Gale, M. Immune Signaling by RIG-I-like Receptors. Immunity 2011, 34, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Takahashi, K.; Sato, S.; Coban, C.; Kumar, H.; Kato, H.; Ishii, K.J.; Takeuchi, O.; Akira, S. IPS-1, an adaptor triggering RIG-I-and Mda5-mediated type I interferon induction. Nat. Immunol. 2005, 6, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Seth, R.B.; Sun, L.; Ea, C.K.; Chen, Z.J. Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-κB and IRF3. Cell 2005, 122, 669–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitour, D.; Dabo, S.; Pour, M.A.; Vilasco, M.; Vidalain, P.O.; Jacob, Y.; Mezel-Lemoine, M.; Paz, S.; Arguello, M.; Lin, R.; et al. Polo-like kinase 1 (PLK1) regulates interferon (IFN) induction by MAVS. J. Biol. Chem. 2009, 284, 21797–21809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, S.; Vilasco, M.; Werden, S.J.; Arguello, M.; Joseph-Pillai, D.; Zhao, T.; Nguyen, T.L.A.; Sun, Q.; Meurs, E.F.; Lin, R.; et al. A functional C-terminal TRAF3-binding site in MAVS participates in positive and negative regulation of the IFN antiviral response. Cell Res. 2011, 21, 895–910. [Google Scholar] [CrossRef]
- Stunnenberg, M.; Geijtenbeek, T.B.H.; Gringhuis, S.I. DDX3 in HIV-1 infection and sensing: A paradox. Cytokine Growth Factor Rev. 2018, 40, 32–39. [Google Scholar] [CrossRef]
- Brenchley, J.M.; Douek, D.C. The mucosal barrier and immune activation in HIV pathogenesis. Curr. Opin. HIV AIDS 2008, 3, 356–361. [Google Scholar] [CrossRef]
- Deeks, S.G. HIV Infection, Inflammation, Immunosenescence, and Aging. Annu. Rev. Med. 2011, 62, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Hazenberg, M.D.; Otto, S.A.; Benthem, B.H.B.; van Roos, M.T.L.; Coutinho, R.A.; Lange, J.M.A.; Hamann, D.; Prins, M.; Miedema, F. Persistent immune activation in HIV-1 infection is associated with progression to AIDS (2003). Aids 2003, 17, 1881–1888. [Google Scholar] [CrossRef]
- Leng, Q.; Borkow, G.; Weisman, Z.; Stein, M.; Kalinkovich, A.; Bentwich, Z. Immune activation correlates better than HIV plasma viral load with CD4 T-cell decline during HIV infection. J. Acquir. Immune Defic. Syndr. 2001, 27, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Paiardini, M.; Muller-Trutwin, M. HIV-associated chronic immune activation. Immunol. Rev. 2013, 254, 78–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; McNevin, J.; Malhotra, U.; McElrath, M.J. Evolution of CD8+ T Cell Immunity and Viral Escape Following Acute HIV-1 Infection. J. Immunol. 2003, 171, 3837–3846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geels, M.J.; Jansen, C.A.; Baan, E.; De Cuyper, I.M.; Van Schijndel, G.J.M.; Schuitemaker, H.; Goudsmit, J.; Pollakis, G.; Miedema, F.; Paxton, W.A.; et al. CTL escape and increased viremia irrespective of HIV-specific CD4 + T-helper responses in two HIV-infected individuals. Virology 2006, 345, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.R.; van der Burg, S.H.; Hovenkamp, E.; Holwerda, A.M.; Drijfhout, J.W.; Melief, C.J.; Miedema, F. Characterization of HLA-B57-restricted human immunodeficiency virus type 1 Gag- and RT-specific cytotoxic T lymphocyte responses. J. Gen. Virol. 1998, 79 Pt 9, 2191–2201. [Google Scholar] [CrossRef] [Green Version]
- Kostense, S.; Ogg, G.S.; Manting, E.H.; Gillespie, G.; Joling, J.; Vandenberghe, K.; Veenhof, E.Z.; Van Baarle, D.; Jurriaans, S.; Klein, M.R.; et al. High viral burden in the presence of major HIV-specific CD8+ T cell expansions: Evidence for impaired CTL effector function. Eur. J. Immunol. 2001, 31, 677–686. [Google Scholar] [CrossRef]
- Van Baalen, C.A.; Klein, M.R.; Geretti, A.M.; Keet, R.I.; Miedema, F.; van Els, C.; Osterhaus, A.D. Selective in vitro expansion of HLA class I-restricted HIV-1 Gag-specific CD8+ T cells: Cytotoxic T-lymphocyte epitopes and precursor frequencies. Aids 1993, 7, 781–786. [Google Scholar] [CrossRef]
- Addo, M.M.; Yu, X.G.; Rathod, A.; Cohen, D.; Eldridge, R.L.; Strick, D.; Johnston, M.N.; Corcoran, C.; Wurcel, A.G.; Fitzpatrick, C.A.; et al. Comprehensive Epitope Analysis of Human Immunodeficiency Virus Type 1 (HIV-1)-Specific T-Cell Responses Directed against the Entire Expressed HIV-1 Genome Demonstrate Broadly Directed Responses, but No Correlation to Viral Load. J. Virol. 2003, 77, 2081–2092. [Google Scholar] [CrossRef] [Green Version]
- Goonetilleke, N.; Liu, M.K.P.; Salazar-Gonzalez, J.F.; Ferrari, G.; Giorgi, E.; Ganusov, V.V.; Keele, B.F.; Learn, G.H.; Turnbull, E.L.; Salazar, M.G.; et al. The first T cell response to transmitted/founder virus contributes to the control of acute viremia in HIV-1 infection. J. Exp. Med. 2009, 206, 1253–1272. [Google Scholar] [CrossRef]
- Borrow, P.; Lewicki, H.; Wei, X.; Horwitz, M.S.; Peffer, N.; Meyers, H.; Nelson, J.A.; Gairin, J.E.; Hahn, B.H.; Oldstone, M.B.; et al. Antiviral pressure exerted by HIV-l-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med. 1997, 3, 205–211. [Google Scholar] [CrossRef]
- Alter, G.; Merchant, A.; Tsoukas, C.M.; Rouleau, D.; LeBlanc, R.P.; Cote, P.; Baril, J.-G.; Thomas, R.; Nguyen, V.-K.; Sekaly, R.-P.; et al. Human Immunodeficiency Virus (HIV)-Specific Effector CD8 T Cell Activity in Patients with Primary HIV Infection. J. Infect. Dis. 2002, 185, 755–765. [Google Scholar] [CrossRef] [Green Version]
- Lichterfeld, M.; Kaufmann, D.E.; Yu, X.G.; Mui, S.K.; Addo, M.M.; Johnston, M.N.; Cohen, D.; Robbins, G.K.; Pae, E.; Alter, G.; et al. Loss of HIV-1-specific CD8+ T cell proliferation after acute HIV-1 infection and restoration by vaccine-induced HIV-1-specific CD4+ T cells. J. Exp. Med. 2004, 200, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Qiu, C.; Qiu, C.; Wang, Y.; Zhang, X.; Xu, J. Development of Skewed Functionality of HIV-1-Specific Cytotoxic CD8+ T Cells from Primary to Early Chronic Phase of HIV Infection. PLoS ONE 2012, 7, e44983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Griensven, G.J.P.; De Vroome, E.M.M.; Goudsmit, J.; Coutinho, R.A. Changes in sexual behaviour and the fall in incidence of HIV infection among homosexual men. Br. Med. J. 1989, 298, 218–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folks, T.M.; Powell, D.; Lightfoote, M.; Koenig, S.; Fauci, A.S.; Benn, S.; Rabson, A.; Daugherty, D.; Gendelman, H.E.; David Hoggan, M.; et al. Biological and biochemical characterization of a cloned leu-3−cell surviving infection with the acquired immune deficiency syndrome retrovirus. J. Exp. Med. 1986, 164, 280–290. [Google Scholar] [CrossRef] [Green Version]
- Mesman, A.W.; Zijlstra-Willems, E.M.; Kaptein, T.M.; De Swart, R.L.; Davis, M.E.; Ludlow, M.; Duprex, W.P.; Gack, M.U.; Gringhuis, S.I.; Geijtenbeek, T.B.H. Measles virus suppresses RIG-I-like receptor activation in dendritic cells via DC-SIGN-mediated inhibition of PP1 phosphatases. Cell Host Microbe 2014, 16, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Connor, R.I.; Chen, B.K.; Choe, S.; Landau, N.R. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology 1995, 206, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Watts, T.H. Tnf/Tnfr Family Members in Costimulation of T Cell Responses. Annu. Rev. Immunol. 2005, 23, 23–68. [Google Scholar] [CrossRef]
- Betts, M.R.; Koup, R.A. Detection of T-cell degranulation: CD107a and b. Methods Cell Biol. 2004, 2004, 497–512. [Google Scholar] [CrossRef]
- Liu, M.; Guo, S.; Stiles, J.K. The emerging role of CXCL10 in cancer. Oncol. Lett. 2011, 2, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Zhang, T.; Wang, R.; Zhang, H.; Huang, X.; Yin, J.; Zhang, L.; Xu, X.; Wu, H. Plasma IP-10 is associated with rapid disease progression in early HIV-1 infection. Viral Immunol. 2012, 25, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Liovat, A.S.; Rey-Cuillé, M.A.; Lécuroux, C.; Jacquelin, B.; Girault, I.; Petitjean, G.; Zitoun, Y.; Venet, A.; Barré-Sinoussi, F.; Lebon, P.; et al. Acute Plasma Biomarkers of T Cell Activation Set-Point Levels and of Disease Progression in HIV-1 Infection. PLoS ONE 2012, 7, e46143. [Google Scholar] [CrossRef] [PubMed]
- Ploquin, M.J.; Madec, Y.; Casrouge, A.; Huot, N.; Passaes, C.; Lécuroux, C.; Essat, A.; Boufassa, F.; Jacquelin, B.; Jochems, S.P.; et al. Elevated Basal Pre-infection CXCL10 in Plasma and in the Small Intestine after Infection Are Associated with More Rapid HIV/SIV Disease Onset. PLoS Pathog. 2016, 12, e1005774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, R.P.; Scully, E.P.; Groden, E.E.; Arnold, K.B.; Chang, J.J.; Lane, K.; Lifson, J.; Rosenberg, E.; Lauffenburger, D.A.; Altfeld, M. HIV-1 infection induces strong production of IP-10 through TLR7/9-dependent pathways. Aids 2013, 27, 2505–2517. [Google Scholar] [CrossRef] [Green Version]
- Durudas, A.; Chen, H.-L.; A Gasper, M.; Sundaravaradan, V.; M Milush, J.; Silvestri, G.; Johnson, W.; D Giavedoni, L.; L Sodora, D. Differential Innate Immune Responses to Low or High Dose Oral SIV Challenge in Rhesus Macaques. Curr. HIV Res. 2011, 9, 276–288. [Google Scholar] [CrossRef] [Green Version]
- Blaak, H.; Van’t Wout, A.B.; Brouwer, M.; Hooibrink, B.; Hovenkamp, E.; Schuitemaker, H. In vivo HIV-1 infection of CD45RA+CD4+ T cells is established primarily by syncytium-inducing variants and correlates with the rate of CD4+ T cell decline. Proc. Natl. Acad. Sci. USA 2000, 97, 1269–1274. [Google Scholar] [CrossRef] [Green Version]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef]
- Doitsh, G.; Cavrois, M.; Lassen, K.G.; Zepeda, O.; Yang, Z.; Santiago, M.L.; Hebbeler, A.M.; Greene, W.C. Abortive HIV Infection Mediates CD4 T-Cell Depletion and Inflammation in Human Lymphoid Tissue. Cell 2010, 143, 789–801. [Google Scholar] [CrossRef] [Green Version]
- Doitsh, G.; Galloway, N.L.K.K.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-arias, I.; et al. Pyroptosis drives CD4 T-cell depletion. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Koup, R.A.; Safrit, J.T.; Cao, Y.; Andrews, C.A.; McLeod, G.; Borkowsky, W.; Farthing, C.; Ho, D.D. Temporal association of cellular immune responses with the initial control of viremia in primary human immunodeficiency virus type 1 syndrome. J. Virol. 1994, 68, 4650–4655. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Bauer, D.E.; Tuttleton, S.E.; Lewin, S.; Gettie, A.; Blanchard, J.; Irwin, C.E.; Safrit, J.T.; Mittler, J.; Weinberger, L.; et al. Dramatic rise in plasma viremia after CD8+ T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med. 1999, 189, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, M.R.; van Baalen, C.A.; Holwerda, A.M.; Garde, S.R.K.; Bende, R.J.; Keet, I.P.M.; Eeftinck-Schattenkerk, J.K.M.; Osterhaus, A.D.M.E.; Schuitemaker, H.; Miedema, F. Kinetics of gag-specific cytotoxic T lymphocyte responses during the clinical course of HIV-1 infection: A longitudinal analysis of rapid progressors and long-term asymptomatics. J. Exp. Med. 1995, 181, 1365–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, S.J.; Woodward, A.; Zhao, A. Human Immunodeficiency Virus Type 1 (HIV-1)-Specific T Cell Responses Correlate with Control of Acute HIV-1 Infection in Macaques. J. Infect. Dis. 1997, 176, 1188–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.E.; Rowland-Jones, S.; Nixon, D.F.; Gotch, F.M.; Edwards, J.P.; Ogunlesi, A.O.; Elvin, J.G.; Rothbard, J.A.; Bangham, C.R.M.; Rizza, C.R.; et al. Human immunodeficiency virus genetic variation that can escape cytotoxic T cell recognition. Nature 1991, 354, 453–459. [Google Scholar] [CrossRef]
- Jones, N.A.; Wei, X.; Flower, D.R.; Wong, M.L.; Michor, F.; Saag, M.S.; Hahn, B.H.; Nowak, M.A.; Shaw, G.M.; Borrow, P. Determinants of human immunodeficiency virus type 1 escape from the primary CD8+ cytotoxic T lymphocyte response. J. Exp. Med. 2004, 200, 1243–1256. [Google Scholar] [CrossRef]
- Leslie, A.J.; Pfafferott, K.J.; Chetty, P.; Draenert, R.; Addo, M.M.; Feeney, M.; Tang, Y.; Holmes, E.C.; Allen, T.; Prado, J.G.; et al. HIV evolution: CTL escape mutation and reversion after transmission. Nat. Med. 2004, 10, 282–289. [Google Scholar] [CrossRef]
- Martinez-Picado, J.; Prado, J.G.; Fry, E.E.; Pfafferott, K.; Leslie, A.; Chetty, S.; Thobakgale, C.; Honeyborne, I.; Crawford, H.; Matthews, P.; et al. Fitness Cost of Escape Mutations in p24 Gag in Association with Control of Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 3617–3623. [Google Scholar] [CrossRef] [Green Version]
- Peyerl, F.W.; Bazick, H.S.; Newberg, M.H.; Barouch, D.H.; Sodroski, J.; Letvin, N.L. Fitness Costs Limit Viral Escape from Cytotoxic T Lymphocytes at a Structurally Constrained Epitope. J. Virol. 2004, 78, 13901–13910. [Google Scholar] [CrossRef] [Green Version]
- Navis, M.; Schellens, I.; van Baarle, D.; Borghans, J.; van Swieten, P.; Miedema, F.; Kootstra, N.; Schuitemaker, H. Viral Replication Capacity as a Correlate of HLA B57/B5801-Associated Nonprogressive HIV-1 Infection. J. Immunol. 2007, 179, 3133–3143. [Google Scholar] [CrossRef] [Green Version]
- Schellens, I.M.M.; Spits, H.B.; Navis, M.; Westerlaken, G.H.A.; Nanlohy, N.M.; Coffeng, L.E.; Kootstra, N.; Miedema, F.; Schuitemaker, H.; Borghans, J.A.M.; et al. Differential characteristics of cytotoxic T lymphocytes restricted by the protective HLA alleles B∗27 and B∗57 in HIV-1 infection. J. Acquir. Immune Defic. Syndr. 2014, 67, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Kiepiela, P.; Ngumbela, K.; Thobakgale, C.; Ramduth, D.; Honeyborne, I.; Moodley, E.; Reddy, S.; De Pierres, C.; Mncube, Z.; Mkhwanazi, N.; et al. CD8+ T-cell responses to different HIV proteins have discordant associations with viral load. Nat. Med. 2007, 13, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Borghans, J.A.M.; Mølgaard, A.; de Boer, R.J.; Keşmir, C. HLA alleles associated with slow progression to AIDS truly prefer to present HIV-1 p24. PLoS ONE 2007, 2, e920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streeck, H.; Lichterfeld, M.; Alter, G.; Meier, A.; Teigen, N.; Yassine-Diab, B.; Sidhu, H.K.; Little, S.; Kelleher, A.; Routy, J.-P.; et al. Recognition of a Defined Region within p24 Gag by CD8+ T Cells during Primary Human Immunodeficiency Virus Type 1 Infection in Individuals Expressing Protective HLA Class I Alleles. J. Virol. 2007, 81, 7725–7731. [Google Scholar] [CrossRef] [Green Version]
- Zuñiga, R.; Lucchetti, A.; Galvan, P.; Sanchez, S.; Sanchez, C.; Hernandez, A.; Sanchez, H.; Frahm, N.; Linde, C.H.; Hewitt, H.S.; et al. Relative Dominance of Gag p24-Specific Cytotoxic T Lymphocytes Is Associated with Human Immunodeficiency Virus Control. J. Virol. 2006, 80, 3122–3125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.Q.; Lu, Y.; Thulasi Raman, S.N.; Xu, F.; Wu, Q.; Li, Z.; Brownlie, R.; Liu, Q.; Zhou, Y. Nuclear-resident RIG-I senses viral replication inducing antiviral immunity. Nat. Commun. 2018, 9, 3199. [Google Scholar] [CrossRef]
- Heerma Van Voss, M.R.; Vesuna, F.; Bol, G.M.; Meeldijk, J.; Raman, A.; Offerhaus, G.J.; Buerger, H.; Patel, A.H.; Van Der Wall, E.; Van Diest, P.J.; et al. Nuclear DDX3 expression predicts poor outcome in colorectal and breast cancer. Onco. Targets Ther. 2017, 10, 3501–3513. [Google Scholar] [CrossRef] [Green Version]
- Brennan, R.; Haap-Hoff, A.; Gu, L.; Gautier, V.; Long, A.; Schröder, M. Investigating nucleo-cytoplasmic shuttling of the human DEAD-box helicase DDX3. Eur. J. Cell Biol. 2018, 97, 501–511. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stunnenberg, M.; van Pul, L.; Sprokholt, J.K.; van Dort, K.A.; Gringhuis, S.I.; Geijtenbeek, T.B.H.; Kootstra, N.A. MAVS Genetic Variation Is Associated with Decreased HIV-1 Replication In Vitro and Reduced CD4+ T Cell Infection in HIV-1-Infected Individuals. Viruses 2020, 12, 764. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070764
Stunnenberg M, van Pul L, Sprokholt JK, van Dort KA, Gringhuis SI, Geijtenbeek TBH, Kootstra NA. MAVS Genetic Variation Is Associated with Decreased HIV-1 Replication In Vitro and Reduced CD4+ T Cell Infection in HIV-1-Infected Individuals. Viruses. 2020; 12(7):764. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070764
Chicago/Turabian StyleStunnenberg, Melissa, Lisa van Pul, Joris K. Sprokholt, Karel A. van Dort, Sonja I. Gringhuis, Teunis B. H. Geijtenbeek, and Neeltje A. Kootstra. 2020. "MAVS Genetic Variation Is Associated with Decreased HIV-1 Replication In Vitro and Reduced CD4+ T Cell Infection in HIV-1-Infected Individuals" Viruses 12, no. 7: 764. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070764