Genetic Characterization of a Novel North American-Origin Avian Influenza A (H6N5) Virus Isolated from Bean Goose of South Korea in 2018

Abstract
:1. Introduction
2. Materials and Methods
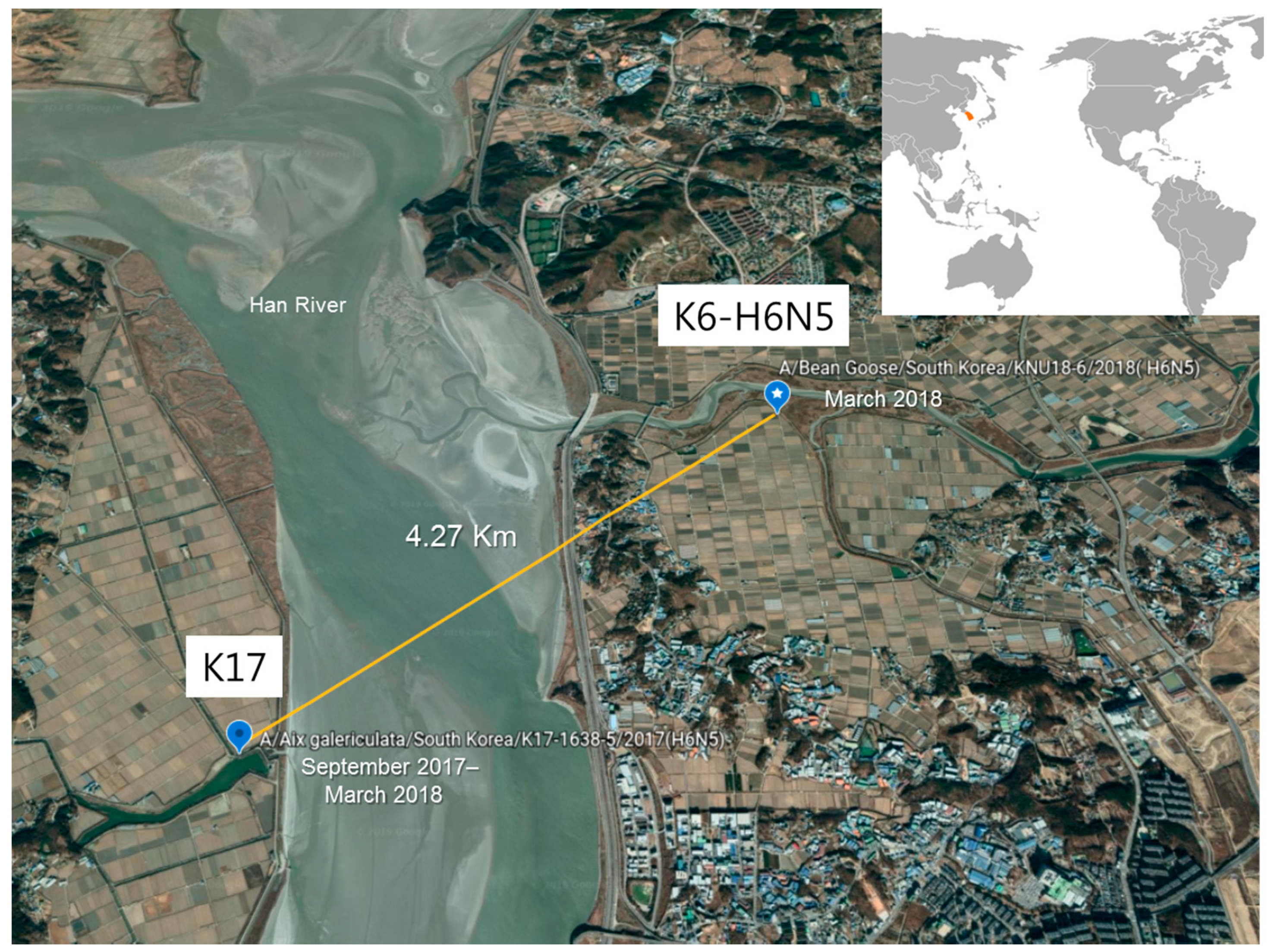
2.1. Sample Collection
2.2. Virus Isolation from the Feces
2.3. RNA Extraction and Bird Identification
2.4. Next-Generation Sequencing (NGS) by Illumina HiSeq X Method
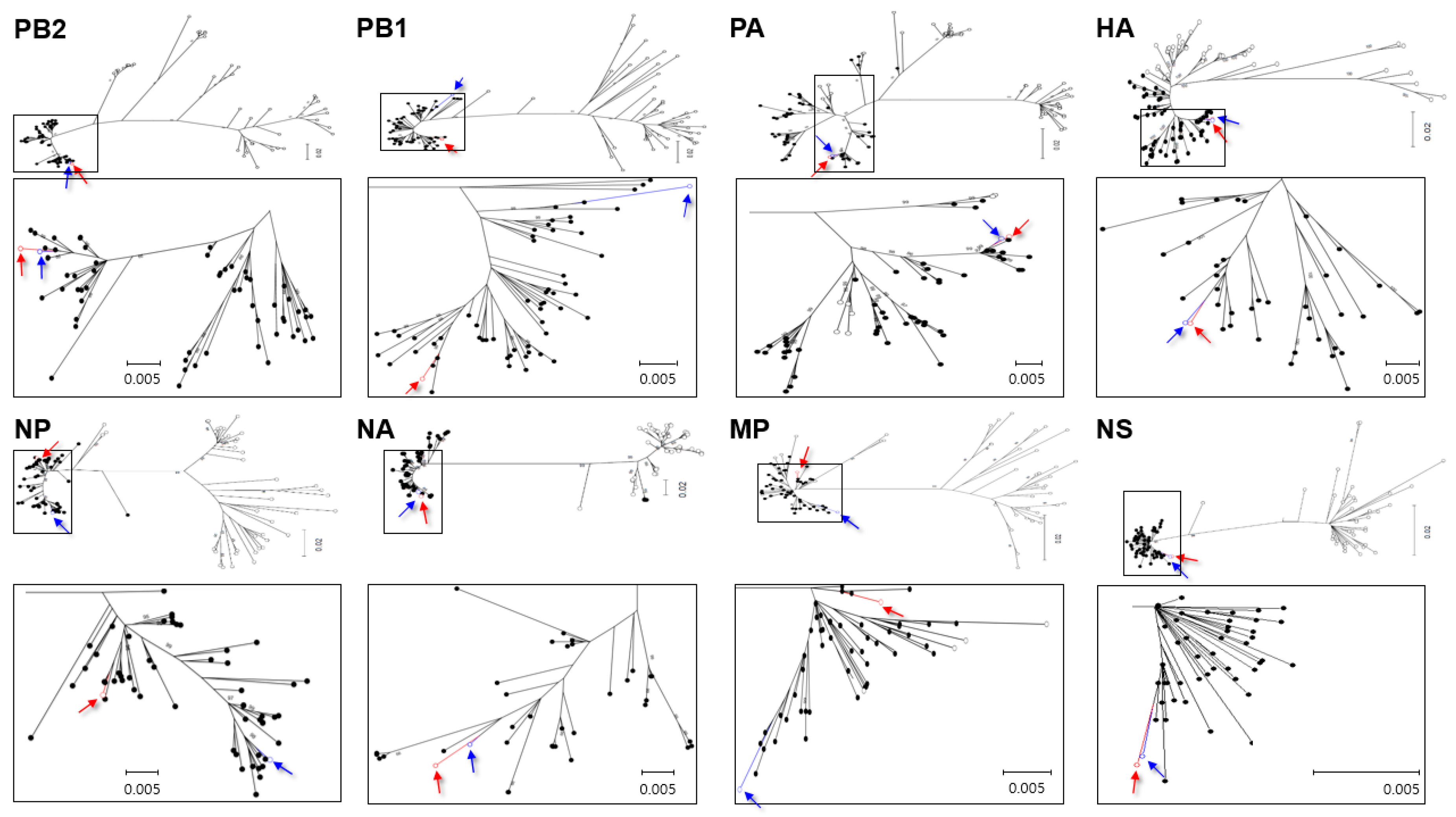
2.5. Phylogenetic Analysis
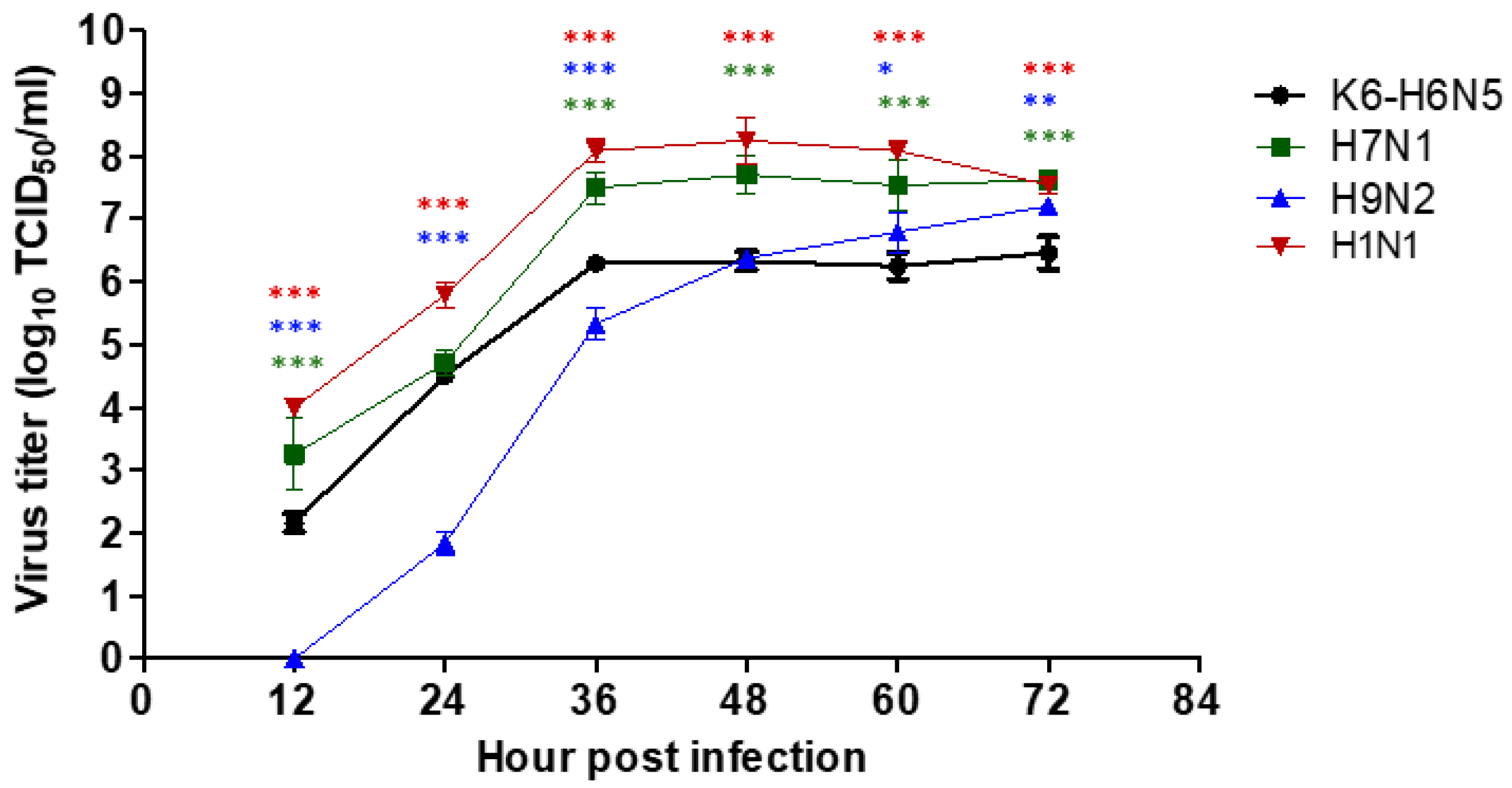
2.6. Viral Replication Kinetics in MDCK Cells
2.7. TCID50—50% Tissue Culture Infectious Dose and EID50—50% Egg Infectious Dose
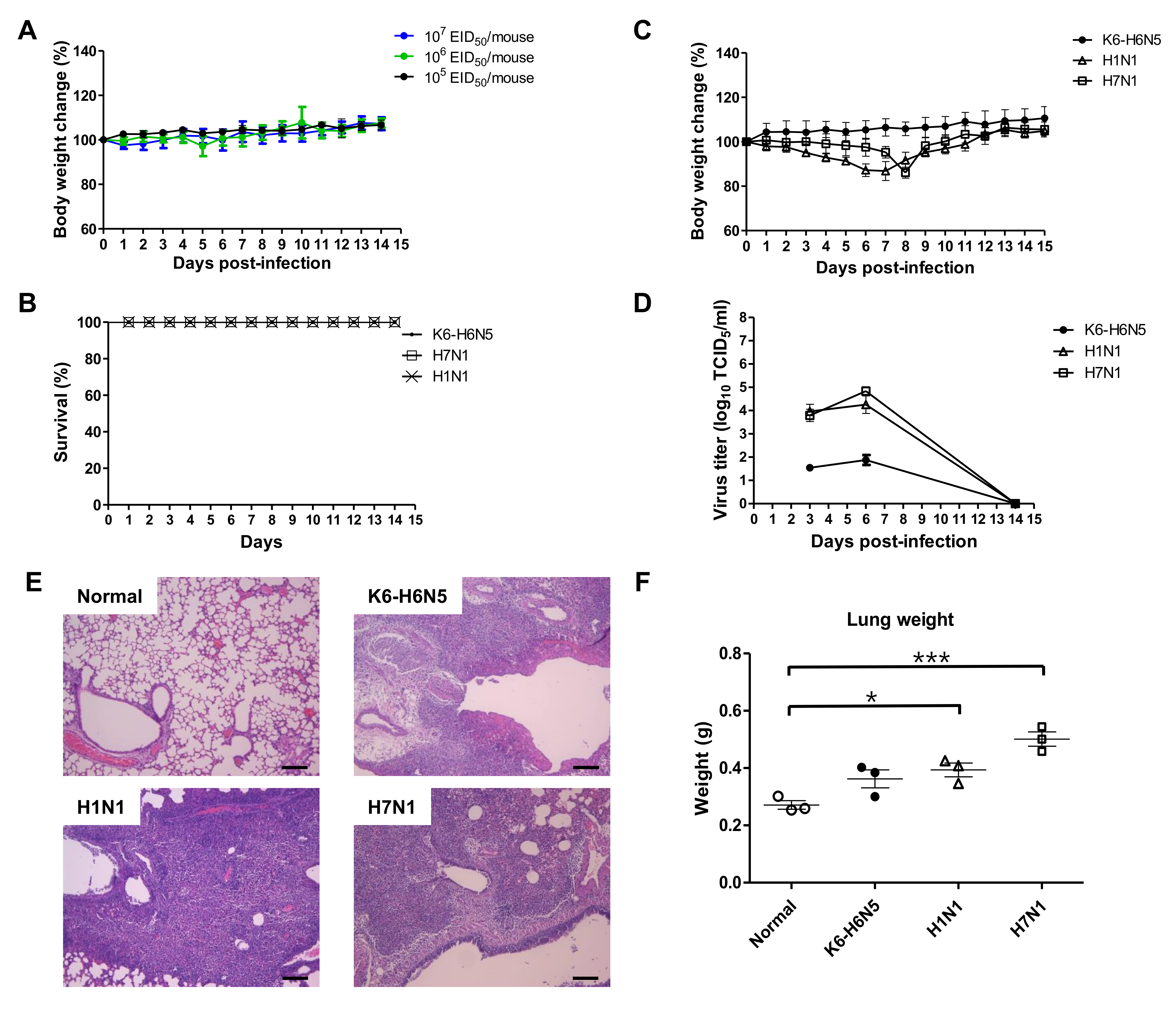
2.8. Animal Study
2.9. Statistics
3. Results
3.1. Genetic Characterization of Novel Avian Influenza A (H6N5) in 2018
3.2. Molecular Characterization of H6N5 Isolate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viral Protein | Amino Acid | K6-2018 | K17-2017 | CN5-2009 | W69-2005 | TW02/2013 | Comments | Reference |
|---|---|---|---|---|---|---|---|---|
| HA * | Cleavage sites | PQIETR↓GLF | PQIETR↓GLF | PQIETR↓GLF | PQIETR↓GLF | PQIATR↓GLF | Monobasic-LPAI | [24] |
| HA *-Receptor-binding site | A138S | A | A | A | A | A | Switch the preference from avian- to human-type receptors | [25] |
| E190D | E | E | E | E | V | Switch the preference from avian- to human-type receptors | [25] | |
| G225D | G | G | G | G | G | Switch the preference from avian- to human-type receptors | [25] | |
| Q226L | Q | Q | Q | Q | Q | Switch the preference from avian- to human-type receptors | [25] | |
| G228S | G | G | G | G | S | Switch the preference from avian- to human-type receptors | [25] | |
| HA* | L102F | L | L | L | L | L | Increased pathogenicity in mice | [28] |
| T/E173G/D/V | T | T | N | N | T | Increased virus binding to α-2,6-linked sialic acid | [28] | |
| N379T | N | N | N | N | N | Increased virulence in mammals | [29] | |
| NA | Amino acid deletion at stalk region | No | No | No | No | Yes | Increased virulence in mice | [26] |
| E119V | V | V | V | V | I | Reduced inhibition by OS (Oseltamivir) and PER (Peramivir) and highly reduced inhibition by ZA (Zanamivir) | [21] | |
| PB2 | T63I (with PB1 M677T) | I | I | I | I | I | Pathogenic in mice | [30] |
| L89V | V | V | V | V | V | Enhanced polymerase activity, increased virulence in mice | [31] | |
| K251R | R | R | R | R | K | Increased virulence in mice | [32] | |
| A274T | T | A | A | A | A | Increased polymerase activity, increased virulence in mammals and birds | [33] | |
| G309D | D | D | D | D | D | Enhanced polymerase activity, increased virulence in mice | [31] | |
| T339K | K | K | K | K | K | Enhanced polymerase activity, increased virulence in mice | [31] | |
| Q368R | R | R | R | R | R | Increased polymerase activity, increased virulence in mammals | [34,35] | |
| H447Q | Q | Q | Q | Q | Q | Increased polymerase activity, increased virulence in mammals | [34,35] | |
| R477G | G | G | G | G | G | Enhanced polymerase activity, increased virulence in mice | [31] | |
| I495V | V | V | V | V | V | Enhanced polymerase activity, increased virulence in mice | [31] | |
| A676T | T | T | T | T | V | Enhanced polymerase active, increased virulence in mice | [31] | |
| D701N | D | D | N | D | D | Increased polymerase activity, increased virulence in mammals, mammalian host marker | [36,37,38] | |
| PB1 | D/A3V | V | V | V | V | V | Increased polymerase activity, increased virulence in mammals | [34,35] |
| L13P | P | P | P | P | P | Increased polymerase activity, increased virulence in mammals, mammalian host marker | [36,37] | |
| R207K | K | K | K | K | K | Increased polymerase activity in mammalian cells | [39] | |
| K328N | N | N | N | N | S | Increased polymerase activity, increased virulence in mammals | [34,35] | |
| S375N/T | N | S | N | N | N | Increased polymerase activity, increased virulence in mammals, human host marker | [34,35,40] | |
| H436Y | Y | Y | Y | Y | Y | Increased polymerase activity and virulence in mallards, ferrets, and mice | [39] | |
| A469T (with NS1 N205K; NEP T48N) | T | T | T | T | T | Conferred in contact transmissibility in guinea pigs | [41] | |
| L473V | V | V | V | V | V | Increased polymerase activity and replication efficiency | [42] | |
| V652A | A | A | A | A | A | Increased virulence in mice | [32] | |
| M677T (with PB2 T63I) | T | T | T | T | T | Pathogenic in mice | [30] | |
| PB1-F2 | N66S | N | S | N | N | Truncated | Increased virulence in mammals | [43,44] |
| PA | S37A | A | A | A | A | A | Significantly increased viral growth and polymerase activity in mammalian cells | [45] |
| H266R | R | R | R | R | R | Increased polymerase activity, increased virulence in mammals and birds | [33] | |
| F277S | S | S | S | S | S | Adapt to mammalian hosts | [46] | |
| C278Q | Q | Q | Q | Q | Q | Adapt to mammalian hosts | [46] | |
| E382D | D | D | E | E | E | Human host marker | [40,47] | |
| S/A515T | T | T | T | T | T | Increased polymerase activity, increased virulence in mammals and birds | [33,39] | |
| L653P | S | P | P | P | P | Adapt to mammalian hosts | [46] | |
| NP | V41I | I | I | I | I | I | Might contribute to viral transmissibility | [48] |
| V105M | M | V | M | I | I | Contribute to the increased virulence | [49] | |
| F253I | I | I | I | I | I | Results in attenuated pathogenicity of the virus in mice | [50] | |
| I353V | V | V | V | V | V | Increased virulence in mice | [32] | |
| MP | N30D | D | D | D | D | D | Increased virulence in mammals | [51] |
| T215A | A | A | A | A | A | Increased virulence in mammals | [52] | |
| NS1 | A/P42S | S | S | S | S | S | Increased virulence in mammals, antagonism of IFN induction | [52,53] |
| T/D/V/R/A127N | N | N | N | N | N | Increased virulence in mammals | [54] | |
| V149A | A | A | A | A | A | Pathogenicity in mice, antagonism of IFN induction | [55] |
3.3. Viral Replication Kinetics in MDCK Cells
3.4. Animal Study
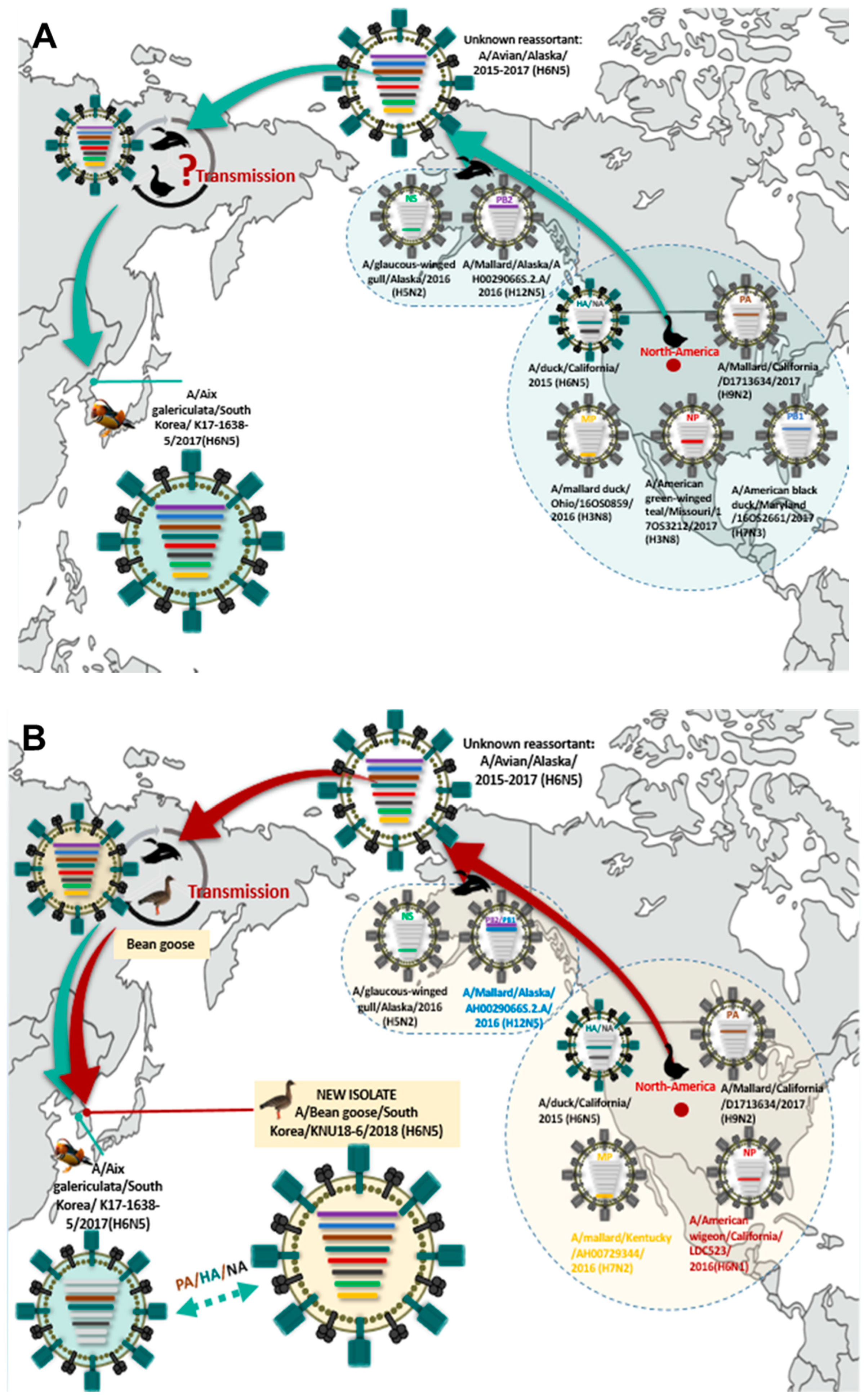
3.5. Hypothesis for Reassortment Event of A/Bean goose/South Korea/KNU18-6/2018(H6N5)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Prevention, C.D.C. Avian Influenza Type A Viruses. Available online: https://www.cdc.gov/flu/avianflu/influenza-a-virus-subtypes.htm (accessed on 19 April 2017).
- Olsen, B.; Munster, V.; Wallensten, A.; Waldenström, J.; Osterhaus, A.; Fouchier, R.A.M. Global patterns of influenza a virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, T.T.-Y.; Ip, H.S.; Ghedin, E.; Wentworth, D.; Halpin, R.A.; Stockwell, T.B.; Spiro, D.J.; Dusek, R.J.; Bortner, J.B.; Hoskins, J.; et al. Migratory flyway and geographical distance are barriers to the gene flow of influenza virus among North American birds. Ecol. Lett. 2012, 15, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Winker, K.; McCracken, K.G.; Gibson, D.D.; Pruett, C.L.; Meier, R.; Huettmann, F.; Wege, M.; Kulikova, I.V.; Zhuravlev, Y.N.; Perdue, M.L.; et al. Movements of birds and avian influenza from Asia into Alaska. Emerg. Infect. Dis. 2007, 13, 547–552. [Google Scholar] [CrossRef]
- Pearce, J.M.; Ramey, A.M.; Flint, P.L.; Koehler, A.; Fleskes, J.P.; Franson, J.; Hall, J.S.; Derksen, D.V.; Ip, H.S. Avian influenza at both ends of a migratory flyway: Characterizing viral genomic diversity to optimize surveillance plans for North America. Evol. Appl. 2009, 2, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Ramey, A.M.; Pearce, J.M.; Flint, P.L.; Ip, H.S.; Derksen, D.V.; Franson, J.; Petrula, M.J.; Scotton, B.D.; Sowl, K.M.; Wege, M.L.; et al. Intercontinental reassortment and genomic variation of low pathogenic avian influenza viruses isolated from northern pintails (Anas acuta) in Alaska: Examining the evidence through space and time. Virology 2010, 401, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheon, S.-H.; Lee, Y.-N.; Kang, S.-I.; Kye, S.-J.; Lee, E.-K.; Heo, G.-B.; Lee, M.-H.; Kim, J.W.; Lee, K.-N.; Son, H.-M.; et al. Genetic evidence for the intercontinental movement of avian influenza viruses possessing North American-origin nonstructural gene allele B into South Korea. Infect. Genet. Evol. 2018, 66, 18–25. [Google Scholar] [CrossRef]
- Jeong, S.; Lee, D.-H.; Kim, Y.-J.; Lee, S.-H.; Cho, A.Y.; Noh, J.-Y.; Tseren-Ochir, E.-O.; Jeong, J.-H.; Song, C.-S. Introduction of Avian Influenza A (H6N5) Virus into Asia from North America by Wild Birds. Emerg. Infect. Dis. 2019, 25, 2138–2140. [Google Scholar] [CrossRef]
- Zou, S.; Gao, R.; Zhang, Y.; Li, X.; Chen, W.; Bai, T.; Dong, L.; Wang, D.; Shu, Y. Molecular characterization of H6 subtype influenza viruses in southern China from 2009 to 2011. Emerg. Microbes Infect. 2016, 5, e73. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Sun, H.; Li, J.; Hou, G.; Wang, S.; Liu, S.; Zhuang, Q.; Cheng, S.; Chen, J.; Jiang, W. Molecular epidemiological survey and complete genomic phylogenetic analysis of H6 subtype avian influenza viruses in poultry in China from 2011 to 2016. Infect. Genet. Evol. 2018, 65, 91–95. [Google Scholar] [CrossRef]
- Li, J.; Quan, C.; Xie, Y.; Ke, C.; Nie, Y.; Chen, Q.; Hu, T.; Chen, J.; Wong, G.; Wang, Q.; et al. Continued reassortment of avian H6 influenza viruses from Southern China, 2014–2016. Transbound. Emerg. Dis. 2019, 66, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Nam, J.-H.; Kim, E.-H.; Song, D.; Choi, Y.K.; Kim, J.-K.; Poo, H. Emergence of mammalian species-infectious and -pathogenic avian influenza H6N5 virus with no evidence of adaptation. J. Virol. 2011, 85, 13271–13277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuong, H.T.; Nguyen, N.M.; Sung, H.W.; Park, H.; Yeo, S.-J. Genetic Characterization of Avian Influenza A (H11N9) Virus Isolated from Mandarin Ducks in South Korea in 2018. Viruses 2020, 12, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killian, M.L. Hemagglutination assay for influenza virus. Methods Mol. Biol. 2014, 1161, 3–9. [Google Scholar] [PubMed]
- World Health Organization. Geneva Switzerland. Available online: https://www.who.int/influenza/gisrs_laboratory/WHO_information_for_the_molecular_detection_of_influenza_viruses_20171023_Final (accessed on 23 November 2017).
- Hebert, P.D.; Stoeckle, M.Y.; Zemlak, T.S.; Francis, C.M. Identification of birds through DNA barcodes. PLoS Biol. 2004, 2, e312. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.-J.; Than, D.-D.; Park, H.-S.; Sung, H.W.; Park, H. Molecular Characterization of a Novel Avian Influenza A (H2N9) Strain Isolated from Wild Duck in Korea in 2018. Viruses 2019, 11, 1046. [Google Scholar] [CrossRef] [Green Version]
- Ambardar, S.; Gupta, R.; Trakroo, D.; Lal, R.; Vakhlu, J. High Throughput Sequencing: An. Overview of Sequencing Chemistry. Indian J. Microbiol. 2016, 56, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- WHO. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization Global Influenza Surveillance Network: Geneva, Switzerland, 2011. [Google Scholar]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Karlsson, E.A.; Ciuoderis, K.A.; Cortez, V.; Marvin, S.A.; Hamilton-West, C.; Schultz-Cherry, S.; Osorio, J.E. Avian H11 influenza virus isolated from domestic poultry in a Colombian live animal market. Emerg. Microbes Infect. 2016, 5, e121. [Google Scholar] [CrossRef]
- The University of IOWA. 2018. Available online: https://animal.research.uiowa.edu/iacuc-guidelines-anesthesia (accessed on 6 September 2018).
- Diederich, S.; Berhane, Y.; Embury-Hyatt, C.; Hisanaga, T.; Handel, K.; Cottam-Birt, C.; Ranadheera, C.; Kobasa, D.; Pasick, J. Hemagglutinin-Neuraminidase Balance Influences the Virulence Phenotype of a Recombinant H5N3 Influenza A Virus Possessing a Polybasic HA0 Cleavage Site. J. Virol. 2015, 89, 10724–10734. [Google Scholar] [CrossRef] [Green Version]
- Lazniewski, M.; Dawson, W.K.; Szczepińska, T.; Plewczynski, D. The structural variability of the influenza A hemagglutinin receptor-binding site. Brief. Funct. Genom. 2018, 17, 415–427. [Google Scholar]
- Matsuoka, Y.; Swayne, D.E.; Thomas, C.; Rameix-Welti, M.-A.; Naffakh, N.; Warnes, C.; Altholtz, M.; Donis, R.; Subbarao, K. Neuraminidase Stalk Length and Additional Glycosylation of the Hemagglutinin Influence the Virulence of Influenza H5N1 Viruses for Mice. J. Virol. 2009, 83, 4704–4708. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.-S.; Jeong, J.H.; Kwon, J.J.; Ahn, S.J.; Lloren, K.K.S.; Kwon, H.-I.; Chae, H.B.; Hwang, J.; Kim, M.H.; Kim, C.-J.; et al. Screening for Neuraminidase Inhibitor Resistance Markers among Avian Influenza Viruses of the N4, N5, N6, and N8 Neuraminidase Subtypes. J. Virol. 2017, 92, e01580-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Su, S.; Smith, D.K.; He, S.; Zheng, Y.; Shao, Z.; Ma, J.; Zhu, H.; Zhang, G. A Combination of HA and PA Mutations Enhances Virulence in a Mouse-Adapted H6N6 Influenza A Virus. J. Virol. 2014, 88, 14116–14125. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Cheng, K.; Xin, Y.; Sun, W.; Li, X.; Huang, J.; Zhang, K.; Yang, S.; Wang, T.; Zheng, X.; et al. Multiple amino acid substitutions involved in the adaptation of H6N1 avian influenza virus in mice. Vet. Microbiol. 2014, 174, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Y.; Hu, Y.; Chang, G.; Sun, W.; Yang, Y.; Kang, X.; Wu, X.; Zhu, Q. PB1-mediated virulence attenuation of H5N1 influenza virus in mice is associated with PB2. J. Gen. Virol. 2011, 92 Pt 6, 1435–1444. [Google Scholar] [CrossRef]
- Li, J.; Ishaq, M.; Prudence, M.; Xi, X.; Hu, T.; Liu, Q.; Guo, D. Single mutation at the amino acid position 627 of PB2 that leads to increased virulence of an H5N1 avian influenza virus during adaptation in mice can be compensated by multiple mutations at other sites of PB2. Virus Res. 2009, 144, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Prokopyeva, E.A.; Sobolev, I.A.; Prokopyev, M.V.; Shestopalov, A.M. Adaptation of influenza A (H1N1)pdm09 virus in experimental mouse models. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 39, 265–271. [Google Scholar] [CrossRef]
- Leung, B.W.; Chen, H.; Brownlee, G.G. Correlation between polymerase activity and pathogenicity in two duck H5N1 influenza viruses suggests that the polymerase contributes to pathogenicity. Virology 2010, 401, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Salomon, R.; Franks, J.; Govorkova, E.A.; Ilyushina, N.A.; Yen, H.L.; Hulse-Post, D.J.; Humberd, J.; Trichet, M.; Rehg, J.E.; Webby, R.J.; et al. The polymerase complex genes contribute to the high virulence of the human H5N1 influenza virus isolate A/Vietnam/1203/04. J. Exp. Med. 2006, 203, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Govorkova, E.A.; Rehg, J.E.; Krauss, S.; Yen, H.-L.; Guan, Y.; Peiris, M.; Nguyen, T.D.; Hanh, T.H.; Puthavathana, P.; Long, H.T.; et al. Lethality to ferrets of H5N1 influenza viruses isolated from humans and poultry in 2004. J. Virol. 2005, 79, 2191–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, G.; Abram, M.; Keiner, B.; Wagner, R.; Klenk, H.D.; Stech, J. Differential polymerase activity in avian and mammalian cells determines host range of influenza virus. J. Virol. 2007, 81, 9601–9604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.-D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. USA 2005, 102, 18590–18595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ágnes, F.; Loucaides, E.M.; Mura, M.; Wise, H.M.; Barclay, W.S.; Digard, P. Influence of PB2 host-range determinants on the intranuclear mobility of the influenza A virus polymerase. J. Gen. Virol. 2011, 92 Pt 7, 1650–1661. [Google Scholar]
- Hulse-Post, D.J.; Franks, J.; Boyd, K.; Salomon, R.; Hoffmann, E.; Yen, H.-L.; Webby, R.J.; Walker, D.; Nguyen, T.D.; Webster, R.G. Molecular changes in the polymerase genes (PA and PB1) associated with high pathogenicity of H5N1 influenza virus in mallard ducks. J. Virol. 2007, 81, 8515–8524. [Google Scholar] [CrossRef] [Green Version]
- Taubenberger, J.K.; Reid, A.H.; Lourens, R.M.; Wang, R.; Jin, G.; Fanning, T.G. Characterization of the 1918 influenza virus polymerase genes. Nature 2005, 437, 889–893. [Google Scholar] [CrossRef]
- Wei, K.; Sun, H.; Sun, Z.; Sun, Y.; Kong, W.; Pu, J.; Ma, G.; Yin, Y.; Yang, H.; Guo, X.; et al. Influenza A virus acquires enhanced pathogenicity and transmissibility after serial passages in swine. J. Virol. 2014, 88, 11981–11994. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Hu, W.-B.; Xu, K.; He, Y.-X.; Wang, T.-Y.; Chen, Z.; Li, T.-X.; Liu, J.-H.; Buchy, P.; Sun, B. Amino acids 473V and 598P of PB1 from an avian-origin influenza A virus contribute to polymerase activity, especially in mammalian cells. J. Gen. Virol. 2012, 93 Pt 3, 531–540. [Google Scholar] [CrossRef]
- Schmolke, M.; Manicassamy, B.; Pena, L.; Sutton, T.; Hai, R.; Varga, Z.T.; Hale, B.G.; Steel, J.; Perez, D.R.; García-Sastre, A. Differential contribution of PB1-F2 to the virulence of highly pathogenic H5N1 influenza A virus in mammalian and avian species. PLoS Pathog. 2011, 7, e1002186. [Google Scholar] [CrossRef] [Green Version]
- Ozawa, M.; Basnet, S.; Burley, L.M.; Neumann, G.; Hatta, M.; Kawaoka, Y. Impact of amino acid mutations in PB2, PB1-F2, and NS1 on the replication and pathogenicity of pandemic (H1N1) 2009 influenza viruses. J. Virol. 2011, 85, 4596–4601. [Google Scholar] [CrossRef] [Green Version]
- Yamayoshi, S.; Yamada, S.; Fukuyama, S.; Murakami, S.; Zhao, D.; Uraki, R.; Watanabe, T.; Tomita, Y.; Macken, C.; Neumann, G.; et al. Virulence-affecting amino acid changes in the PA protein of H7N9 influenza A viruses. J. Virol. 2014, 88, 3127–3134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, K.; Liu, G.; Chen, Z.; Gao, Z.; Zhao, L.; Jin, T.; Yu, X.; Chen, Q. Deep sequencing reveals the viral adaptation process of environment-derived H10N8 in mice. Infect. Genet. Evol. 2016, 37, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Cooper, L.; Xu, X.; Thompson, W.; Krauss, S.; Guan, Y.; Zhou, N.; Klimov, A.; Cox, N.; Webster, R.; et al. Molecular changes associated with the transmission of avian influenza a H5N1 and H9N2 viruses to humans. J. Med. Virol. 2002, 66, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zou, X.; Zhou, J.; Tang, J.; Shu, Y. Residues 41V and/or 210D in the NP protein enhance polymerase activities and potential replication of novel influenza (H7N9) viruses at low temperature. Virol. J. 2015, 12, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Chen, H.; Huang, J.; Chen, Y.; Gu, M.; Wang, X.; Hu, S.; Liu, X.; Liu, X. A nonpathogenic duck-origin H9N2 influenza A virus adapts to high pathogenicity in mice. Arch. Virol. 2014, 159, 2243–2252. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zheng, W.; Hou, L.; Chen, C.; Fan, W.; Qu, H.; Jiang, J.; Liu, J.; Gao, G.F.; Zhou, J.; et al. Differential nucleocytoplasmic shuttling of the nucleoprotein of influenza a viruses and association with host tropism. Cell. Microbiol. 2017, 19, e12692. [Google Scholar] [CrossRef] [Green Version]
- Ou, X.; Chen, F.; Zhang, R.; Chen, J.; Liu, R.; Sun, B. Analysis of the full-length genome of a novel strain of the H7N9 avian influenza virus. Exp. Ther. Med. 2014, 7, 1369–1375. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Deng, G.; Song, J.; Tian, G.; Suo, Y.; Jiang, Y.; Guan, Y.; Bu, Z.; Kawaoka, Y.; Chen, H. Two amino acid residues in the matrix protein M1 contribute to the virulence difference of H5N1 avian influenza viruses in mice. Virology 2009, 384, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Jiao, P.; Tian, G.; Li, Y.; Deng, G.; Jiang, Y.; Liu, C.; Liu, W.; Bu, Z.; Kawaoka, Y.; Chen, H. A single-amino-acid substitution in the NS1 protein changes the pathogenicity of H5N1 avian influenza viruses in mice. J. Virol. 2008, 82, 1146–1154. [Google Scholar] [CrossRef] [Green Version]
- Min, J.-Y.; Li, S.; Sen, G.C.; Krug, R.M. A site on the influenza A virus NS1 protein mediates both inhibition of PKR activation and temporal regulation of viral RNA synthesis. Virology 2007, 363, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Jiang, Y.; Jiao, P.; Wang, A.; Zhao, F.; Tian, G.; Wang, X.; Yu, K.; Bu, Z.; Chen, H. The NS1 gene contributes to the virulence of H5N1 avian influenza viruses. J. Virol. 2006, 80, 11115–11123. [Google Scholar] [CrossRef] [Green Version]
- Ramey, A.M.; Reeves, A.B.; Donnelly, T.; Poulson, R.L.; Stallknecht, D.E. Introduction of Eurasian-Origin Influenza A(H8N4) Virus into North America by Migratory Birds. Emerg. Infect. Dis. 2018, 24, 1950–1953. [Google Scholar] [CrossRef] [PubMed]
- Ramey, A.M.; Reeves, A.B.; Sonsthagen, S.A.; TeSlaa, J.; Nashold, S.; Donnelly, T.; Casler, B.; Hall, J.S. Dispersal of H9N2 influenza A viruses between East Asia and North America by wild birds. Virology 2015, 482, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Wille, M.; Robertson, G.J.; Whitney, H.; Bishop, M.A.; Runstadler, J.A.; Lang, A.S. Extensive geographic mosaicism in avian influenza viruses from gulls in the northern hemisphere. PLoS ONE 2011, 6, e20664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramey, A.M.; Pearce, J.M.; Reeves, A.B.; Poulson, R.L.; Dobson, J.; Lefferts, B.; Spragens, K.; Stallknecht, D.E. Surveillance for Eurasian-origin and intercontinental reassortant highly pathogenic influenza A viruses in Alaska, spring and summer 2015. Virol. J. 2016, 13, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonthabenjawan, N.; Cardona, C.J.; Amonsin, A.; Sreevatsan, S. Time-space analysis of highly pathogenic avian influenza H5N2 outbreak in the US. Virol. J. 2016, 13, 147. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Tanikawa, T.; Uchida, Y.; Takemae, N.; Kanehira, K.; Tsunekuni, R. Intracontinental and intercontinental dissemination of Asian H5 highly pathogenic avian influenza virus (clade 2.3.4.4) in the winter of 2014-2015. Rev. Med. Virol. 2015, 25, 388–405. [Google Scholar] [CrossRef]
- Ip, H.S.; Kim-Torchetti, M.; Crespo, R.; Kohrs, P.; Debruyn, P.; Mansfield, K.G.; Baszler, T.; Badcoe, L.; Bodenstein, B.; Shearn-Bochsler, V.; et al. Novel Eurasian highly pathogenic avian influenza A H5 viruses in wild birds, Washington, USA, 2014. Emerg. Infect. Dis. 2015, 21, 886–890. [Google Scholar] [CrossRef]
- Okuya, K.; Kanazawa, N.; Kanda, T.; Kuwahara, M.; Matsuu, A.; Horie, M.; Toda, S.; Ozawa, M.; Masatani, T. Genetic characterization of an avian H4N6 influenza virus isolated from the Izumi plain, Japan. Microbiol. Immunol. 2017, 61, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Lee, H.-J.; Lee, Y.-N.; Park, J.-K.; Lim, T.-H.; Kim, M.-S.; Youn, H.-N.; Lee, J.-B.; Park, S.-Y.; Choi, I.-S.; et al. Evidence of intercontinental transfer of North American lineage avian influenza virus into Korea. Infect. Genet. Evol. 2011, 11, 232–236. [Google Scholar] [CrossRef] [PubMed]
- The Global Register of Migratory Species. Available online: http://www.groms.de/groms/JPGs/Small_Ones/index.html (accessed on 30 April 2019).
- Gillim-Ross, L.; Santos, C.; Chen, Z.; Aspelund, A.; Yang, C.-F.; Ye, D.; Jin, H.; Kemble, G.; Subbarao, K. Avian influenza h6 viruses productively infect and cause illness in mice and ferrets. J. Virol. 2008, 82, 10854–10863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.-H.; Yang, J.-R.; Wu, H.-S.; Chang, M.-C.; Lin, J.-S.; Lin, C.-Y.; Liu, Y.-L.; Lo, Y.-C.; Yang, C.-H.; Chuang, J.-H.; et al. Human infection with avian influenza A H6N1 virus: An epidemiological analysis. Lancet Respir. Med. 2013, 1, 771–778. [Google Scholar] [CrossRef]
- Shi, W.; Shi, Y.; Wu, Y.; Liu, D.; Gao, G.F. Origin and molecular characterization of the human-infecting H6N1 influenza virus in Taiwan. Protein Cell 2013, 4, 846–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Li, J.; Hu, J.; Jiang, D.; Xing, C.; Zhan, T.; Liu, X. Genetic analysis and biological characteristics of different internal gene origin H5N6 reassortment avian influenza virus in China in 2016. Vet. Microbiol. 2018, 219, 200–211. [Google Scholar] [CrossRef]
- Jiao, P.; Cui, J.; Song, Y.; Song, H.; Zhao, Z.; Wu, S.; Qu, N.; Wang, N.; Ouyang, G.; Jiao, P. New Reassortant H5N6 Highly Pathogenic Avian Influenza Viruses in Southern China, 2014. Front. Microbiol. 2016, 7, 754. [Google Scholar] [CrossRef]
- Hu, C.; Li, X.; Zhu, C.; Zhou, F.; Tang, W.; Wu, D.; Li, Z.; Zhou, L.; Liu, J.; Wei, X.; et al. Co-circulation of multiple reassortant H6 subtype avian influenza viruses in wild birds in eastern China, 2016–2017. Virol. J. 2020, 17, 62. [Google Scholar] [CrossRef]
- Yu, Z.; Sun, W.; Li, X.; Chen, Q.; Chai, H.; Gao, X.; Guo, J.; Zhang, K.; Wang, T.; Feng, N.; et al. Adaptive amino acid substitutions enhance the virulence of a reassortant H7N1 avian influenza virus isolated from wild waterfowl in mice. Virology 2015, 476, 233–239. [Google Scholar] [CrossRef] [Green Version]





| Gene | GenBank Accession # of K6 | Strain Having Highest Nucleotide Identity with K6 | Genetic Identity between K6 and K17 | |
|---|---|---|---|---|
| Strain | GenBank Accession # (% Genetic Identity) | |||
| PB2 | MN749559 | A/Mallard/Alaska/AH0029066S.1.A/2016(H12N5) | MN254630.1 (99.34%) | 99.21% |
| PB1 | MN749560 | A/Mallard /Alaska/AH0029066S.2.A /2016(H12N5) | MN254483.1 (99.56%) | 94.64% |
| PA | MN749561 | A/Aix galericulata/South Korea/K17-1638-5/2017(H6N5) | MK830102.1 (99.49%) | 99.49% |
| HA | MN749562 | A/Aix galericulata/South Korea/K17-1638-5/2017(H6N5) | EPI1526581 99.30% | 99.30% |
| NP | MN749563 | A/lesser scaup/Wisconsin/17OS5811/2017(H6N1) | EPI1537365 99.67% | 96.32% |
| NA | MN749564 | A/Aix galericulata/South Korea/K17-1638-5/2017(H6N5) | EPI1526586 99.16% | 99.16% |
| MP | MN749565 | A/mallard/Kentucky/AH00729344/2016(H7N2) | EPI952771 99.59% | 97.66% |
| NS | MN749566 | A/blue-wingedteal/Missouri/17OS3211/2017(A/H3N1) | EPI1537413 99.52% | 99.40% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, N.M.; Sung, H.W.; Yun, K.-J.; Park, H.; Yeo, S.-J. Genetic Characterization of a Novel North American-Origin Avian Influenza A (H6N5) Virus Isolated from Bean Goose of South Korea in 2018. Viruses 2020, 12, 774. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070774
Nguyen NM, Sung HW, Yun K-J, Park H, Yeo S-J. Genetic Characterization of a Novel North American-Origin Avian Influenza A (H6N5) Virus Isolated from Bean Goose of South Korea in 2018. Viruses. 2020; 12(7):774. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070774
Chicago/Turabian StyleNguyen, Ngoc Minh, Haan Woo Sung, Ki-Jung Yun, Hyun Park, and Seon-Ju Yeo. 2020. "Genetic Characterization of a Novel North American-Origin Avian Influenza A (H6N5) Virus Isolated from Bean Goose of South Korea in 2018" Viruses 12, no. 7: 774. https://0-doi-org.brum.beds.ac.uk/10.3390/v12070774