The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 2: Clinical Signs and Gross Pathology


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Inoculum
2.3. Statistics
3. Results
3.1. Incidence of Stillbirths, Preweaning Deaths and Mummified Pigs
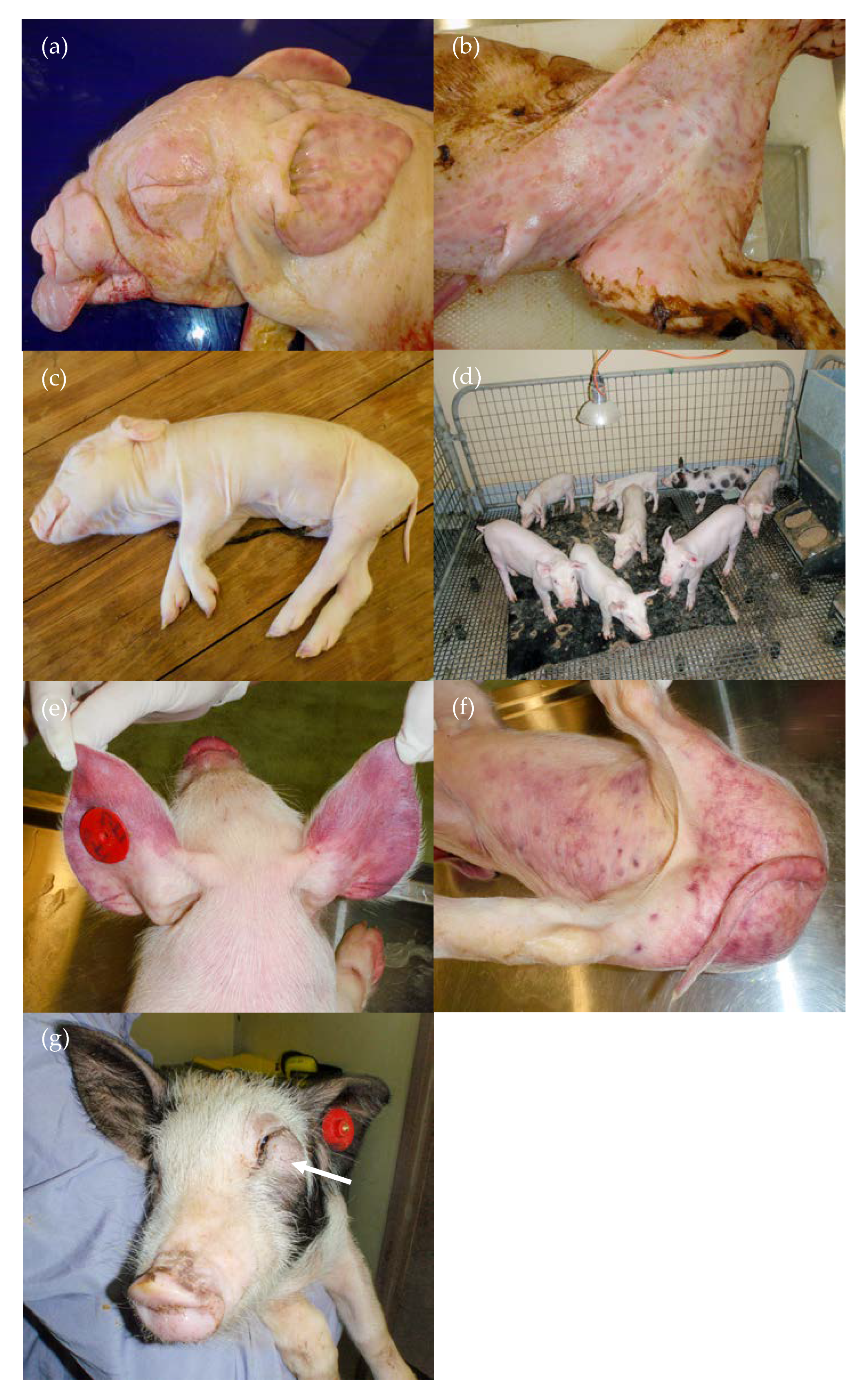
3.2. Clinical Signs
3.2.1. Group D35
3.2.2. Group D55
3.2.3. Group D75
3.2.4. Group D90
3.2.5. Uninfected Litters
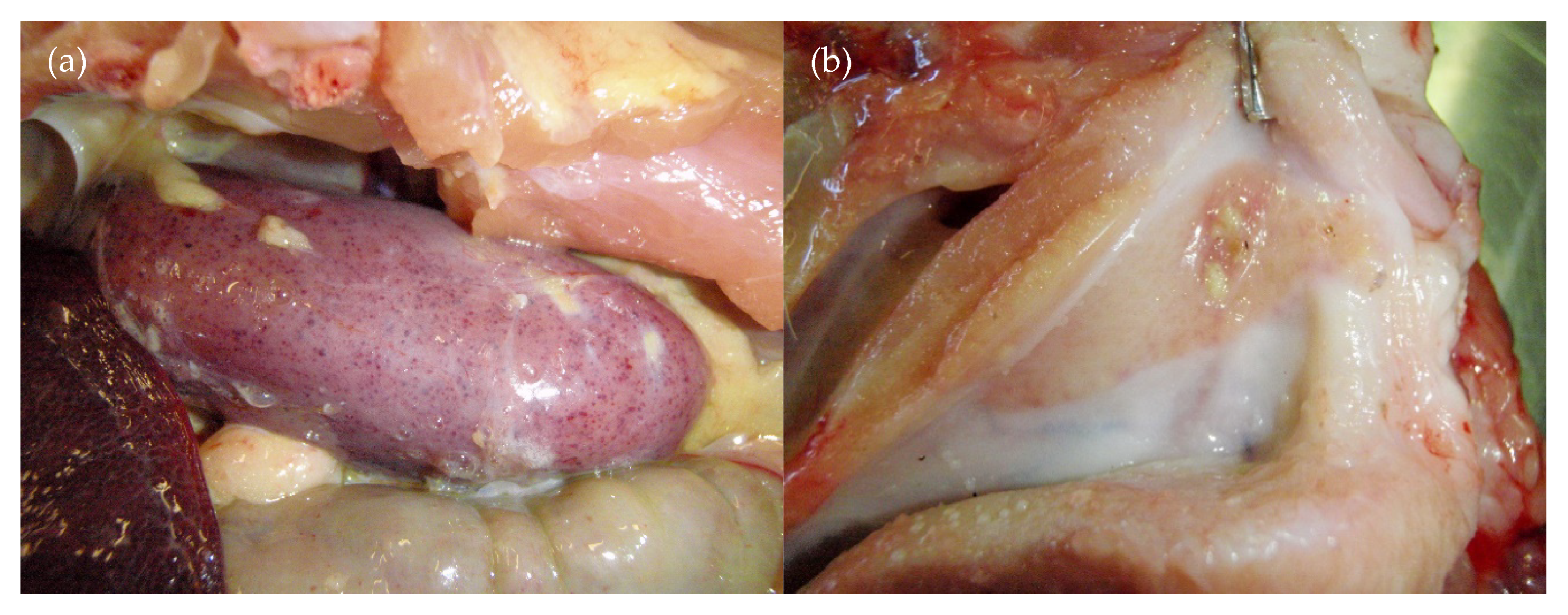
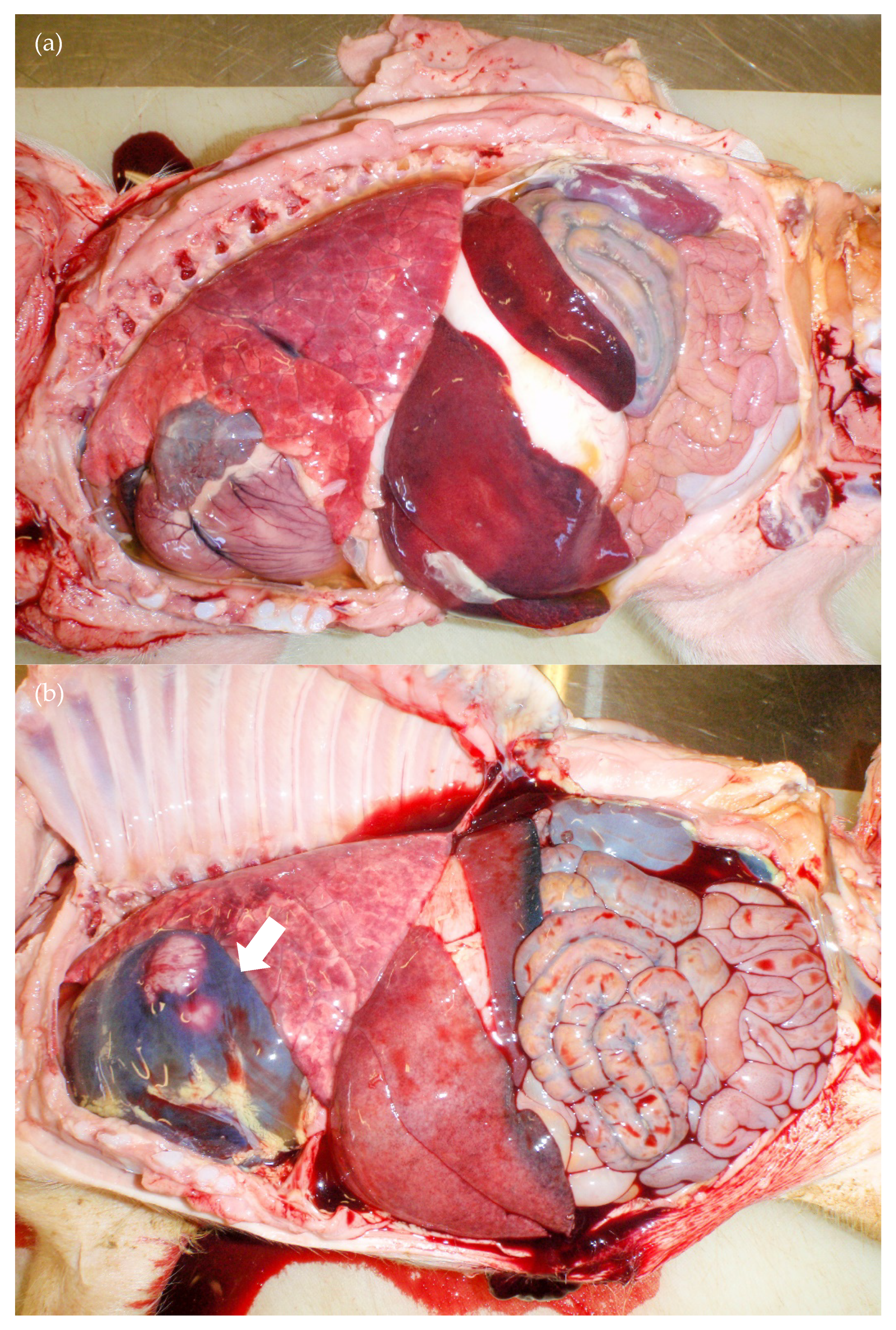
3.3. Necropsy Findings
3.3.1. Group D35
3.3.2. Group D55
3.3.3. Group D75
3.3.4. Group D90
3.3.5. Uninfected Litters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McOrist, S.; Thornton, E.; Peake, A.; Walker, R.; Robson, S.; Finlaison, D.; Kirkland, P.; Reece, R.; Ross, A.; Walker, K.; et al. An infectious myocarditis syndrome affecting late-term and neonatal piglets. Aust. Vet. J. 2004, 82, 509–511. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Frost, M.J.; Finlaison, D.S.; King, K.R.; Ridpath, J.F.; Gu, X. Identification of a novel virus in pigs-Bungowannah virus: A possible new species of pestivirus. Virus Res. 2007, 129, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hause, B.M.; Collin, E.A.; Peddireddi, L.; Yuan, F.; Chen, Z.; Hesse, R.A.; Gauger, P.C.; Clement, T.; Fang, Y.; Anderson, G. Discovery of a novel putative atypical porcine pestivirus in pigs in the USA. J. Gen. Virol. 2015, 96, 2994–2998. [Google Scholar] [CrossRef] [PubMed]
- Lamp, B.; Schwarz, L.; Hogler, S.; Riedel, C.; Sinn, L.; Rebel-Bauder, B.; Weissenbock, H.; Ladinig, A.; Rumenapf, T. Novel Pestivirus Species in Pigs, Austria, 2015. Emerg. Infect. Dis. 2017, 23, 1176–1179. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V.; Liess, B. Pathogenesis of intrauterine infections with bovine viral diarrhea virus. Vet. Clin. N. Am. Food Anim. Pract. 1995, 11, 477–487. [Google Scholar] [CrossRef]
- Roeder, P.L.; Jeffrey, M.; Cranwell, M.P. Pestivirus fetopathogenicity in cattle: Changing sequelae with fetal maturation. Vet. Rec. 1986, 118, 44–48. [Google Scholar] [CrossRef]
- Baker, J.C. The Clinical Manifestations of Bovine Viral Diarrhea Infection. Vet. Clin. N. Am. Food A 1995, 11, 425–445. [Google Scholar] [CrossRef]
- McGowan, M.R.; Kirkland, P.D. Early reproductive loss due to bovine pestivirus infection. Br. Vet. J. 1995, 151, 263–270. [Google Scholar] [CrossRef]
- Garcia-Perez, A.L.; Minguijon, E.; Estevez, L.; Barandika, J.F.; Aduriz, G.; Juste, R.A.; Hurtado, A. Clinical and laboratorial findings in pregnant ewes and their progeny infected with Border disease virus (BDV-4 genotype). Res. Vet. Sci. 2009, 86, 345–352. [Google Scholar] [CrossRef]
- Arruda, B.L.; Arruda, P.H.; Magstadt, D.R.; Schwartz, K.J.; Dohlman, T.; Schleining, J.A.; Patterson, A.R.; Visek, C.A.; Victoria, J.G. Identification of a Divergent Lineage Porcine Pestivirus in Nursing Piglets with Congenital Tremors and Reproduction of Disease following Experimental Inoculation. PLoS ONE 2016, 11, e0150104. [Google Scholar] [CrossRef] [Green Version]
- Trautwein, G.; Hewicker, M.; Liess, B.; Orban, S.; Grunert, E. Studies on Transplacental Transmissibility of a Bovine Virus Diarrhoea (BVD) Vaccine Virus in Cattle III. Occurrence of Central Nervous System Malformations in Calves Born from Vaccinated Cows. J. Vet. Med. Ser. B 1986, 33, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, P.F. Border disease. In Infectious Diseases of Livestock, 2nd ed.; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press Southern Africa: Cape Town, South Africa, 2004; Volume 2, pp. 970–974. [Google Scholar]
- Van Oirschot, J.T. Experimental production of congenital persistent swine fever infections: I. Clinical, pathological and virological observations. Vet. Microbiol. 1979, 4, 117–132. [Google Scholar] [CrossRef]
- Paton, D.J.; Done, S.H. Congenital infection of pigs with ruminant-type pestiviruses. J. Comp. Pathol. 1994, 111, 151–163. [Google Scholar] [CrossRef]
- Van Oirschot, J.T.; Terpstra, C. A congenital persistent swine fever infection. I. Clinical and virological observations. Vet. Microbiol. 1977, 2, 121–132. [Google Scholar] [CrossRef]
- Terpstra, C.; Wensvoort, G. A congenital persistent infection of bovine virus diarrhoea virus in pigs: Clinical, virological and immunological observations. Vet. Q. 1997, 19, 97–101. [Google Scholar] [CrossRef]
- Young, G.A.; Kitchell, R.L.; Luedke, A.J.; Sautter, J.H. The effect of viral and other infections of the dam on fetal development in swine. I. Modified live hog cholera viruses; immunological, virological, and gross pathological studies. J. Am. Vet. Med. Assoc. 1955, 126, 165–171. [Google Scholar]
- Leforban, Y.; Vannier, P.; Cariolet, R. Pathogenicity of border disease and bovine viral diarrhoea for pig: Experimental study on the vertical and horizontal transmission of the viruses. In Proceedings of the 11th International Pig Veterinary Society Congress, Lausanne, Switzerland, 1–5 July 1990; p. 204. [Google Scholar]
- Postel, A.; Hansmann, F.; Baechlein, C.; Fischer, N.; Alawi, M.; Grundhoff, A.; Derking, S.; Tenhundfeld, J.; Pfankuche, V.M.; Herder, V.; et al. Presence of atypical porcine pestivirus (APPV) genomes in newborn piglets correlates with congenital tremor. Sci. Rep. 2016, 6, 27735. [Google Scholar] [CrossRef] [Green Version]
- Finlaison, D.S.; King, K.R.; Frost, M.J.; Kirkland, P.D. Field and laboratory evidence that Bungowannah virus, a recently recognised pestivirus, is the causative agent of the porcine myocarditis syndrome (PMC). Vet. Microbiol. 2009, 136, 259–265. [Google Scholar] [CrossRef]
- Finlaison, D.S.; Cook, R.W.; Srivastava, M.; Frost, M.J.; King, K.R.; Kirkland, P.D. Experimental infections of the porcine foetus with Bungowannah virus, a novel pestivirus. Vet. Microbiol. 2010, 144, 32–40. [Google Scholar] [CrossRef]
- Finlaison, D.S.; Kirkland, P.D. The outcome of porcine foetal infection with Bungowannah virus is dependent on the stage of gestation at which infection occurs. Part 1: Serology and virology. Viruses 2020, 12, 691. [Google Scholar] [CrossRef]
- Finlaison, D.S.; King, K.R.; Gabor, M.; Kirkland, P.D. An experimental study of Bungowannah virus infection in weaner aged pigs. Vet. Microbiol. 2012, 160, 245–250. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statisical Computing: Vienna, Austria, 2019; Available online: http://www.R-project.org (accessed on 1 April 2019).
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J. ASReml-R Reference Manual, release 3 ed.; Queensland Department of Primary Industries and Fisheries: Brisbane, Australia, 2009.
- Friendship, R.M.; Lumsden, J.H.; McMillan, I.; Wilson, M.R. Hematology and biochemistry reference values for Ontario swine. Can. J. Comp. Med. 1984, 48, 390–393. [Google Scholar] [PubMed]
- Finlaison, D.S. Studies of the Porcine Myocarditis Syndrome. Ph.D. Thesis, University of Sydney, Sydney, Australia, 2010. [Google Scholar]
- Finlaison, D.S.; Gestier, S.; Kirkland, P.D. The outcome of porcine foetal infection with Bungowannah virus is dependent on the stage of gestation at which infection occurs—Histopathology. in preparation.
- Frey, H.R.; Liess, B.; Richter-Reichhelm, H.B.; von Benten, K.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. I. Virological and serological studies. Zentralbl. Veterinarmed. B 1980, 27, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Meyer, H.; Liess, B.; Frey, H.R.; Hermanns, W.; Trautwein, G. Experimental transplacental transmission of hog cholera virus in pigs. IV. Virological and serological studies in newborn piglets. Zentralbl. Veterinarmed. B 1981, 28, 659–668. [Google Scholar] [CrossRef]
- Hermanns, W.; Trautwein, G.; Meyer, H.; Liess, B. Experimental transplacental transmission of hog cholera virus in pigs. V. Immunopathological findings in newborn pigs. Zentralbl. Veterinarmed. B 1981, 28, 669–683. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Sow Number (Litter ID) | Day of Gestation at Challenge | Day of Farrowing | Born Alive | Stillborn/AutolySing (%) | Mummified (%) | Total Litter Size | Preweaning Losses (%) | Number Weaned | Comments |
|---|---|---|---|---|---|---|---|---|---|---|
| D35 | 4558 (15) | 34 | 112 | 8 | 1 (7%) | 5 (36%) | 14 | 8 (100%) | 0 | |
| D35 | 4536 (17) | 34 | 114 | 7 | 6 (46%) | 0 (0%) | 13 | 2 (29%) | 5 | |
| D35 | 4540 (12) | 35 | 111 | 7 | 3 (25%) | 2 (17%) | 12 | 7 (100%) | 0 | |
| D35 | 4596 (16) | 35 | 114 | 4 | 4 (40%) | 2 (20%) | 10 | 3 (75%) | 1 | |
| D35 | 4570 (14) | 36 | 114 | 9 | 0 (0%) | 2 (18%) | 11 | 4 (44%) | 5 | |
| Mean | 7.0 | 24% | 18% | 12 | 70% | 2.2 | ||||
| D55 | 4261 (10) | 55 | 114 | 11 | 1 (8%) | 0 (0%) | 12 | 2 (18%) | 9 | |
| D55 | 325 (11) | 55 | 114 | 13 | 2 (13%) | 0 (0%) | 15 | 3 (23%) | 10 | Parity 1; pig 11-15 not infected |
| D55 | 4409 (8) | 56 | 114 | 6 | 2 (25%) | 0 (0%) | 8 | 0 (0%) | 6 | |
| D55 | 4268 (7) | 58 | 114 | 11 | 0 (0%) | 0 (0%) | 11 | 2 (18%) | 6 | 3 piglets savaged |
| Mean | 10.3 | 12% | 0% | 11.5 | 15% | 7.8 | ||||
| D75 | 4294 (3) | 73 | 114 | 11 | 0 | 0 | 11 | 3 (27%) | 8 | |
| D75 | 4300 (4) | 73 | 114 | 7 | 1 | 2 | 10 | 0 (0%) | 7 | |
| D75 | 4486 (5) | 73 | 114 | 10 | 1 | 0 | 11 | 0 (0%) | 0 2 | 10 piglets savaged/poor milk production |
| D75 | 4279 (1) | 76 | 110 | 11 | 3 | 0 | 14 | 0 (0%) | 0 2 | 11 piglets savaged |
| D75 | 4281 (2) | 76 | 114 | 7 | 5 | 0 | 12 | 0 (0%) | 7 | |
| Mean | 9.2 | 16% | 4% | 11.6 | 5% | 7.3 | ||||
| D90 | 739 (18) | 90 | 111 | 6 | 3 | 1 | 10 | 1 (17%) | 1 | 4 piglets savaged |
| D90 | 735 (22) | 90 | 113 | 9 | 2 | 0 | 11 | 5 (56%) | 4 | |
| D90 | 738 (23) | 90 | 114 | 10 | 1 | 0 | 11 | 0 (0%) | 10 | |
| D90 | 736 (20) | 91 | 112 | 9 | 3 | 0 | 12 | 2 (22%) | 7 | |
| D90 | 2622 (21) | 91 | 114 | 10 | 0 | 0 | 10 | 4 (40%) | 6 | |
| D90 | 2623 (19) | 92 | 113 | 8 | 0 | 0 | 8 | 3 (38%) | 5 | |
| Mean | 8.7 | 14% | 2% | 10.3 | 29% | 5.5 | ||||
| D35 | 35 (13) | 36 | 114 | 13 | 3 | 1 | 17 | 1 (8%) | 12 | NI 3; Parity 1 |
| D55 | 4260 (9) | 56 | 114 | 11 | 2 | 2 | 15 | 4 (38%) | 6 | NI; 1 death due to accident |
| D75 | 4292 (6) | 72 | 114 | 10 | 0 | 0 | 10 | 1 (10%) | 9 | NI |
| Mean | 11.3 | 10% | 6% | 14.0 | 18% | 9 |
| Clinical Signs | Necropsy Findings | |||||
|---|---|---|---|---|---|---|
| Day of Gestation at Challenge | Reproductive Effects 1 | Persistent Infections? 2 | Preweaning | Postweaning | Preweaning | Postweaning |
| 35 days | Born alive ↓↓↓ Stillbirths ↑↑ Mummified foetuses ↑↑↑ Preweaning deaths ↑↑↑ | Yes (>90%) | Purpura, subcutaneous oedema, absence of facial whiskers, weak-born pigs that did not suckle, neurological signs—abnormal behaviour | Stunting, pallor due to anaemia, conjunctivitis, subcutaneous facial skin mass, leukopaenia, thrombocytopaenia, purplish discolouration of the snout and ears, ecchymoses in the skin of the hind-quarter | Purpura, subcutaneous oedema, white foci in the cerebrum, orange/mottled liver, fibrinous adhesions in pleural and abdominal cavities (older animals) | Blood clots and serosanguinous fluid in abdomen/thoracic cavities, petechiae, red/haemorrhagic lymph nodes, ulcerated tonsils, facial mass |
| 55 days | Stillbirths ↑ | At birth, 18% were seronegative to Bungowannah virus and apparently persistently infected. The 3 seronegative animals retained postweaning later seroconverted | Melaena, pallor, ataxia | Stunting of apparently persistently infected pigs, occasional diarrhoea | Occasional fibrinous adhesions in thoracic and abdominal cavities | No significant findings |
| 75 days | Stillbirths ↑ | No | No disease observed | No disease observed | No significant findings | No significant findings |
| 90 days | Stillbirths ↑ Preweaning deaths ↑↑ | No | Sudden death, tachypnoea and dyspnoea | No disease observed | Cardiomegally, liver enlargement, fibrin tags in the thoracic and/or abdominal cavities, blood clots in abdominal cavity, pericardial/thoracic effusions, red/haemorrhagic mesenteric and/or inguinal lymph nodes | No significant findings |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Finlaison, D.S.; Kirkland, P.D. The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 2: Clinical Signs and Gross Pathology. Viruses 2020, 12, 873. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080873
Finlaison DS, Kirkland PD. The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 2: Clinical Signs and Gross Pathology. Viruses. 2020; 12(8):873. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080873
Chicago/Turabian StyleFinlaison, Deborah S., and Peter D. Kirkland. 2020. "The Outcome of Porcine Foetal Infection with Bungowannah Virus Is Dependent on the Stage of Gestation at Which Infection Occurs. Part 2: Clinical Signs and Gross Pathology" Viruses 12, no. 8: 873. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080873