Gammaherpesvirus in Cervid Species from Norway: Characterization of a New Virus in Wild and Semi-Domesticated Eurasian Tundra Reindeer (Rangifer tarandus tarandus)




Abstract
:1. Introduction
2. Materials and Methods
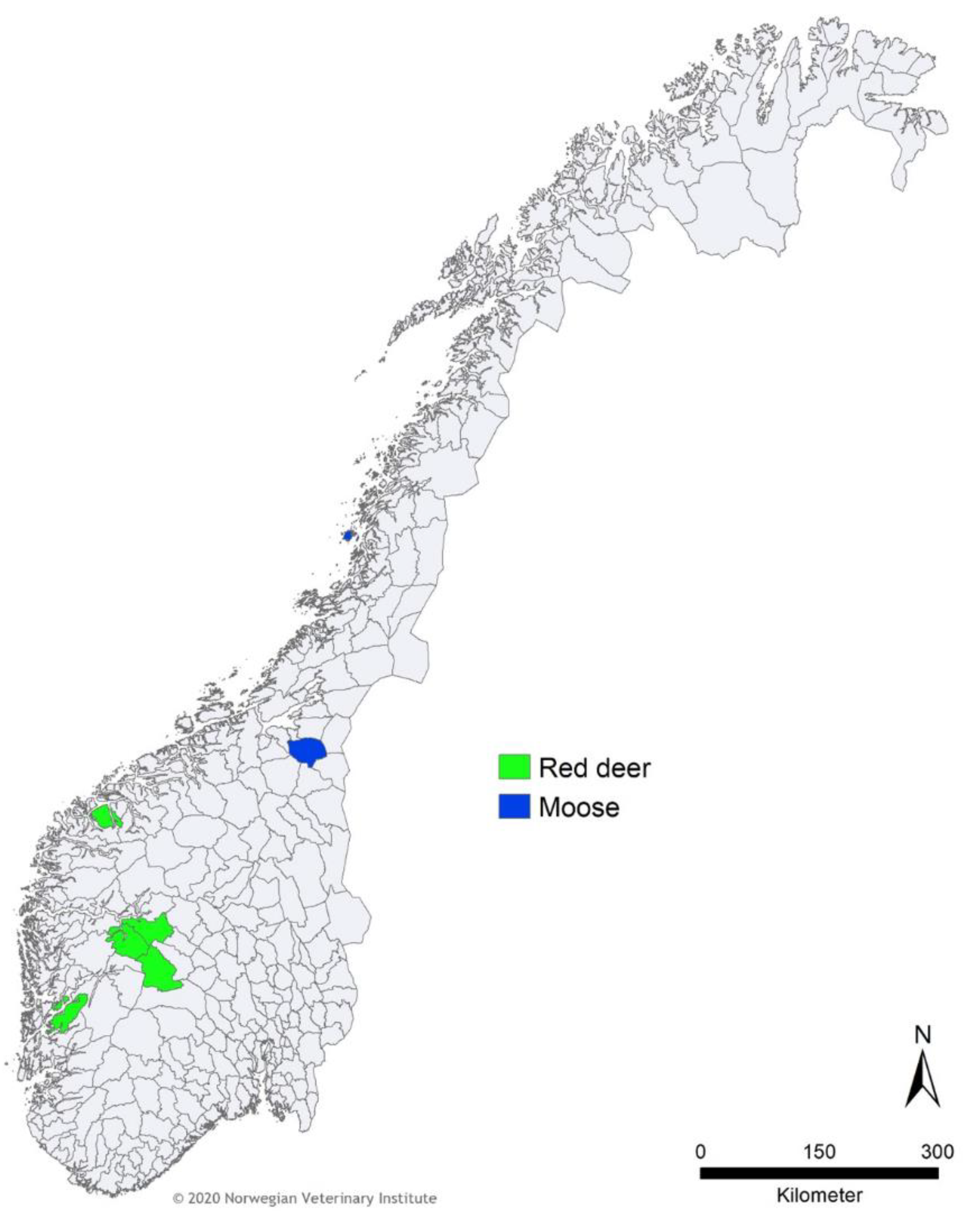
2.1. Samples
2.2. Molecular Diagnostics
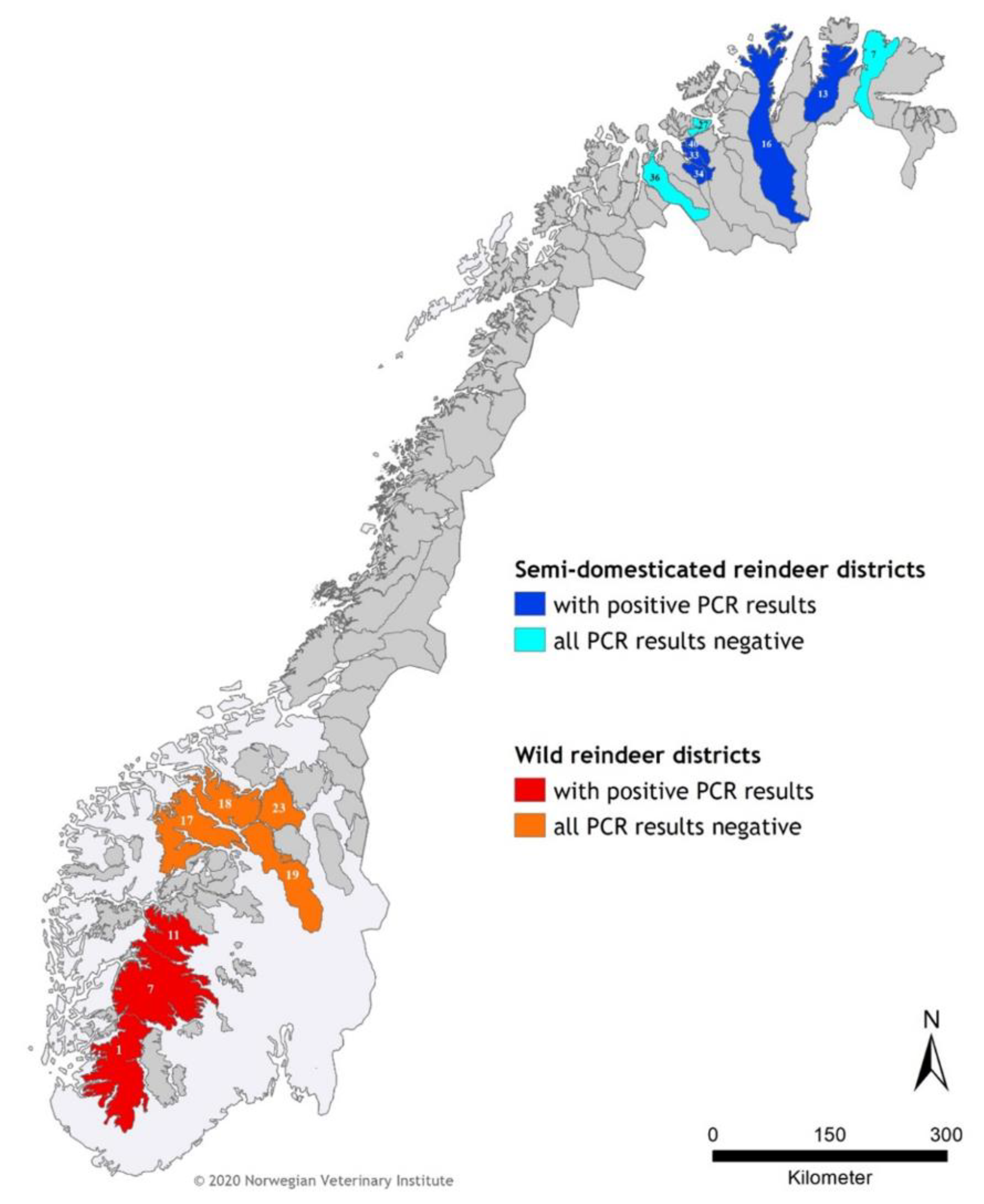
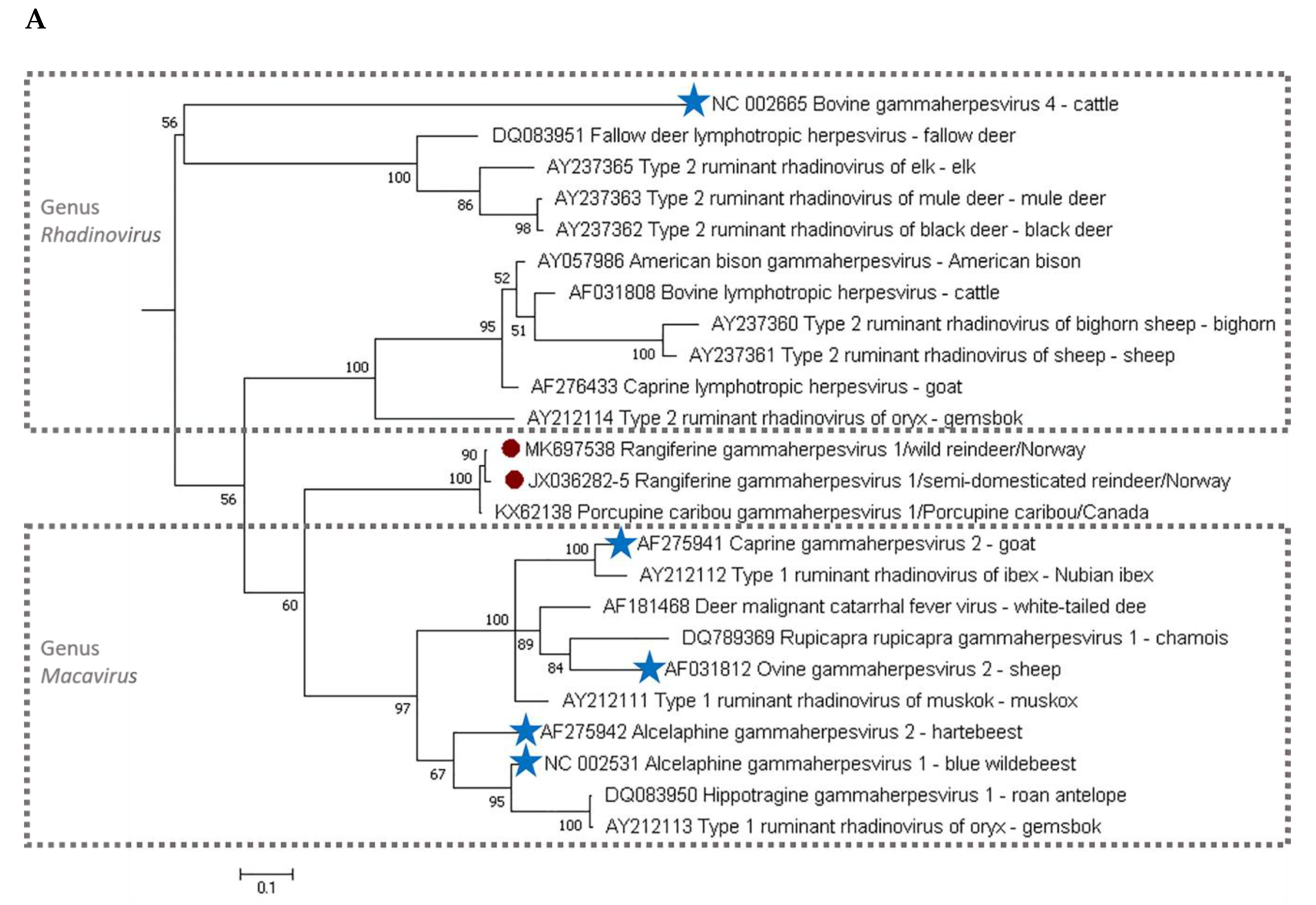
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Number | Organism | Common Designation |
|---|---|---|
| NC_020700.1 | Dama dama | Fallow deer |
| NC_007704.2 | Cervus elaphus | Red deer |
| NC_020684.1 | Capreolus capreolus | Roe deer |
| NC_020677.1 | Alces alces | Moose/elk |
| NC_015247.1 | Odocoileus virginianus | White-tailed deer |
| NC_020729.1 | Odocoileus hemionus | Mule deer |
| NC_007703.1 | Rangifer tarandus | Reindeer |
| NC_001941.1 | Ovis aries | Sheep |
| NC_005044.2 | Capra hircus | Goat |
| NC_020633.1 | Rupicapra rupicapra | Chamois |
| NC_020631.1 | Ovibus moschatus | Muskox |
| NC_020699.1. | Connochaetes taurinus | Blue wildebeest |
| NC_016421.1 | Oryx dammah | Scimitar oryx |
| AF492351.1 | Bos Taurus | Bovine/Cattle |
References
- ICTV. Virus Taxonomy: 2019. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/dsdna-viruses-2011/w/dsdna_viruses/91/herpesviridae (accessed on 10 July 2020).
- Smits, S.L.; Schapendonk, C.M.; van Leeuwen, M.; Kuiken, T.; Bodewes, R.; Stalin Raj, V.; Haagmans, B.L.; das Neves, C.G.; Tryland, M.; Osterhaus, A.D. Identification and characterization of two novel viruses in ocular infections in reindeer. PLoS ONE 2013, 8, e69711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widén, F.; das Neves, C.G.; Ruiz-Fons, F.; Reid, H.W.; Kuiken, T.; Gavier-Widén, D.; Kaleta, E.F. Herpesvirus Infections. In Infectious Diseases of Wild Mammals and Birds in Europe; Wiley: Hoboken, NJ, USA, 2012; pp. 1–36. [Google Scholar]
- das Neves, C.G.; Rimstad, E.; Tryland, M. Cervid herpesvirus 2 causes respiratory and fetal infections in semidomesticated reindeer. J. Clin. Microbiol. 2009, 47, 1309–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiry, J.; Dams, L.; Muylkens, B.; Thiry, E. Isolation of cervid herpesvirus 1 from the genital tract of a farmed red deer in Northern France. Vet. J. 2011, 187, 282–283. [Google Scholar] [CrossRef]
- Tryland, M.; das Neves, C.G.; Sunde, M.; Mork, T. Cervid herpesvirus 2, the primary agent in an outbreak of infectious keratoconjunctivitis in semidomesticated reindeer. J. Clin. Microbiol. 2009, 47, 3707–3713. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellet, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dyer, N.; Keller, J.; Crawford, T.B. Newly recognized herpesvirus causing malignant catarrhal fever in white-tailed deer (Odocoileus virginianus). J. Clin. Microbiol. 2000, 38, 1313–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Gailbreath, K.; Bender, L.C.; West, K.; Keller, J.; Crawford, T.B. Evidence of three new members of malignant catarrhal fever virus group in muskox (Ovibos moschatus), Nubian ibex (Capra nubiana), and gemsbok (Oryx gazella). J. Wildl. Dis. 2003, 39, 875–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klieforth, R.; Maalouf, G.; Stalis, I.; Terio, K.; Janssen, D.; Schrenzel, M. Malignant catarrhal fever-like disease in Barbary red deer (Cervus elaphus barbarus) naturally infected with a virus resembling alcelaphine herpesvirus 2. J. Clin. Microbiol. 2002, 40, 3381–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vikoren, T.; Li, H.; Lillehaug, A.; Jonassen, C.M.; Bockerman, I.; Handeland, K. Malignant catarrhal fever in free-ranging cervids associated with OvHV-2 and CpHV-2 DNA. J. Wildl. Dis. 2006, 42, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Huang, Q.; Hu, X.; Chu, W.; Zhang, J.; Jiang, L.; Yu, X.; Zhang, X.; Cheng, S. Caprine herpesvirus 2-associated malignant catarrhal fever of captive sika deer (Cervus nippon) in an intensive management system. BMC Vet. Res. 2018, 14, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Gailbreath, K.; Flach, E.J.; Taus, N.S.; Cooley, J.; Keller, J.; Russell, G.C.; Knowles, D.P.; Haig, D.M.; Oaks, J.L.; et al. A novel subgroup of rhadinoviruses in ruminants. J. Gen. Virol. 2005, 86, 3021–3026. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cunha, C.W.; Taus, N.S.; Knowles, D.P. Malignant catarrhal fever: Inching toward understanding. Annu. Rev. Anim. Biosci. 2014, 2, 209–233. [Google Scholar] [CrossRef] [PubMed]
- Loken, T.; Aleksandersen, M.; Reid, H.; Pow, I. Malignant catarrhal fever caused by ovine herpesvirus-2 in pigs in Norway. Vet. Rec. 1998, 143, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cunha, C.W.; Abbitt, B.; deMaar, T.W.; Lenz, S.D.; Hayes, J.R.; Taus, N.S. Goats are a potential reservoir for the herpesvirus (MCFV-WTD), causing malignant catarrhal fever in deer. J. Zoo Wildl. Med. 2013, 44, 484–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Keller, J.; Knowles, D.P.; Crawford, T.B. Recognition of another member of the malignant catarrhal fever virus group: An endemic gammaherpesvirus in domestic goats. J. Gen. Virol. 2001, 82, 227–232. [Google Scholar] [CrossRef]
- O’Toole, D.; Li, H. The pathology of malignant catarrhal fever, with an emphasis on ovine herpesvirus 2. Vet. Pathol. 2014, 51, 437–452. [Google Scholar] [CrossRef] [Green Version]
- Kekkonen, J.; Wikstrom, M.; Ala-Ajos, I.; Lappalainen, V.; Brommer, J.E. Growth and age structure in an introduced and hunted cervid population: White-tailed deer in Finland. Ann. Zool. Fenn. 2016, 53, 69–80. [Google Scholar] [CrossRef]
- Morellet, N.; Klein, F.; Solberg, E.; Andersen, R. The census and management of populations of ungulates in Europe. In Ungulate Management in Europe: Problems and Practices, 1st ed.; Putman, R., Apollonio, M., Andersen, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 106–143. [Google Scholar]
- Mysterud, A. Diet overlap among ruminants in Fennoscandia. Oecologia 2000, 124, 130–137. [Google Scholar] [CrossRef]
- das Neves, C.G.; Roth, S.; Rimstad, E.; Thiry, E.; Tryland, M. Cervid herpesvirus 2 infection in reindeer: A review. Vet. Microbiol. 2010, 143, 70–80. [Google Scholar] [CrossRef]
- Solberg, M.F.; Strandskog, G.; Veits, J.; Andersen, R.; Heim, M.; Rolandsen, C.M.; Horak, I.G.; Solem, M.I.; Eriksen, R.; Astrup, R. Wild Cervids 2009—Annual Report from the National Monitoring Program for Wild Cervids in Norway; The Norwegian Institute for Nature Research (NINA): Trondheim, Norway, 2010; pp. 1–81. [Google Scholar]
- Austrheim, G.; Solberg, E.J.; Mysterud, A. Spatio-temporal variation in large herbivore pressure in Norway during 1949–1999: Has decreased grazing by livestock been countered by increased browsing by cervids? Wildlife Biol. 2011, 17, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Loken, T.; Bosman, A.M.; van Vuuren, M. Infection with Ovine herpesvirus 2 in Norwegian herds with a history of previous outbreaks of malignant catarrhal fever. J. Vet. Diagn. Investig. 2009, 21, 257–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Westover, W.C.; Crawford, T.B. Sheep-associated malignant catarrhal fever in a petting zoo. J. Zoo Wildl. Med. 1999, 30, 408–412. [Google Scholar] [PubMed]
- Tryland, M.; Mork, T. Gammaherpesvirus–Herpesvirus infections. In Reindeer and Caribou: Health and Disease; Tryland, M., Kutz, S., Eds.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2018; pp. 286–289. [Google Scholar] [CrossRef]
- das Neves, C.G.; Ihlebaek, H.M.; Skjerve, E.; Hemmingsen, W.; Li, H.; Tryland, M. Gammaherpesvirus infection in semidomesticated reindeer (Rangifer tarandus tarandus): A cross-sectional, serologic study in northern norway. J. Wildl. Dis. 2013, 49, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Benestad, S.L.; Mitchell, G.; Simmons, M.; Ytrehus, B.; Vikøren, T. First case of chronic wasting disease in Europe in a Norwegian free-ranging reindeer. Vet. Res. 2016, 47, 88. [Google Scholar] [CrossRef]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, B.; Dural, G.; Yasmum, N.; Lembo, T.; de, T.B.; Ryser-Degiorgis, M.P.; Ulrich, R.G.; McGeoch, D.J. Novel mammalian herpesviruses and lineages within the Gammaherpesvirinae: Cospeciation and interspecies transfer. J. Virol. 2008, 82, 3509–3516. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Dalton, C.S.; van de Rakt, K.; Fahlman, Å.; Ruckstuhl, K.; Neuhaus, P.; Popko, R.; Kutz, S.; van der Meer, F. Discovery of herpesviruses in Canadian wildlife. Arch. Virol. 2017, 162, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Tryland, M.; Sánchez Romano, J.; Nymo, I.H.; Breines, E.M.; Ancin Murguzur, F.J.; Kjenstad, O.C.; Li, H. A screening for virus infections in eight different herds of semi-domesticated Eurasian tundra reindeer (Rangifer tarandus tarandus) in Norway, 2013–2017. Unpublished work.
- Pellet, P.E.; Roizmann, B. Herpesviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1802–1822. [Google Scholar]
- Sánchez Romano, J.; Grund, L.; Obiegala, A.; Nymo, I.H.; Ancin-Murguzur, F.J.; Li, H.; Król, N.; Pfeffer, M.; Tryland, M. A Multi-Pathogen Screening of Captive Reindeer (Rangifer tarandus) in Germany Based on Serological and Molecular Assays. Front. Vet. Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Zarnke, R.L.; Li, H.; Crawford, T.B. Serum antibody prevalence of malignant catarrhal fever viruses in seven wildlife species from Alaska. J. Wildl. Dis. 2002, 38, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Thacker, T.C.; Madison, R.J.; Koster, L.G.; Swenson, S.L.; Li, H. Active and latent ovine herpesvirus-2 (OvHV-2) infection in a herd of captive white-tailed deer (Odocoileus virginianus). J. Comp. Pathol. 2013, 149, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Grinde, B. Herpesviruses: Latency and reactivation—Viral strategies and host response. J. Oral Microbiol. 2013, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vikoren, T.; Klevar, S.; Li, H.; Hauge, A.G. A geographic cluster of malignant catarrhal fever in moose (Alces alces) in Norway. J. Wildl. Dis. 2015, 51, 471–474. [Google Scholar] [CrossRef]
- Waramel, Y.; Steen, M. Malignant catarrhal fever in wild Swedish moose (Alces alces L). Rangifer 1989, 9, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Neimanis, A.S.; Hill, J.E.; Jardine, C.M.; Bollinger, T.K. Sheep-associated malignant catarrhal fever in free-ranging moose (Alces alces) in Saskatchewan, Canada. J. Wildl. Dis. 2009, 45, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Squires, R.A.; Wilson, P.R.; Whelan, N.C.; Johnstone, A.C.; Ayanegui-Alcerreca, M.A.; Castillo-Alcala, F.; Knight, D. Alpha and gamma herpesvirus detection in two herds of farmed red deer (Cervus elaphus) in New Zealand. N. Zeal. Vet. J. 2012, 60, 69–75. [Google Scholar] [CrossRef]
- Pruvot, M.; Kutz, S.; van der Meer, F.; Musiani, M.; Barkema, H.W.; Orsel, K. Pathogens at the livestock-wildlife interface in Western Alberta: Does transmission route matter? Vet. Res. 2014, 45. [Google Scholar] [CrossRef] [Green Version]
- Rzhetsky, A.; Nei, M. A Simple Method for Estimating and Testing Minimum-Evolution Trees. Mol. Biol. Evol. 1992, 9, 945. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Species | Municipality (for Red Deer and Moose) or Reindeer District | Number of Samples | Gammaherpesvirus PCR | |
|---|---|---|---|---|
| DPOL + | gB + | |||
| Semi-domesticated reindeer | 7 ZE Rákkonjárga | 3 | 0 | 0 |
| 13ZG Lágesduottar/Ifjordfjellet | 3 | 1 | 0 | |
| 16 ZS Kárašjoga oarjjabealli Karasjok vestre | 10 | 4 | 2 | |
| 27 YK Joahkonjárga | 3 | 0 | 0 | |
| 33YP Spalca | 1 | 1 | 1 | |
| 34 YR Ábborašša | 3 | 1 | 0 | |
| 36 YT Cohkolat ja Biertavárri | 1 | 0 | 0 | |
| 40 YX Orda | 15 | 7 | 1 | |
| subtotal | 39 | 14 | 4 | |
| Wild reindeer | 1 Setesdal Ryfylke | 4 | 2 | 1 |
| 7 Hardangervidda | 13 | 9 | 8 | |
| 11 Nordfjella | 10 | 6 | 6 | |
| 17 Reinheimen-Breheimen | 1 | 0 | 0 | |
| 18 Snøhetta | 1 | 0 | 0 | |
| 19 Rondane sør | 2 | 0 | 0 | |
| 23 Knutshø | 4 | 0 | 0 | |
| subtotal | 35 | 17 | 15 | |
| Red deer | Aurland | 4 | 0 | 0 |
| Hol | 6 | 0 | 0 | |
| Kvinnherad | 11 | 0 | 0 | |
| Lærdal | 16 | 0 | 0 | |
| Ørstad | 4 | 0 | 0 | |
| subtotal | 41 | 0 | 0 | |
| Moose | Selbu | 24 | 0 | 0 |
| Vega | 27 | 0 | 0 | |
| subtotal | 51 | 0 | 0 | |
| Total | 166 | 31 | 19 | |
| cELISA | ||||
|---|---|---|---|---|
| POS | NEG | TOTALS | ||
| PCR Results | DPOL +/gB + | 4 | 0 | 4 |
| DPOL +/gB − | 7 | 3 | 10 | |
| DPOL −/gB + | 0 | 0 | 0 | |
| DPOL −/gB − | 12 | 13 | 25 | |
| TOTALS | 23 | 16 | 39 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
das Neves, C.G.; Sacristán, C.; Madslien, K.; Tryland, M. Gammaherpesvirus in Cervid Species from Norway: Characterization of a New Virus in Wild and Semi-Domesticated Eurasian Tundra Reindeer (Rangifer tarandus tarandus). Viruses 2020, 12, 876. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080876
das Neves CG, Sacristán C, Madslien K, Tryland M. Gammaherpesvirus in Cervid Species from Norway: Characterization of a New Virus in Wild and Semi-Domesticated Eurasian Tundra Reindeer (Rangifer tarandus tarandus). Viruses. 2020; 12(8):876. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080876
Chicago/Turabian Styledas Neves, Carlos G., Carlos Sacristán, Knut Madslien, and Morten Tryland. 2020. "Gammaherpesvirus in Cervid Species from Norway: Characterization of a New Virus in Wild and Semi-Domesticated Eurasian Tundra Reindeer (Rangifer tarandus tarandus)" Viruses 12, no. 8: 876. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080876