Combinatorial Treatment of Birinapant and Zosuquidar Enhances Effective Control of HBV Replication In Vivo

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Tissue Culture and Viability Assays
2.3. In Vivo Model Studies of Hepatitis B Virus (HBV)
2.4. Preparation of Mouse Livers for Western Blotting
2.5. Western Blot Protein Analysis
2.6. HBcAg Immunohistochemistry
2.7. Statistical Analysis
3. Results
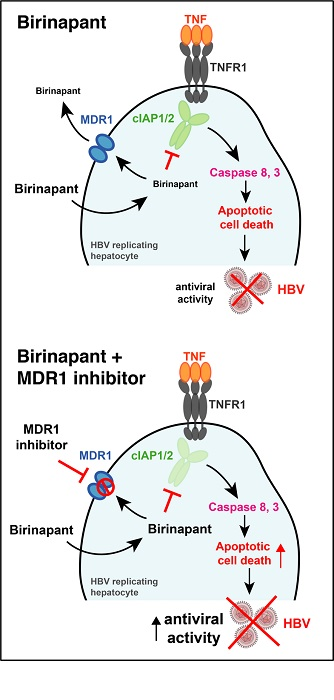
3.1. MDR1 Inhibition Enhances Birinapant-Mediated Killing of HepG2 Cells
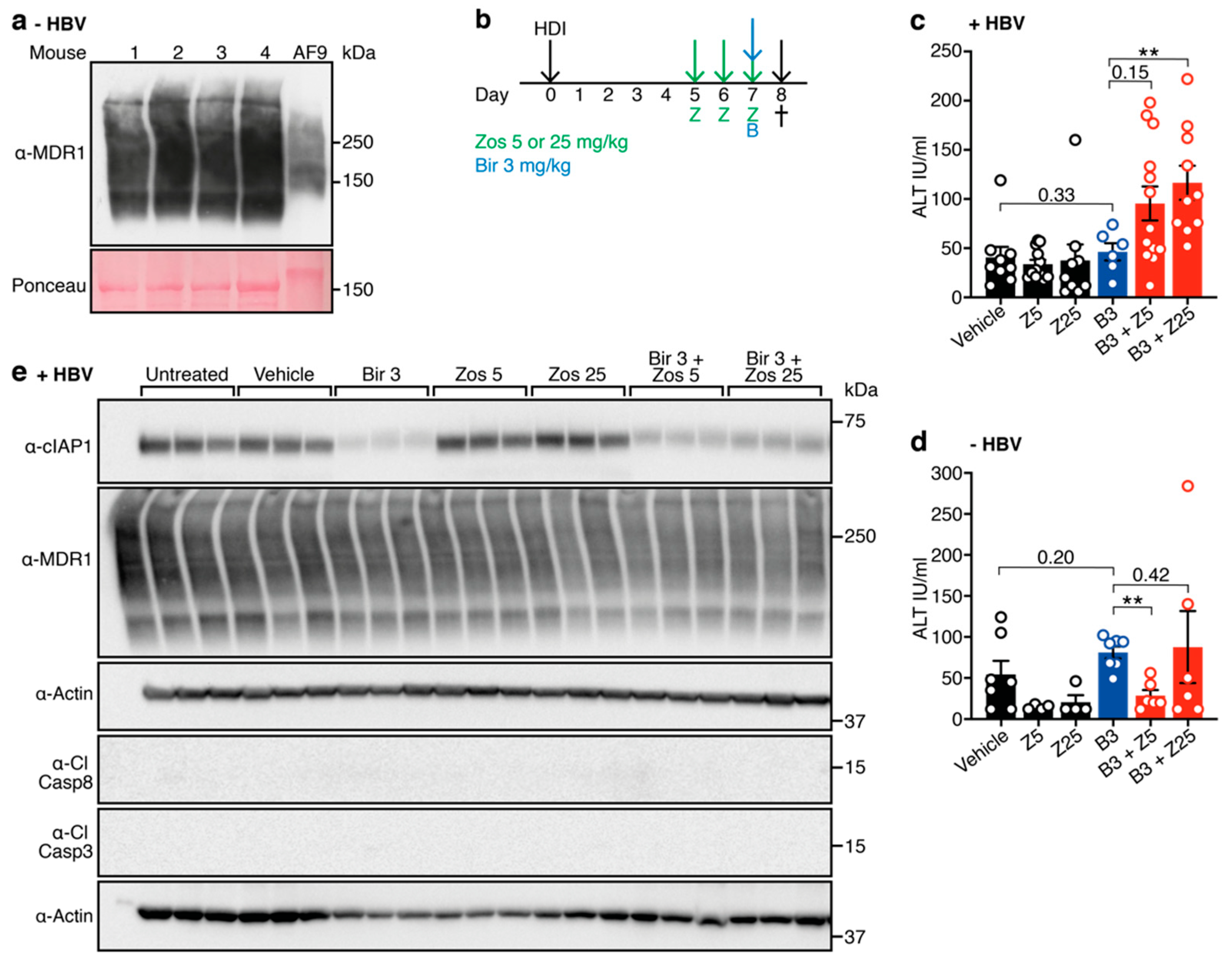
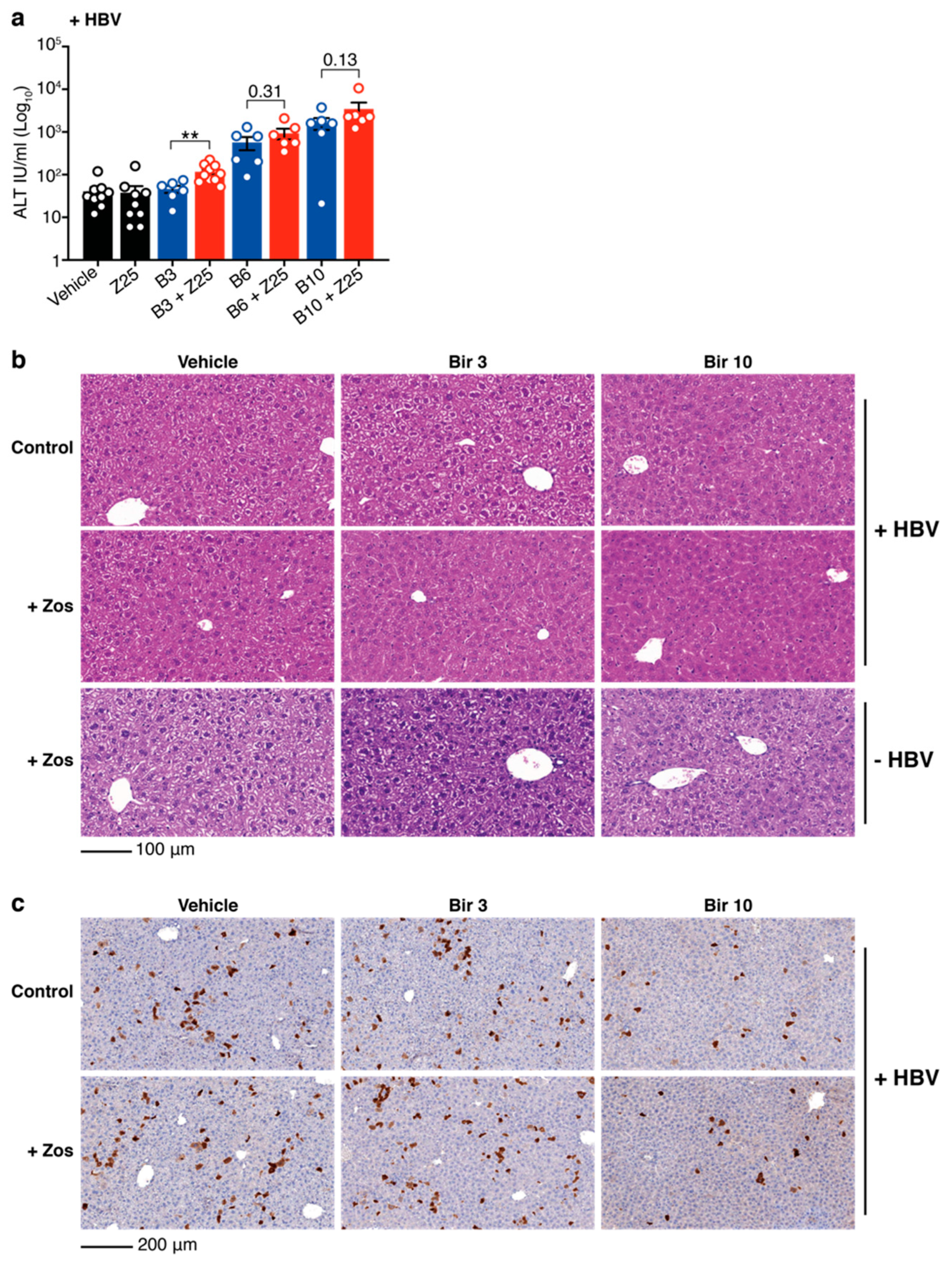
3.2. Combination Treatment with Birinapant and Zosuquidar Is Safe and Increases Death of Hepatocytes in the Liver of HBV-Replicating Mice
3.3. Birinapant and Zosuquidar Combination Therapy Specifically Kills HBV Expresisng Cells in the Liver of Mice
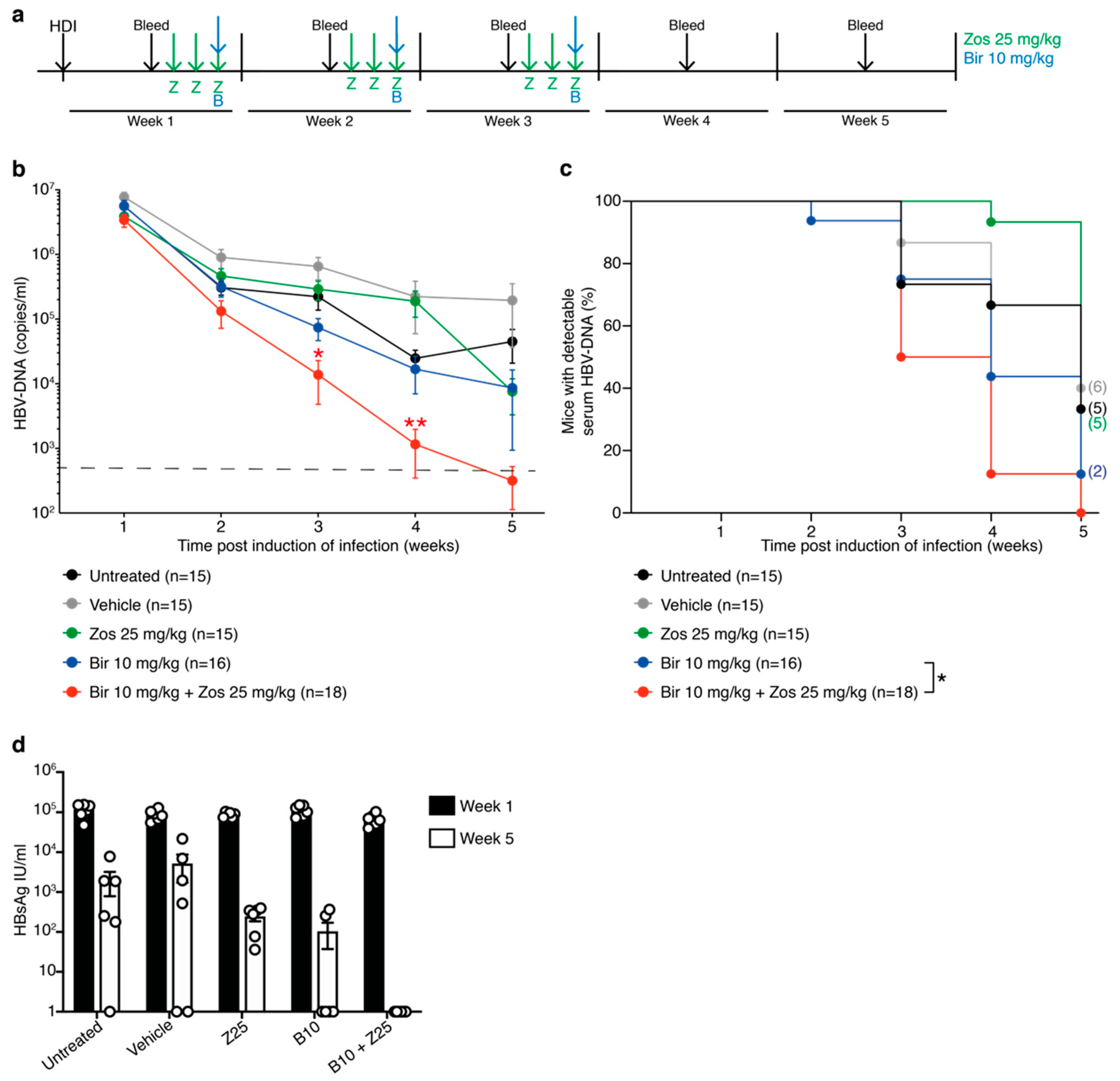
3.4. The MDR1 Inhibitor Zosuquidar Enhances the Efficacy of Birinapant to Diminish HBV Replication In Vivo
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Herbein, G.; O’Brien, W.A. Tumor necrosis factor (TNF)-alpha and TNF receptors in viral pathogenesis. Proc. Soc. Exp. Biol. Med. 2000, 223, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Ebert, G.; Preston, S.; Allison, C.; Cooney, J.; Toe, J.G.; Stutz, M.D.; Ojaimi, S.; Scott, H.W.; Baschuk, N.; Nachbur, U.; et al. Cellular inhibitor of apoptosis proteins prevent clearance of hepatitis B virus. Proc. Natl. Acad. Sci. USA 2015, 112, 5797–5802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almagro, M.C.; Vucic, D. The inhibitor of apoptosis (IAP) proteins are critical regulators of signaling pathways and targets for anti-cancer therapy. Exp. Oncol. 2012, 34, 200–211. [Google Scholar] [PubMed]
- Silke, J. The regulation of TNF signalling: What a tangled web we weave. Curr. Opin. Immunol. 2011, 23, 620–626. [Google Scholar] [CrossRef]
- Morrish, E.; Brumatti, G.; Silke, J. Future Therapeutic Directions for Smac-Mimetics. Cells 2020, 9, 406. [Google Scholar] [CrossRef] [Green Version]
- Richmond, J.; Robbins, A.; Evans, K.; Beck, D.; Kurmasheva, R.T.; Billups, C.A.; Carol, H.; Heatley, S.; Sutton, R.; Marshall, G.M.; et al. Acute Sensitivity of Ph-like Acute Lymphoblastic Leukemia to the SMAC-Mimetic Birinapant. Cancer Res. 2016, 76, 4579–4591. [Google Scholar] [CrossRef] [Green Version]
- Brumatti, G.; Ma, C.; Lalaoui, N.; Nguyen, N.-Y.; Navarro, M.; Tanzer, M.C.; Richmond, J.; Ghisi, M.; Salmon, J.M.; Silke, N.; et al. The caspase-8 inhibitor emricasan combines with the SMAC mimetic birinapant to induce necroptosis and treat acute myeloid leukemia. Sci. Transl. Med. 2016, 8, 339ra69. [Google Scholar] [CrossRef] [Green Version]
- Lalaoui, N.; Hänggi, K.; Brumatti, G.; Chau, D.; Nguyen, N.-Y.N.; Vasilikos, L.; Spilgies, L.M.; Heckmann, D.A.; Ma, C.; Ghisi, M.; et al. Targeting p38 or MK2 Enhances the Anti-Leukemic Activity of Smac-Mimetics. Cancer Cell 2016, 29, 145–158. [Google Scholar] [CrossRef] [Green Version]
- McComb, S.; Aguadé-Gorgorió, J.; Harder, L.; Marovca, B.; Cario, G.; Eckert, C.; Schrappe, M.; Stanulla, M.; von Stackelberg, A.; Bourquin, J.-P.; et al. Activation of concurrent apoptosis and necroptosis by SMAC mimetics for the treatment of refractory and relapsed ALL. Sci. Transl. Med. 2016, 8, 339ra70. [Google Scholar] [CrossRef]
- Vince, J.E.; Wong, W.W.-L.; Khan, N.; Feltham, R.; Chau, D.; Ahmed, A.U.; Benetatos, C.A.; Chunduru, S.K.; Condon, S.M.; McKinlay, M.; et al. IAP antagonists target cIAP1 to induce TNFalpha-dependent apoptosis. Cell 2007, 131, 682–693. [Google Scholar] [CrossRef] [Green Version]
- Varfolomeev, E.; Blankenship, J.W.; Wayson, S.M.; Fedorova, A.V.; Kayagaki, N.; Garg, P.; Zobel, K.; Dynek, J.N.; Elliott, L.O.; Wallweber, H.J.; et al. IAP antagonists induce autoubiquitination of c-IAPs, NF-kappaB activation, and TNFalpha-dependent apoptosis. Cell 2007, 131, 669–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaither, A.; Porter, D.; Yao, Y.; Borawski, J.; Yang, G.; Donovan, J.; Sage, D.; Slisz, J.; Tran, M.; Straub, C.; et al. A Smac mimetic rescue screen reveals roles for inhibitor of apoptosis proteins in tumor necrosis factor-alpha signaling. Cancer Res. 2007, 67, 11493–11498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, G.; Allison, C.; Preston, S.; Cooney, J.; Toe, J.G.; Stutz, M.D.; Ojaimi, S.; Baschuk, N.; Nachbur, U.; Torresi, J.; et al. Eliminating hepatitis B by antagonizing cellular inhibitors of apoptosis. Proc. Natl. Acad. Sci. USA 2015, 112, 5803–5808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Hou, J.; Zhang, X. Targeting cIAPs, a New Option for Functional Cure of Chronic Hepatitis B Infection? Virol. Sin. 2018, 33, 459–461. [Google Scholar] [CrossRef]
- Ebert, G.; Lopaticki, S.; O’Neill, M.T.; Steel, R.W.J.; Doerflinger, M.; Rajasekaran, P.; Yang, A.S.P.; Erickson, S.; Ioannidis, L.; Arandjelovic, P.; et al. Targeting the Extrinsic Pathway of Hepatocyte Apoptosis Promotes Clearance of Plasmodium Liver Infection. Cell Rep. 2020, 30, 4343–4354.e4. [Google Scholar] [CrossRef]
- Morrish, E.; Copeland, A.; Moujalled, D.M.; Powell, J.A.; Silke, S.; Lin, A.; Jarman, K.E.; Sandow, J.J.; Ebert, G.; Mackiewicz, L.; et al. Clinical MDR1 inhibitors enhance Smac-mimetic bioavailability to kill LSCs and improve survival of AML models. Blood Adv. 2020. under minor revision. [Google Scholar]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef]
- Starling, J.J.; Shepard, R.L.; Cao, J.; Law, K.L.; Norman, B.H.; Kroin, J.S.; Ehlhardt, W.J.; Baughman, T.M.; Winter, M.A.; Bell, M.G.; et al. Pharmacological characterization of LY335979: A potent cyclopropyldibenzosuberane modulator of P-glycoprotein. Adv. Enzyme Regul. 1997, 37, 335–347. [Google Scholar] [CrossRef]
- Ros, J.E.; Libbrecht, L.; Geuken, M.; Jansen, P.L.M.; Roskams, T.A.D. High expression of MDR1, MRP1, and MRP3 in the hepatic progenitor cell compartment and hepatocytes in severe human liver disease. J. Pathol. 2003, 200, 553–560. [Google Scholar] [CrossRef]
- Bennukul, K.; Numkliang, S.; Leardkamolkarn, V. Melatonin attenuates cisplatin-induced HepG2 cell death via the regulation of mTOR and ERCC1 expressions. World J. Hepatol 2014, 6, 230–242. [Google Scholar] [CrossRef]
- Huang, L.-R.; Wu, H.-L.; Chen, P.-J.; Chen, D.-S. An immunocompetent mouse model for the tolerance of human chronic hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2006, 103, 17862–17867. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.L.; Althage, A.; Chung, J.; Chisari, F.V. Hydrodynamic injection of viral DNA: A mouse model of acute hepatitis B virus infection. Proc. Natl. Acad. Sci. USA 2002, 99, 13825–13830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wang, C.; Tang, S.; Sun, Y.; Zhao, D.; Zhang, S.; Deng, S.; Zhou, Y.; Xiao, X. TNFR1/TNF-α and mitochondria interrelated signaling pathway mediates quinocetone-induced apoptosis in HepG2 cells. Food Chem. Toxicol. 2013, 62, 825–838. [Google Scholar] [CrossRef]
- World Health Organization WHO. Global Hepatitis Report 2017. Available online: https://www.who.int/hepatitis/publications/global-hepatitis-report2017/en/ (accessed on 30 April 2017).
- Ganem, D.; Prince, A.M. Hepatitis B virus infection--natural history and clinical consequences. N. Engl. J. Med. 2004, 350, 1118–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, H.; Zhong, G.; Xu, G.; He, W.; Jing, Z.; Gao, Z.; Huang, Y.; Qi, Y.; Peng, B.; Wang, H.; et al. Sodium taurocholate cotransporting polypeptide is a functional receptor for human hepatitis B and D virus. eLife 2012, 1, e00049. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, T.; Jost, P.J.; Pellegrini, M.; Puthalakath, H.; Gugasyan, R.; Gerondakis, S.; Cretney, E.; Smyth, M.J.; Silke, J.; Hakem, R.; et al. Fatal hepatitis mediated by tumor necrosis factor TNFalpha requires caspase-8 and involves the BH3-only proteins Bid and Bim. Immunity 2009, 30, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crook, N.E.; Clem, R.J.; Miller, L.K. An apoptosis-inhibiting baculovirus gene with a zinc finger-like motif. J. Virol. 1993, 67, 2168–2174. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrish, E.; Mackiewicz, L.; Silke, N.; Pellegrini, M.; Silke, J.; Brumatti, G.; Ebert, G. Combinatorial Treatment of Birinapant and Zosuquidar Enhances Effective Control of HBV Replication In Vivo. Viruses 2020, 12, 901. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080901
Morrish E, Mackiewicz L, Silke N, Pellegrini M, Silke J, Brumatti G, Ebert G. Combinatorial Treatment of Birinapant and Zosuquidar Enhances Effective Control of HBV Replication In Vivo. Viruses. 2020; 12(8):901. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080901
Chicago/Turabian StyleMorrish, Emma, Liana Mackiewicz, Natasha Silke, Marc Pellegrini, John Silke, Gabriela Brumatti, and Gregor Ebert. 2020. "Combinatorial Treatment of Birinapant and Zosuquidar Enhances Effective Control of HBV Replication In Vivo" Viruses 12, no. 8: 901. https://0-doi-org.brum.beds.ac.uk/10.3390/v12080901