Identification of Inhibitors of ZIKV Replication

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Lines
2.2. ZIKV Strains
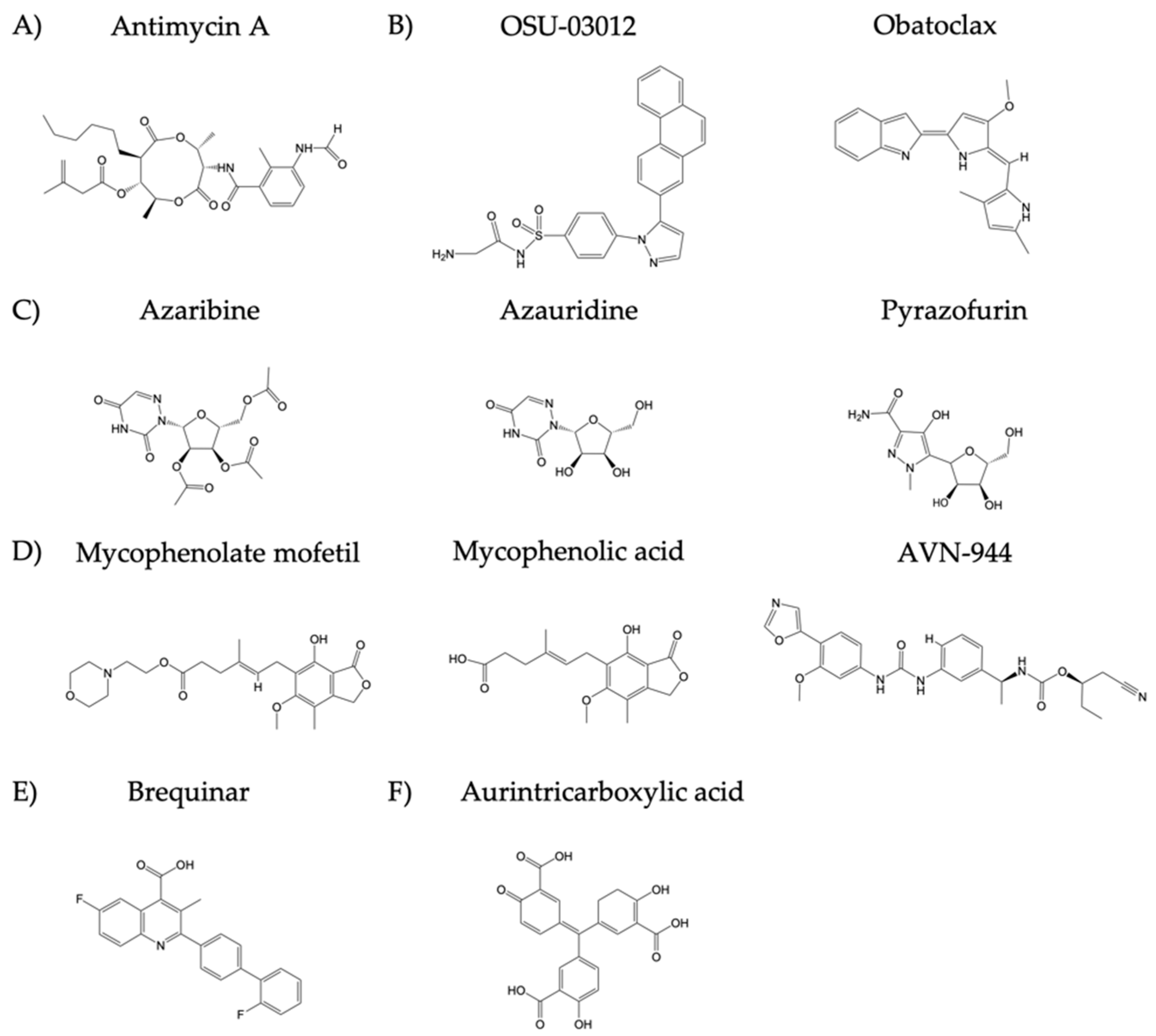
2.3. Compounds
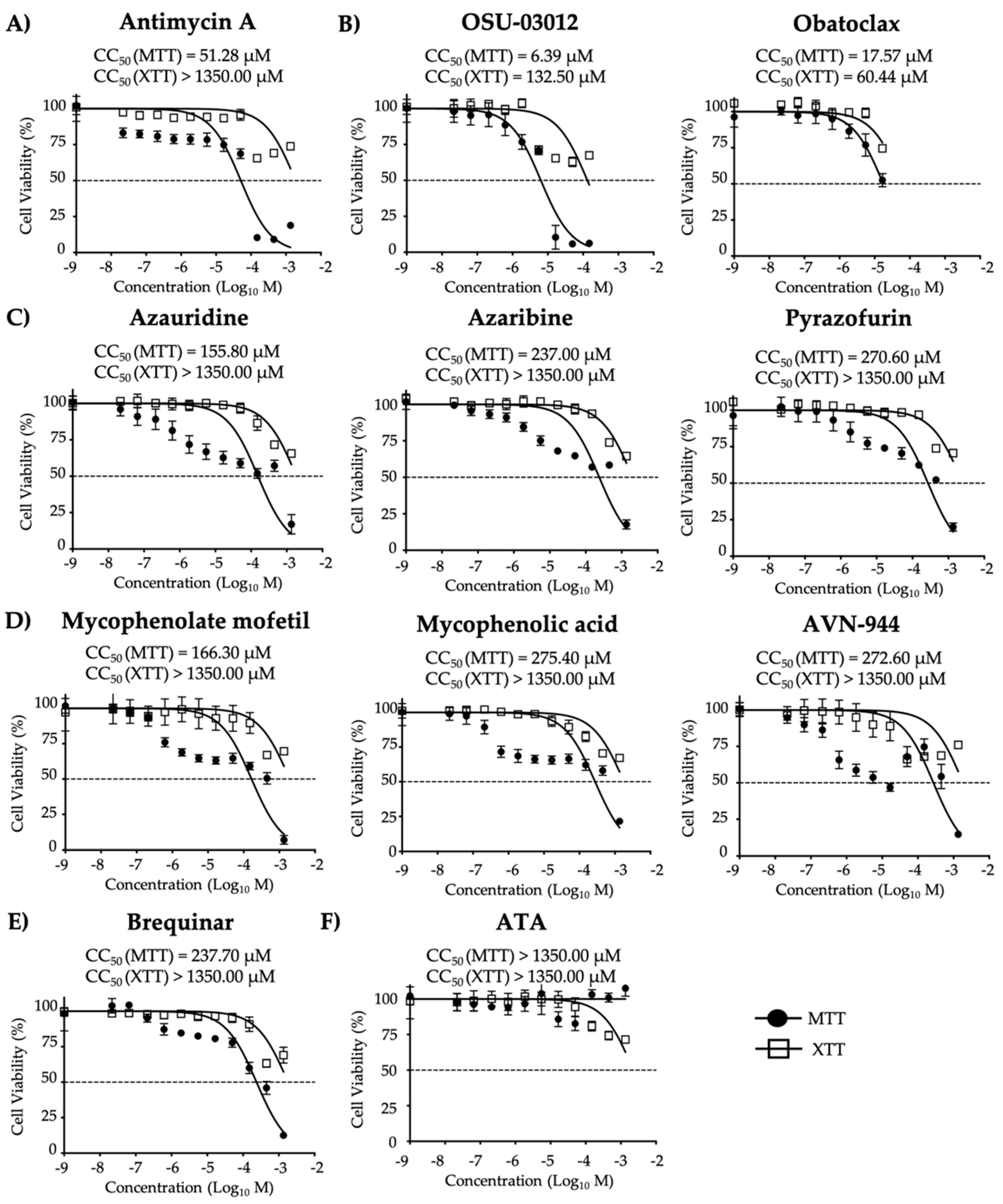
2.4. Cell Viability Assays
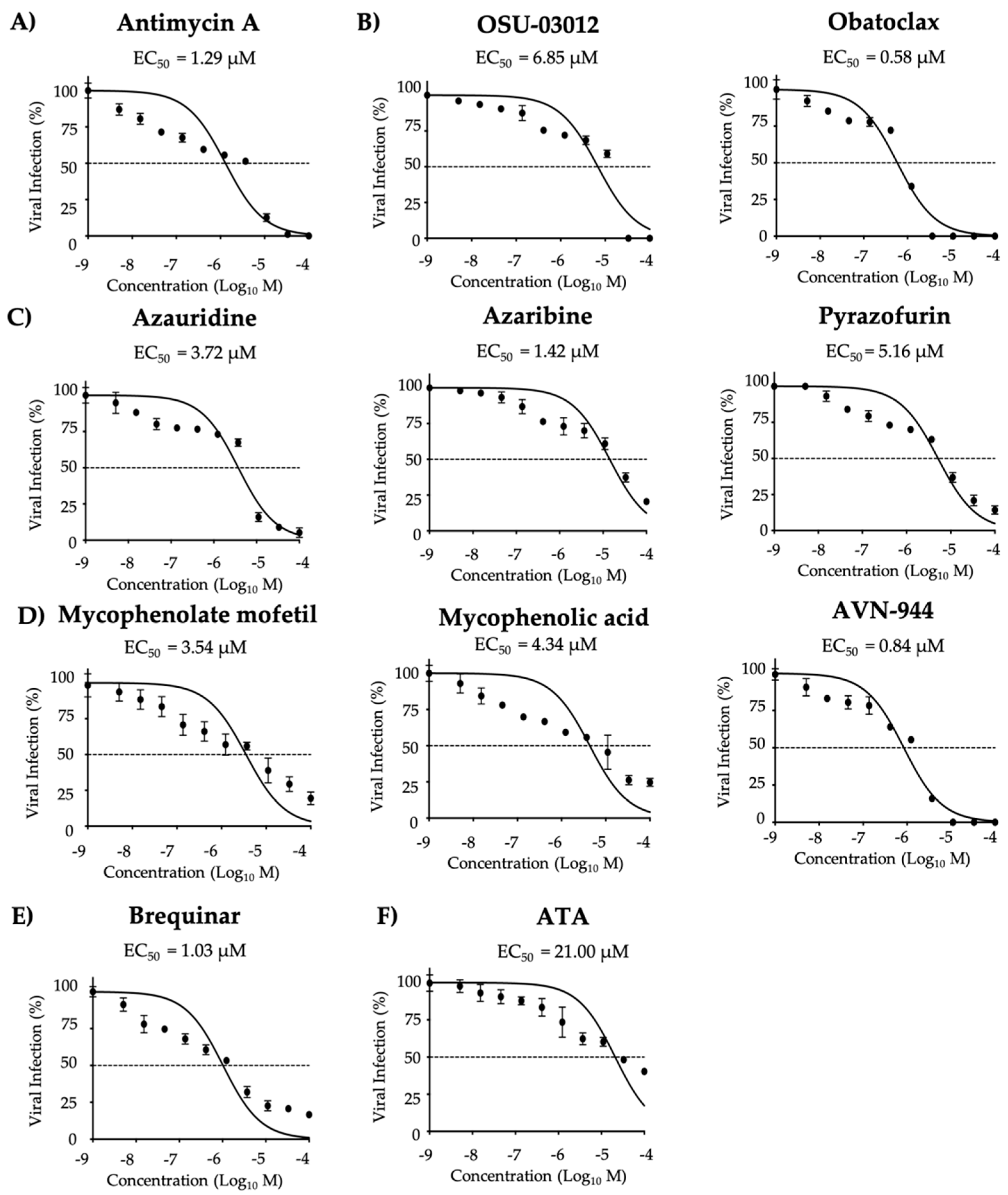
2.5. Viral Inhibition Assays
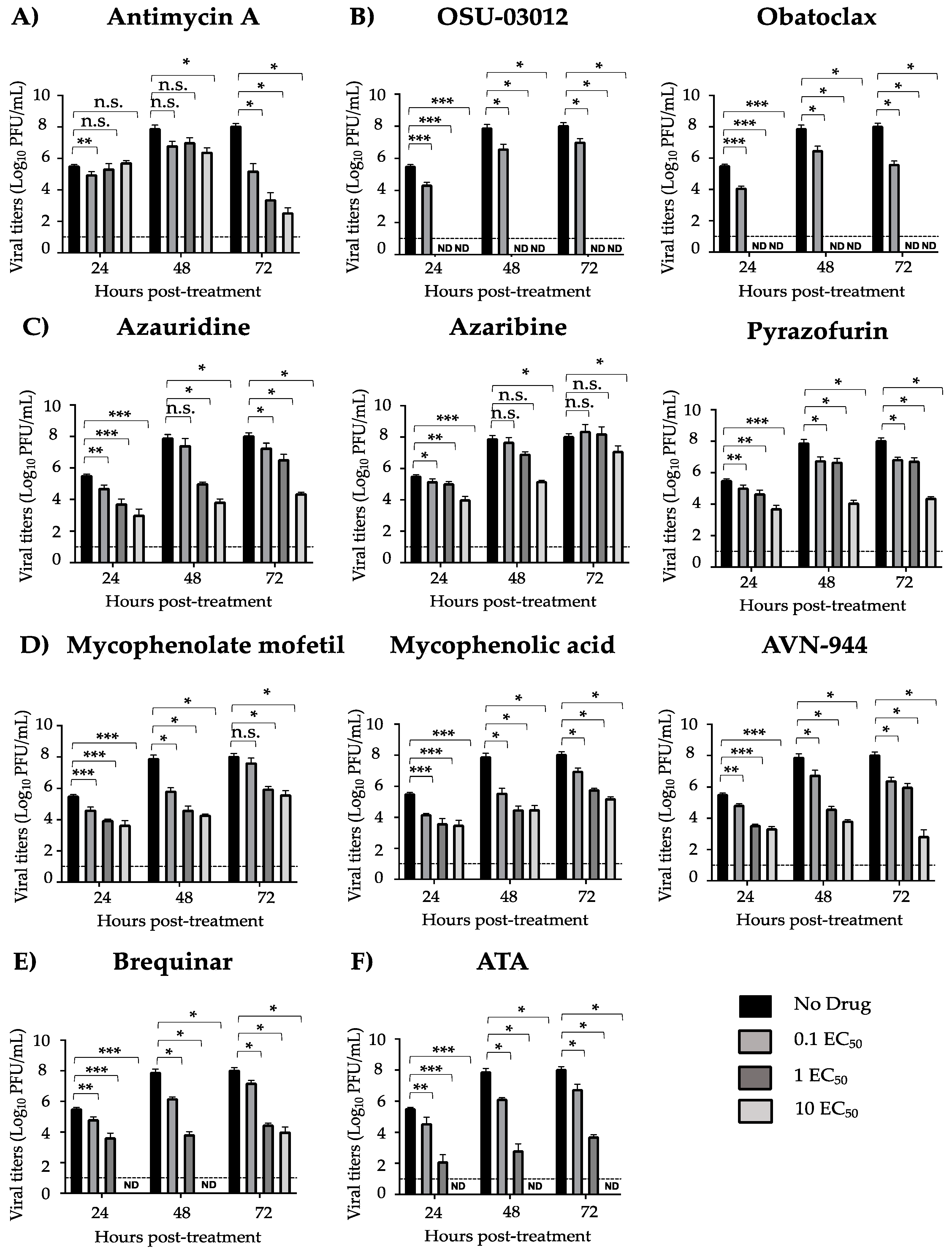
2.6. Growth Inhibition Assays
2.7. Protection Against ZIKV-Induced Cytopathic Effect (CPE)
2.8. Apoptosis Assay
2.9. Statistical Analysis
3. Results
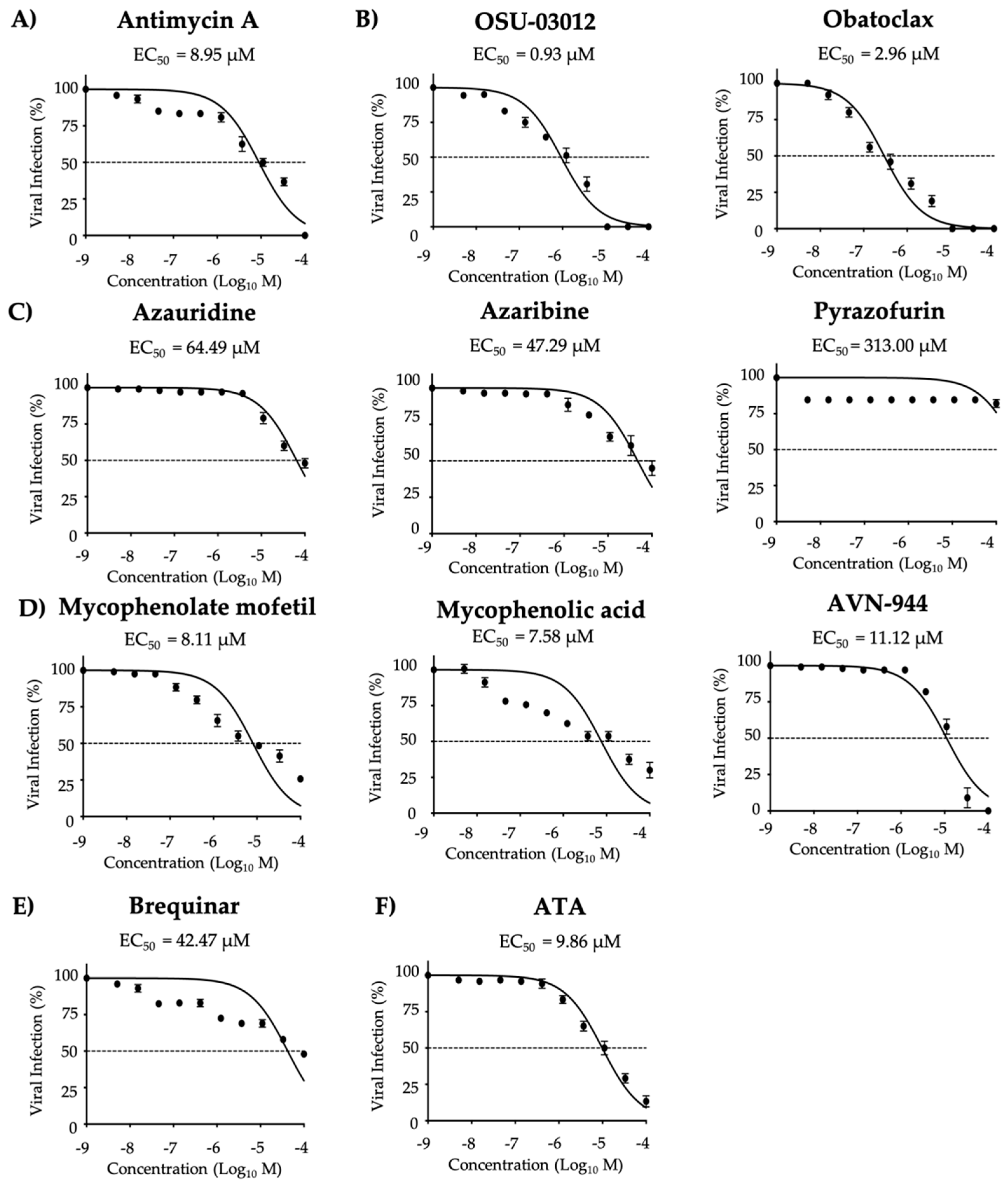
3.1. Antiviral Activity of Compounds Against ZIKV
3.2. Pre- and Co-Treatment Effect of Compounds on ZIKV Infection
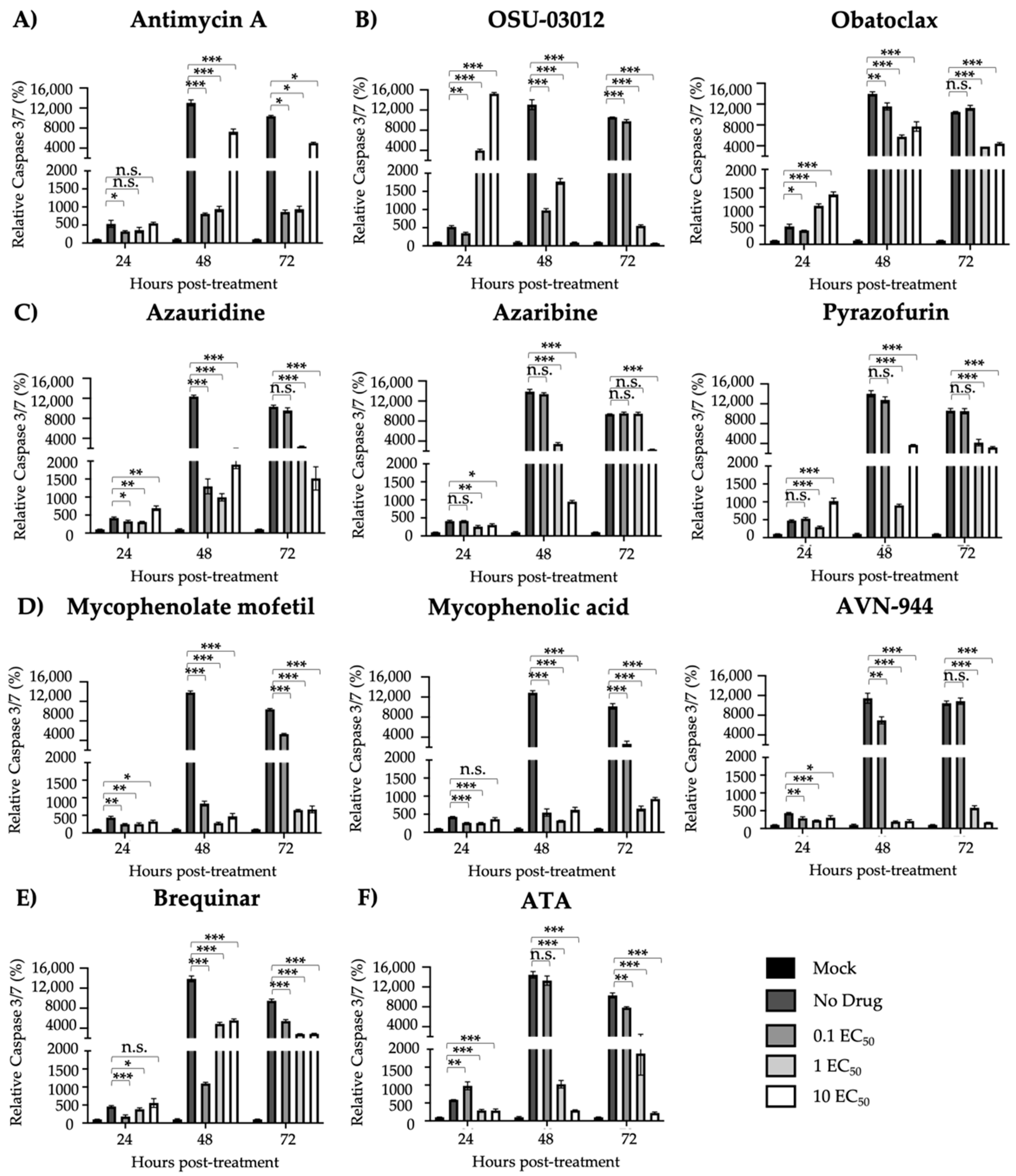
3.3. Protection from ZIKV-Induced Cell Death
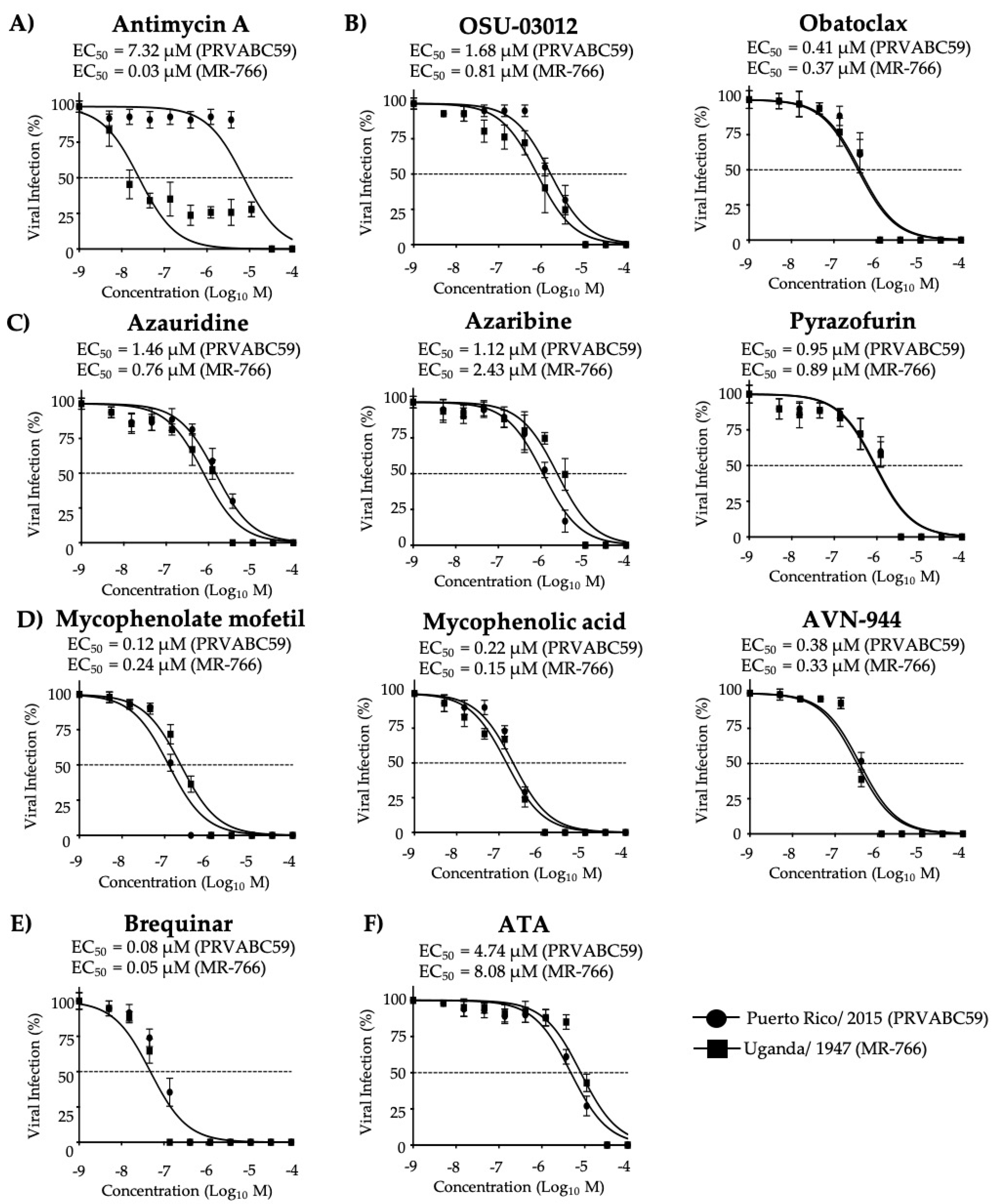
3.4. Effect of the Compounds on Old World and New World ZIKV Strains
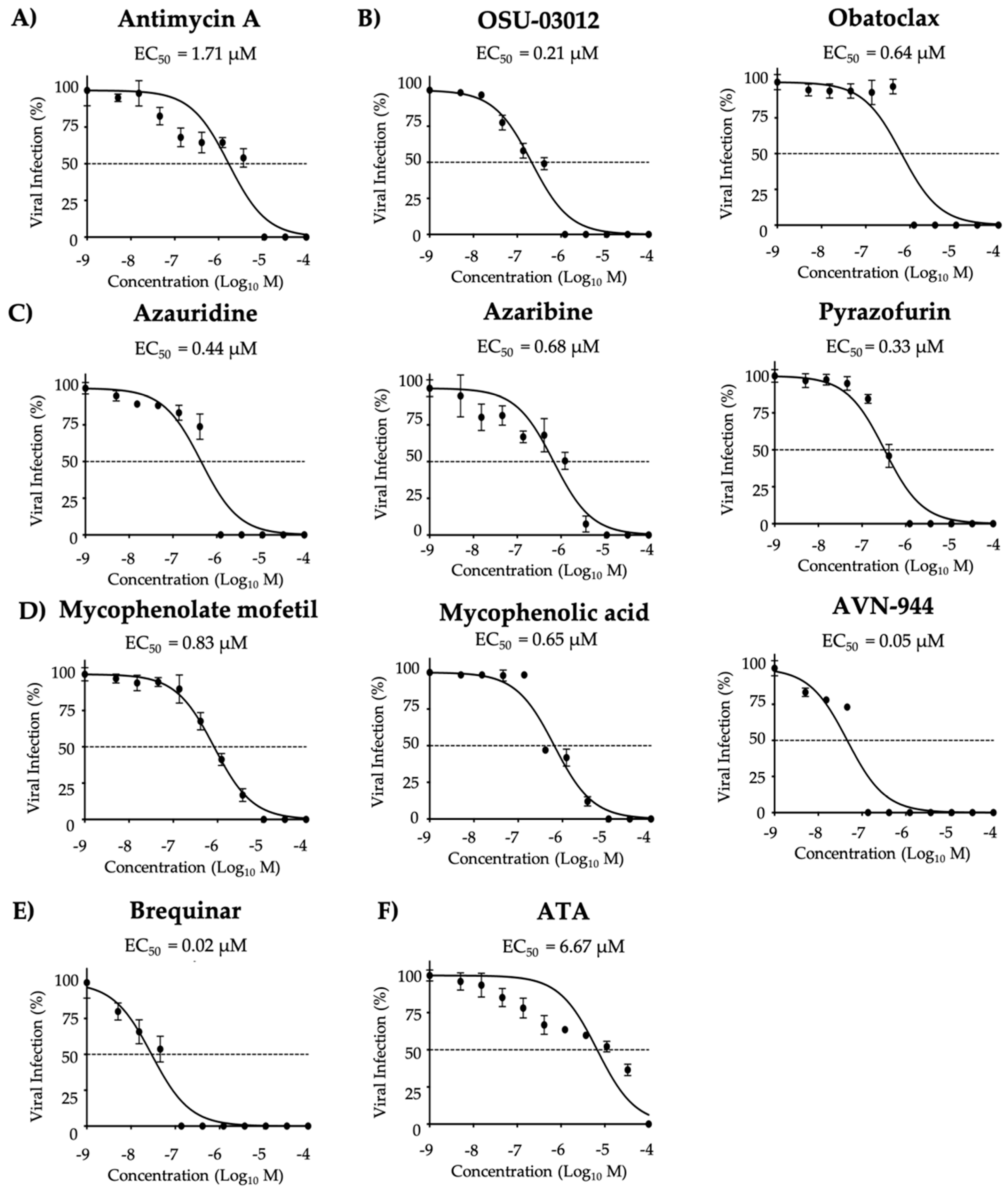
3.5. Effect of the Compounds on ZIKV Infection in Human A549 Cells
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Compound | Literature Findings | References | |
|---|---|---|---|---|
| A | Antimycin A | Viruses | Togaviridae (Venezuelan equine encephalitis virus, VEEV), Bunyaviridae (La Crosse virus, LACV), Picornaviridae (encephalomyocarditis virus, EMCV) Rhabdoviridae (vesicular stomatitis virus, VSV), Paramyxoviridae (Sendai virus, SeV), Flaviviridae (hepatitis C virus, HCV), Orthomyxoviridae (influenza A and B viruses) families | [17,21] |
| B | OSU-03012 | Bacteria | Salmonella enterica and Francisella tularensis | [22,23] |
| Fungi | Candida albicans, Cryptococcus neoformans, Fusarium sp., mucorales, Blastomyces dermatitidis, Histoplasma capsulatum, and Coccidioides immitis | |||
| Parasite | Leishmania donovani | |||
| Viruses | Lassa virus, LASV; Marburg virus, MARV; Ebola virus, EBOV; DENV; Junin virus, JUNV; rubella virus, RV; and human immunodeficiency virus, HIV | |||
| Obatoclax | Viruses | Influenza A and B viruses, Bunyamwera virus (BUNV), and Sindbis virus (SINV), Chikungunya virus (CHIKV) and Semliki Forest virus (SFV) | [25,26] | |
| C | Azaribine | Viruses | WNV and influenza A and B viruses | [17,28,29] |
| Azauridine | Viruses | CHIKV, SFV, YFV, DENV, JEV, LCMV, parainfluenza 3 virus (HPIV-3), polyomavirus (PV), influenza A and B viruses, and WNV | [16,17,29,31,32,33] | |
| Pyrazofurin | Viruses | Rhinovirus, Herpes simplex virus (HSV), vaccinia virus, WNV, Picornaviridae (polio and Coxsackie B4), SINV, YFV, Paramyxoviridae (measles virus, MV; and respiratory syncytial virus, RSV), Orthomyxoviridae (influenza A and B viruses), arenavirus (JUNV and Tacaribe virus, TCRV), and Rhabdoviridae (VSV) | [17,29,35,36] | |
| D | Mycophenolate mofetil | Viruses | Influenza A and B viruses, foot-and-mouth disease virus (FMDV), and HPIV-2 | [17,37,39,40] |
| Mycophenolic acid | Viruses | Influenza A and B viruses, camelpox virus, cowpox virus, monkeypox virus, vaccinia virus, reovirus, and HCV, among | [17,43,44,45,46,47] | |
| AVN-944 | Viruses | FDMV, influenza A and B viruses, and JUNV | [17,37,50,51]. | |
| E | Brequinar | Viruses | FMDV, HIV, DENV, WNV, YFV, Powassan virus (POW), Western equine encephalitis virus (WEEV), VSV, rotavirus, EBOV, and Cantagalo virus | [52,53,54,55,56,57] |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacKenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging flaviviruses: The spread and resurgence of Japanese encephalitis, West Nile and dengue viruses. Nat. Med. 2004, 10, S98–S109. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Pérez, G.; Nogales, A.; Martín, V.; Almazán, F.; Martínez-Sobrido, L. Reverse Genetic Approaches for the Generation of Recombinant Zika Virus. Viruses 2018, 10, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J.; Zika Virus, I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Ioos, S.; Mallet, H.-P.; Goffart, I.L.; Gauthier, V.; Cardoso, T.; Herida, M. Current Zika virus epidemiology and recent epidemics. Med. Mal. Infect. 2014, 44, 302–307. [Google Scholar] [CrossRef]
- Duffy, M.R.; Chen, T.-H.; Hancock, W.T.; Powers, A.M.; Kool, J.L.; Lanciotti, R.S.; Pretrick, M.; Marfel, M.; Holzbauer, S.; DuBray, C.; et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. N. Engl. J. Med. 2009, 360, 2536–2543. [Google Scholar] [CrossRef]
- Cao-Lormeau, V.-M.; Blake, A.; Mons, S.; Lastère, S.; Roche, C.; Vanhomwegen, J.; Dub, T.; Baudouin, L.; Teissier, A.; Larre, P.; et al. Guillain-Barré Syndrome outbreak associated with Zika virus infection in French Polynesia: A case-control study. Lancet 2016, 387, 1531–1539. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C.; Costa, F.; Garcia-Blanco, M.A.; Ko, A.I.; Ribeiro, G.S.; Saade, G.; Shi, P.-Y.; Vasilakis, N. Zika virus: History, emergence, biology, and prospects for control. Antivir. Res. 2016, 130, 69–80. [Google Scholar] [CrossRef]
- Baz, M.; Boivin, G. Baz Antiviral Agents in Development for Zika Virus Infections. Pharmaceuticals 2019, 12, 101. [Google Scholar] [CrossRef] [Green Version]
- Shuaib, W.; Stanazai, H.; Abazid, A.G.; Mattar, A.A. Re-Emergence of Zika Virus: A Review on Pathogenesis, Clinical Manifestations, Diagnosis, Treatment, and Prevention. Am. J. Med. 2016, 129, 879.e7–879.e12. [Google Scholar] [CrossRef] [Green Version]
- Gunawardana, S.A.; Shaw, R.H. Cross-reactive dengue virus-derived monoclonal antibodies to Zika virus envelope protein: Panacea or Pandora’s box? BMC Infect. Dis. 2018, 18, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbink, P.; Stephenson, K.E.; Barouch, D.H. Zika virus vaccines. Nat. Rev. Genet. 2018, 16, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Garg, H.; Mehmetoglu-Gurbuz, T.; Joshi, A. Recent Advances in Zika Virus Vaccines. Viruses 2018, 10, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McArthur, M.A. Zika Virus: Recent Advances towards the Development of Vaccines and Therapeutics. Viruses 2017, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-G.; Ávila-Pérez, G.; Madere, F.; Hilimire, T.A.; Nogales, A.; Almazán, F.; Martínez-Sobrido, L. Potent Inhibition of Zika Virus Replication by Aurintricarboxylic Acid. Front. Microbiol. 2019, 10, 718. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Cubitt, B.; Chen, E.; Hull, M.V.; Chatterjee, A.K.; Cai, Y.; Kuhn, J.H.; De La Torre, J.C. The ReFRAME library as a comprehensive drug repurposing library to identify mammarenavirus inhibitors. Antivir. Res. 2019, 169, 104558. [Google Scholar] [CrossRef]
- Park, J.-G.; Ávila-Pérez, G.; Nogales, A.; Blanco-Lobo, P.; De La Torre, J.C.; Martínez-Sobrido, L. Identification and Characterization of Novel Compounds with Broad-Spectrum Antiviral Activity against Influenza A and B Viruses. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [Green Version]
- Ávila-Pérez, G.; Nogales, A.; Park, J.-G.; Márquez-Jurado, S.; Iborra, F.J.; Almazán, F.; Martínez-Sobrido, L. A natural polymorphism in Zika virus NS2A protein responsible of virulence in mice. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef]
- Márquez-Jurado, S.; Nogales, A.; Ávila-Pérez, G.; Iborra, F.J.; Martínez-Sobrido, L.; Almazán, F. An Alanine-to-Valine Substitution in the Residue 175 of Zika Virus NS2A Protein Affects Viral RNA Synthesis and Attenuates the Virus In Vivo. Viruses 2018, 10, 547. [Google Scholar] [CrossRef] [Green Version]
- Labs, M.; Rühle, T.; Leister, D. The antimycin A-sensitive pathway of cyclic electron flow: From 1963 to 2015. Photosynth. Res. 2016, 129, 231–238. [Google Scholar] [CrossRef]
- Raveh, A.; Delekta, P.C.; Dobry, C.J.; Peng, W.; Schultz, P.J.; Blakely, P.K.; Tai, A.W.; Matainaho, T.; Irani, D.N.; Sherman, D.H.; et al. Discovery of Potent Broad Spectrum Antivirals Derived from Marine Actinobacteria. PLoS ONE 2013, 8, e82318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.F.; Zhu, Z.; Chu, H.; Yuan, S.; Chik, K.K.-H.; Chan, C.C.-S.; Poon, V.K.-M.; Yip, C.C.-Y.; Zhang, X.; Tsang, J.O.-L.; et al. The celecoxib derivative kinase inhibitor AR-12 (OSU-03012) inhibits Zika virus via down-regulation of the PI3K/Akt pathway and protects Zika virus-infected A129 mice: A host-targeting treatment strategy. Antivir. Res. 2018, 160, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.; Roberts, J.L.; Ecroyd, H.; Tritsch, S.R.; Bavari, S.; Reid, S.P.; Proniuk, S.; Zukiwski, A.; Jacob, A.; Sepúlveda, C.S.; et al. AR-12 Inhibits Multiple Chaperones Concomitant With Stimulating Autophagosome Formation Collectively Preventing Virus Replication. J. Cell. Physiol. 2016, 231, 2286–2302. [Google Scholar] [CrossRef] [PubMed]
- Rausch, K.; Hackett, B.A.; Weinbren, N.L.; Reeder, S.; Sadovsky, Y.; Hunter, C.A.; Schultz, D.C.; Coyne, C.B.; Cherry, S. Screening Bioactives Reveals Nanchangmycin as a Broad Spectrum Antiviral Active against Zika Virus. Cell Rep. 2017, 18, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Kuivanen, S.; Bespalov, M.M.; Nandania, J.; Ianevski, A.; Velagapudi, V.; De Brabander, J.K.; Kainov, D.E.; Vapalahti, O. Obatoclax, saliphenylhalamide and gemcitabine inhibit Zika virus infection in vitro and differentially affect cellular signaling, transcription and metabolism. Antivir. Res. 2017, 139, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.S.; Rausalu, K.; Hakanen, M.; Saul, S.; Kümmerer, B.M.; Susi, P.; Merits, A.; Ahola, T. Obatoclax Inhibits Alphavirus Membrane Fusion by Neutralizing the Acidic Environment of Endocytic Compartments. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Denisova, O.V.; Kakkola, L.; Feng, L.; Stenman, J.; Nagaraj, A.; Lampe, J.; Yadav, B.; Aittokallio, T.; Kaukinen, P.; Ahola, T.; et al. Obatoclax, Saliphenylhalamide, and Gemcitabine Inhibit Influenza A Virus Infection. J. Boil. Chem. 2012, 287, 35324–35332. [Google Scholar] [CrossRef] [Green Version]
- Shupack, J.L.; Grieco, A.J.; Epstein, A.M.; Sansaricq, C.; E Snyderman, S. Azaribine, Homocystinemia, and Thrombosis. Arch. Dermatol. 1977, 113, 1301. [Google Scholar] [CrossRef]
- Morrey, J.; Smee, D.F.; Sidwell, R.W.; Tseng, C. Identification of active antiviral compounds against a New York isolate of West Nile virus. Antivir. Res. 2002, 55, 107–116. [Google Scholar] [CrossRef]
- Pascoalino, B.S.; Courtemanche, G.; Cordeiro, M.T.; Gil, L.H.V.G.; Freitas-Junior, L.H. Zika antiviral chemotherapy: Identification of drugs and promising starting points for drug discovery from an FDA-approved library. F1000Research 2016, 5, 2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briolant, S.; Garin, D.; Scaramozzino, N.; Jouan, A.; Crance, J. In vitro inhibition of Chikungunya and Semliki Forest viruses replication by antiviral compounds: Synergistic effect of interferon-α and ribavirin combination. Antivir. Res. 2004, 61, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Crance, J.M.; Scaramozzino, N.; Jouan, A.; Garin, D. Interferon, ribavirin, 6-azauridine and glycyrrhizin: Antiviral compounds active against pathogenic flaviviruses. Antivir. Res. 2003, 58, 73–79. [Google Scholar] [CrossRef]
- Rada, B.; Dragúň, M. Antiviral action and selectivity of 6-azauridine. Ann. N. Y. Acad. Sci. 1977, 284, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Adcock, R.S.; Chu, Y.-K.; Golden, J.E.; Chung, N.-H. Evaluation of anti-Zika virus activities of broad-spectrum antivirals and NIH clinical collection compounds using a cell-based, high-throughput screen assay. Antivir. Res. 2017, 138, 47–56. [Google Scholar] [CrossRef]
- Westhead, J.E.; Price, H.D. Quantitative Assay of Pyrazofurin a New Antiviral, Antitumor Antibiotic. Antimicrob. Agents Chemother. 1974, 5, 90–91. [Google Scholar] [CrossRef] [Green Version]
- De Clercq, E. New Nucleoside Analogues for the Treatment of Hemorrhagic Fever Virus Infections. Chem. Asian J. 2019, 14, 3962–3968. [Google Scholar] [CrossRef]
- Mei-Jiao, G.; Shi-Fang, L.; Yan-Yan, C.; Jun-Jun, S.; Yue-Feng, S.; Ting-Ting, R.; Yong-Guang, Z.; Hui-Yun, C. Antiviral effects of selected IMPDH and DHODH inhibitors against foot and mouth disease virus. Biomed. Pharmacother. 2019, 118, 109305. [Google Scholar] [CrossRef]
- Sievers, T.M.; Rossi, S.J.; Ghobrial, R.M.; Arriola, E.; Nishimura, P.; Kawano, M.; Holt, C.D. Mycophenolate mofetil. Pharmacotherapy 1997, 17, 1178–1197. [Google Scholar]
- Cho, J.; Yi, H.; Jang, E.Y.; Lee, M.-S.; Lee, J.-Y.; Kang, C.; Lee, C.H.; Kim, K. Mycophenolic mofetil, an alternative antiviral and immunomodulator for the highly pathogenic avian influenza H5N1 virus infection. Biochem. Biophys. Res. Commun. 2017, 494, 298–304. [Google Scholar] [CrossRef]
- Allison, A.C. Mechanisms of action of mycophenolate mofetil. Lupus 2005, 14, 2–8. [Google Scholar] [CrossRef]
- Dong, S.; Kang, S.; Dimopoulos, G. Identification of anti-flaviviral drugs with mosquitocidal and anti-Zika virus activity in Aedes aegypti. PLoS Neglected Trop. Dis. 2019, 13, e0007681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Van Vuuren, A.J.; Van Der Laan, L.J.W.; Peppelenbosch, M.P.; Janssen, H.L.A. Antiviral or proviral action of mycophenolic acid in hepatitis B infection? Hepatology 2012, 56, 1586–1587. [Google Scholar] [CrossRef]
- Manchala, N.R.; Dungdung, R.; Trivedi, P.; Unniyampurath, U.; Pilankatta, R. Mycophenolic acid (MPA) modulates host cellular autophagy progression in sub genomic dengue virus-2 replicon cells. Microb. Pathog. 2019, 137, 103762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, K.K.-W.; Mok, K.-Y.; Chan, A.S.F.; Cheung, N.N.; Wang, P.; Lui, Y.-M.; Chan, J.F.; Chen, H.; Chan, K.-H.; Kao, R.Y.; et al. Mycophenolic acid, an immunomodulator, has potent and broad-spectrum in vitro antiviral activity against pandemic, seasonal and avian influenza viruses affecting humans. J. Gen. Virol. 2016, 97, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Smee, D.F.; Bray, M.; Huggins, J.W. Antiviral activity and mode of action studies of ribavirin and mycophenolic acid against orthopoxviruses in vitro. Antivir. Chem. Chemother. 2001, 12, 327–335. [Google Scholar] [CrossRef]
- Hermann, L.L.; Coombs, K.M. Inhibition of Reovirus by Mycophenolic Acid Is Associated with the M1 Genome Segment. J. Virol. 2004, 78, 6171–6179. [Google Scholar] [CrossRef] [Green Version]
- Henry, S.D.; Metselaar, H.J.; Lonsdale, R.C.; Kok, A.; Haagmans, B.L.; Tilanus, H.W.; Van Der Laan, L.J.W. Mycophenolic Acid Inhibits Hepatitis C Virus Replication and Acts in Synergy With Cyclosporin A and Interferon-α. Gastroenterology 2006, 131, 1452–1462. [Google Scholar] [CrossRef]
- Albulescu, I.C.; Kovacikova, K.; Tas, A.; Snijder, E.J.; Van Hemert, M.J. Suramin inhibits Zika virus replication by interfering with virus attachment and release of infectious particles. Antivir. Res. 2017, 143, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Barrows, N.J.; Campos, R.K.; Powell, S.T.; Prasanth, K.R.; Schott-Lerner, G.; Soto-Acosta, R.; Galarza-Muñoz, G.; McGrath, E.L.; Urrabaz-Garza, R.; Gao, J.; et al. A Screen of FDA-Approved Drugs for Inhibitors of Zika Virus Infection. Cell Host Microbe 2016, 20, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Floryk, D.; Thompson, T.C. Antiproliferative effects of AVN944, a novel inosine 5-monophosphate dehydrogenase inhibitor, in prostate cancer cells. Int. J. Cancer 2008, 123, 2294–2302. [Google Scholar] [CrossRef] [Green Version]
- Dunham, E.C.; Leske, A.; Shifflett, K.; Watt, A.; Feldmann, H.; Hoenen, T.; Groseth, A. Lifecycle modelling systems support inosine monophosphate dehydrogenase (IMPDH) as a pro-viral factor and antiviral target for New World arenaviruses. Antivir. Res. 2018, 157, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-F.; Gong, M.-J.; Sun, Y.-F.; Shao, J.-J.; Zhang, Y.-G.; Chang, H. Antiviral activity of brequinar against foot-and-mouth disease virus infection in vitro and in vivo. Biomed. Pharmacother. 2019, 116, 108982. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.I.; Krpina, K.; Ianevski, A.; Shtaida, N.; Jo, E.; Yang, J.; Koit, S.; Tenson, T.; Hukkanen, V.; Anthonsen, M.W.; et al. Novel Antiviral Activities of Obatoclax, Emetine, Niclosamide, Brequinar, and Homoharringtonine. Viruses 2019, 11, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, M.; Zou, G.; Wang, Q.-Y.; Xu, H.Y.; Dong, H.; Yuan, Z.; Shi, P.-Y. Characterization of Dengue Virus Resistance to Brequinar in Cell Culture. Antimicrob. Agents Chemother. 2010, 54, 3686–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Ding, S.; Yin, Y.; Xu, L.; Li, P.; Peppelenbosch, M.P.; Pan, Q.; Wang, W. Suppression of pyrimidine biosynthesis by targeting DHODH enzyme robustly inhibits rotavirus replication. Antivir. Res. 2019, 167, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Luthra, P.; Naidoo, J.; Pietzsch, C.A.; De, S.; Khadka, S.; Anantpadma, M.; Williams, C.G.; Edwards, M.R.; Davey, R.A.; Bukreyev, A.; et al. Inhibiting pyrimidine biosynthesis impairs Ebola virus replication through depletion of nucleoside pools and activation of innate immune responses. Antivir. Res. 2018, 158, 288–302. [Google Scholar] [CrossRef]
- Schnellrath, L.C.; Damaso, C.R. Potent antiviral activity of brequinar against the emerging Cantagalo virus in cell culture. Int. J. Antimicrob. Agents 2011, 38, 435–441. [Google Scholar] [CrossRef] [Green Version]









| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | 51.28 | >1350.00 | 2.00 | 25.64 | >675.00 |
| OSU-03012 | 6.39 | 132.50 | 8.40 | 0.76 | 15.77 |
| Obatoclax | 17.57 | 60.44 | 0.63 | 27.89 | 95.94 |
| Azauridine | 155.80 | >1350.00 | 4.29 | 36.32 | >314.69 |
| Azaribine | 237.00 | >1350.00 | 1.62 | 146.29 | >833.33 |
| Pyrazofurin | 270.60 | >1350.00 | 5.96 | 45.35 | >226.51 |
| Mycophenolate mofetil | 166.30 | >1350.00 | 3.52 | 47.24 | >383.52 |
| Mycophenolic acid | 275.40 | >1350.00 | 4.26 | 64.65 | >316.90 |
| AVN-944 | 272.60 | >1350.00 | 0.98 | 278.16 | >1377.55 |
| Brequinar | 237.70 | >1350.00 | 1.51 | 157.42 | >894.04 |
| ATA | >1350.00 | >1350.00 | 25.00 | >54.00 | >54.00 |
| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | 51.28 | >1350.00 | 8.95 | 5.73 | >150.84 |
| OSU-03012 | 6.39 | 132.50 | 0.93 | 6.87 | 142.47 |
| Obatoclax | 17.57 | 60.44 | 2.96 | 5.94 | 20.4 |
| Azauridine | 155.80 | >1350.00 | 64.49 | 2.42 | >20.93 |
| Azaribine | 237.00 | >1350.00 | 47.29 | 5.01 | >28.55 |
| Pyrazofurin | 270.60 | >1350.00 | 313.00 | 0.86 | >4.31 |
| Mycophenolate mofetil | 166.30 | >1350.00 | 8.11 | 20.51 | >166.46 |
| Mycophenolic acid | 275.40 | >1350.00 | 7.58 | 36.33 | >178.10 |
| AVN-944 | 272.60 | >1350.00 | 11.12 | 24.51 | >121.40 |
| Brequinar | 237.70 | >1350.00 | 42.47 | 5.59 | >31.79 |
| ATA | >1350.00 | >1350.00 | 9.86 | >136.92 | >136.92 |
| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | 51.28 | >1350.00 | 3.50 | 14.65 | >385.71 |
| OSU-03012 | 6.39 | 132.50 | 1.77 | 3.61 | 36.70 |
| Obatoclax | 17.57 | 60.44 | 0.29 | 60.58 | 208.41 |
| Azauridine | 155.80 | >1350.00 | 130.30 | 1.19 | >10.36 |
| Azaribine | 237.00 | >1350.00 | 2.59 | 91.51 | >521.23 |
| Pyrazofurin | 270.60 | >1350.00 | 39.07 | 6.93 | >34.55 |
| Mycophenolate mofetil | 166.30 | >1350.00 | 2.27 | 73.26 | >594.71 |
| Mycophenolic acid | 275.40 | >1350.00 | 41.75 | 6.59 | >32.33 |
| AVN-944 | 272.60 | >1350.00 | 7.98 | 34.16 | >169.17 |
| Brequinar | 237.70 | >1350.00 | 8.71 | 27.29 | >154.99 |
| ATA | >1350.00 | >1350.00 | 1.73 | >780.34 | >780.34 |
| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | 51.28 | >1350.00 | 0.03 | 1709.33 | >45,000.00 |
| OSU-03012 | 6.39 | 132.50 | 0.81 | 7.89 | 163.58 |
| Obatoclax | 17.57 | 60.44 | 0.37 | 47.49 | 163.35 |
| Azauridine | 155.80 | >1350.00 | 0.76 | 205.00 | >1776.32 |
| Azaribine | 237.00 | >1350.00 | 2.43 | 97.53 | >555.56 |
| Pyrazofurin | 270.60 | >1350.00 | 0.89 | 304.04 | >1516.85 |
| Mycophenolate mofetil | 166.30 | >1350.00 | 0.24 | 692.92 | >5625.00 |
| Mycophenolic acid | 275.40 | >1350.00 | 0.15 | 1836.00 | >9000.00 |
| AVN-944 | 272.60 | >1350.00 | 0.33 | 826.06 | >4090.91 |
| Brequinar | 237.70 | >1350.00 | 0.05 | 4754.00 | >27,000.00 |
| ATA | >1350.00 | >1350.00 | 8.08 | >167.08 | >167.08 |
| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | 51.28 | >1350.00 | 7.32 | 7.01 | >184.43 |
| OSU-03012 | 6.39 | 132.50 | 1.68 | 3.80 | 78.87 |
| Obatoclax | 17.57 | 60.44 | 0.41 | 42.85 | 147.41 |
| Azauridine | 155.80 | >1350.00 | 1.46 | 106.71 | >924.66 |
| Azaribine | 237.00 | >1350.00 | 1.12 | 211.61 | >1205.36 |
| Pyrazofurin | 270.60 | >1350.00 | 0.95 | 284.84 | >1421.06 |
| Mycophenolate mofetil | 166.30 | >1350.00 | 0.12 | 1385.83 | >11,250.00 |
| Mycophenolic acid | 275.40 | >1350.00 | 0.22 | 1251.82 | >6136.36 |
| AVN-944 | 272.60 | >1350.00 | 0.38 | 717.37 | >3552.63 |
| Brequinar | 237.70 | >1350.00 | 0.08 | 2971.25 | >16,875.00 |
| ATA | >1350.00 | >1350.00 | 4.74 | >284.81 | >284.81 |
| Compound | CC50 (MTT) (µM) | CC50 (XTT) (µM) | EC50 (µM) | SI (MTT) | SI (XTT) |
|---|---|---|---|---|---|
| Antimycin A | >50.00 | >50.00 | 1.71 µM | >29.24 | >29.24 |
| OSU-03012 | >50.00 | >50.00 | 0.21 µM | >238.09 | >238.09 |
| Obatoclax | >50.00 | >50.00 | 0.64 µM | >78.13 | >78.13 |
| Azauridine | >50.00 | >50.00 | 0.44 µM | >113.64 | >113.64 |
| Azaribine | >50.00 | >50.00 | 0.68 µM | >73.53 | >73.53 |
| Pyrazofurin | >50.00 | >50.00 | 0.33 µM | >151.52 | >151.52 |
| Mycophenolate mofetil | >50.00 | >50.00 | 0.83 µM | >60.24 | >60.24 |
| Mycophenolic acid | >50.00 | >50.00 | 0.65 µM | >76.92 | >76.92 |
| AVN-944 | >50.00 | >50.00 | 0.05 µM | >1000.00 | >1000.00 |
| Brequinar | >50.00 | >50.00 | 0.02 µM | >2500.00 | >2500.00 |
| ATA | >50.00 | >50.00 | 6.67 µM | >7.49 | >7.49 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales Vasquez, D.; Park, J.-G.; Ávila-Pérez, G.; Nogales, A.; de la Torre, J.C.; Almazan, F.; Martinez-Sobrido, L. Identification of Inhibitors of ZIKV Replication. Viruses 2020, 12, 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091041
Morales Vasquez D, Park J-G, Ávila-Pérez G, Nogales A, de la Torre JC, Almazan F, Martinez-Sobrido L. Identification of Inhibitors of ZIKV Replication. Viruses. 2020; 12(9):1041. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091041
Chicago/Turabian StyleMorales Vasquez, Desarey, Jun-Gyu Park, Ginés Ávila-Pérez, Aitor Nogales, Juan Carlos de la Torre, Fernando Almazan, and Luis Martinez-Sobrido. 2020. "Identification of Inhibitors of ZIKV Replication" Viruses 12, no. 9: 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/v12091041