
Environmental Surveillance through Next-Generation Sequencing to Unveil the Diversity of Human Enteroviruses beyond the Reported Clinical Cases

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Human Enteroviruses Detection and Sequencing
2.3. Bioinformatics Pipeline
2.4. Phylogenetic Analyses and Genetic Diversity
2.5. Backgrounds of Human Enteroviruses Detection in Argentina. Link between Environmental and Clinical Isolates
3. Results
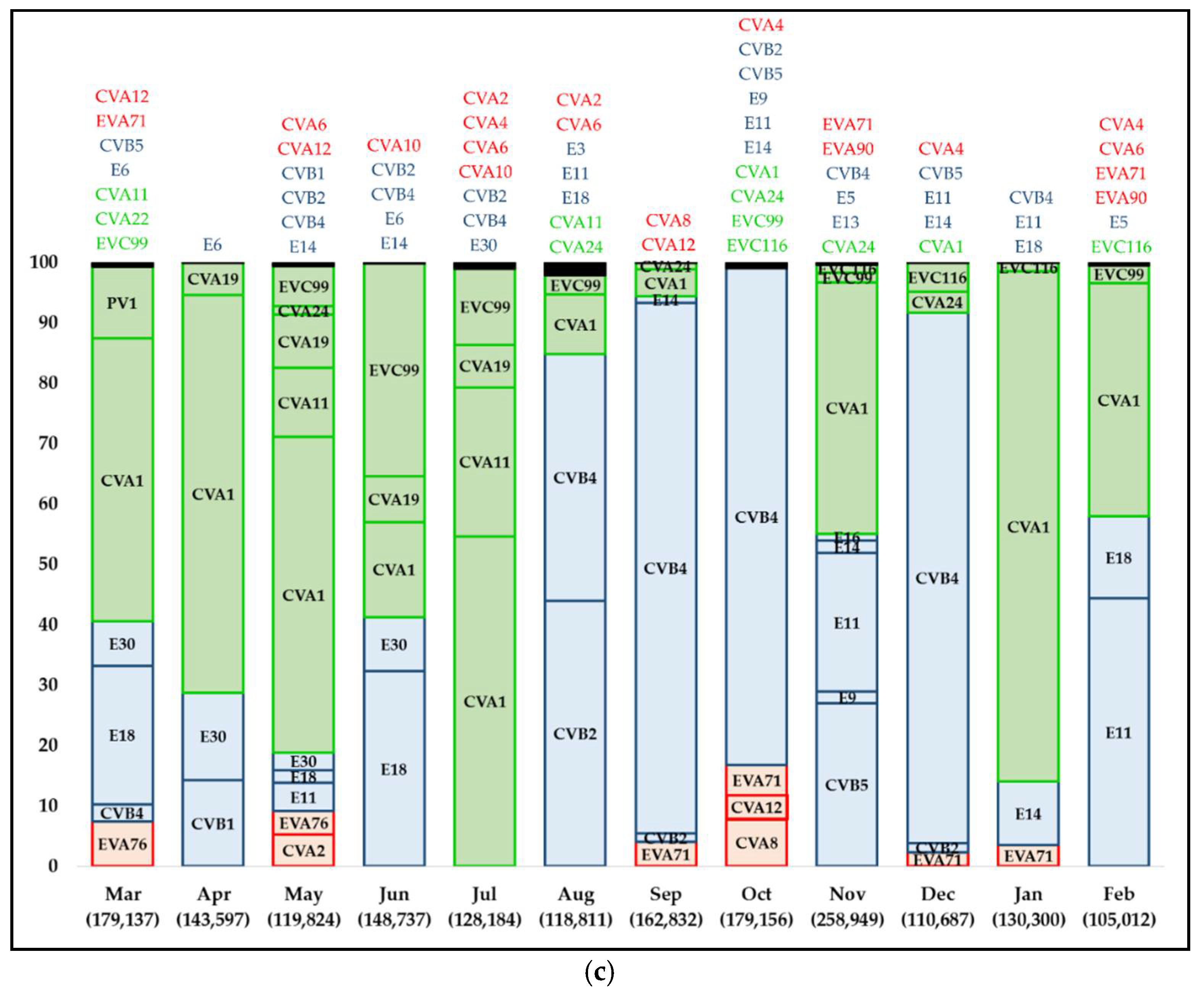
3.1. Taxonomic Allocation and Distribution of Human Enteroviruses Species’ Frequency and Abundance
3.2. Human Enteroviruses Characterization
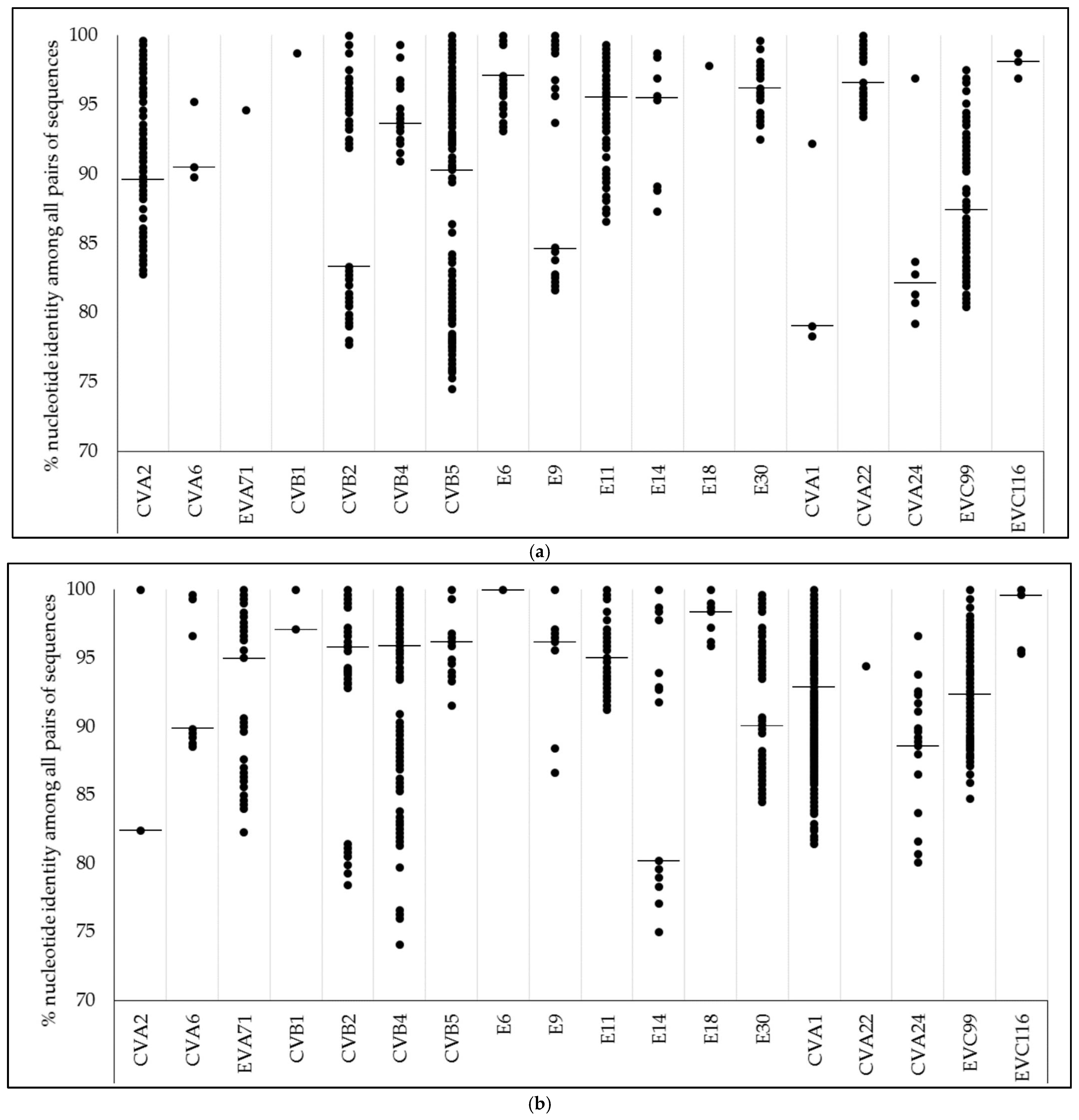
3.3. Nucleotide Identity
3.4. Backgrounds of Human Enteroviruses Detection in Argentina
3.5. Link between Human Enteroviruses Detected in Wastewater and Clinical Isolates
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zell, R.; Delwart, E.; Gorbalenya, A.E.; Hovi, T.; King, A.M.Q.; Knowles, N.J.; Lindberg, A.M.; Pallansch, M.A.; Palmenberg, A.C.; Reuter, G.; et al. ICTV Virus Taxonomy Profile: Picornaviridae. J. Gen. Virol. 2017, 98, 2421–2422. [Google Scholar] [CrossRef]
- Muehlenbachs, A.; Bhatnagar, J.; Zaki, S.R. Tissue tropism, pathology and pathogenesis of enterovirus infection. J. Pathol. 2014, 235, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stalkup, J.R.; Chilukuri, S. Enterovirus infections: A review of clinical presentation, diagnosis, and treatment. Dermatol. Clin. 2002, 20, 217–223. [Google Scholar] [CrossRef]
- Tapparel, C.; Siegrist, F.; Petty, T.J.; Kaiser, L. Picornavirus and enterovirus diversity with associated human diseases. Infect. Genet. Evol. 2013, 14, 282–293. [Google Scholar] [CrossRef] [PubMed]
- De Crom, S.C.M.; Rossen, J.W.A.; Van Furth, A.M.; Obihara, C.C. Enterovirus and parechovirus infection in children: A brief overview. Eur. J. Nucl. Med. Mol. Imaging 2016, 175, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. National Center for Immunization and Respiratory Diseases (NCIRD), Division of Viral Diseases Non-Polio Enterovirus. Transmission. Available online: https://www.cdc.gov/non-polio-enterovirus/about/transmission.html (accessed on 30 March 2020).
- Melnick, J.L.; Rennick, V. Infectivity liters of enterovirus as found in human stools. J. Med. Virol. 1980, 5, 205–220. [Google Scholar] [CrossRef]
- Tao, Z.; Wang, Z.; Lin, X.; Wang, S.; Wang, H.; Yoshida, H.; Xu, A.; Song, Y. One-year Survey of human enteroviruses from sewage and the factors affecting virus adsorption to the suspended solids. Sci. Rep. 2016, 6, 31474. [Google Scholar] [CrossRef]
- Benschop, K.S.M.; Van Der Avoort, H.G.; Jusic, E.; Vennema, H.; Van Binnendijk, R.; Duizer, E. Polio and Measles Down the Drain: Environmental Enterovirus Surveillance in the Netherlands, 2005 to 2015. Appl. Environ. Microbiol. 2017, 83, e00558-17. [Google Scholar] [CrossRef] [Green Version]
- Lukashev, A.N.; Vakulenko, Y.A.; Turbabina, N.A.; Deviatkin, A.A.; Drexler, J.F. Molecular epidemiology and phylogenetics of human enteroviruses: Is there a forest behind the trees? Rev. Med. Virol. 2018, 28, e2002. [Google Scholar] [CrossRef]
- Palacios, G.; Casas, I.; Cisterna, D.; Trallero, G.; Tenorio, A.; Freire, C. Molecular Epidemiology of Echovirus 30: Temporal Circulation and Prevalence of Single Lineages. J. Virol. 2002, 76, 4940–4949. [Google Scholar] [CrossRef] [Green Version]
- Farías, A.; Cabrerizo, M.; Ré, V.; Glatstein, N.; Pisano, B.; Spinsanti, L.; Contigiani, M.S. Molecular identification of human enteroviruses in children with neurological infections from the central region of Argentina. Arch. Virol. 2010, 156, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Lema, C.; Torres, C.; Van Der Sanden, S.; Cisterna, D.M.; Freire, M.C.; Gomez, R.M. Global phylodynamics of Echovirus 30 revealed differential behavior among viral lineages. Virology 2019, 531, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Freire, M.C.; Cisterna, D.M.; Rivero, K.; Palacios, G.; Casas, I.; Tenorio, A.; Gomez, J.A. Análisis de un brote de meningitis viral en la provincia de Tucumán, Argentina. Rev. Panam. Salud. Pública. 2003, 13, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Grenón, S.L.; Robledo, M.L.; von Specht, M.H.; Cisterna, D.M.; Lema, C.L.; Freire, M.C. Brote epidémico de meningitis viral causado por echovirus tipo 4 en la provincia de Misiones. Rev. Argent. Microbiol. 2008, 40, 41–46. [Google Scholar]
- Ruggieri, V.; Paz, M.I.; Peretti, M.G.; Rugilo, C.; Bologna, R.; Freire, C.; Vergel, S.; Savransky, A. Enterovirus D68 infection in a cluster of children with acute flaccid myelitis, Buenos Aires, Argentina, 2016. Eur. J. Paediatr. Neurol. 2017, 21, 884–890. [Google Scholar] [CrossRef]
- Cisterna, D.M.; Lema, C.L.; Martinez, L.M.; Verón, E.; Contarino, L.P.; Acosta, D.; Freire, M.C. Atypical hand, foot, and mouth disease caused by Coxsackievirus A6 in Argentina in 2015. Rev. Argent. Microbiol. 2019, 51, 140–143. [Google Scholar] [CrossRef]
- Cisterna, D.; Lema, C.; Riviello López, G.; Freyre, L.; Martínez, L.; Torres, C.; Mbayed, V.; Freire, M.C. Detección y Caracterización de Enterovirus y Hepatitis A en el Riachuelo y Río de la Plata. In Libro de Resúmenes del IX Congreso Argentino de Virología 2008. Buenos Aires, Argentina, 2008; Revista Argentina de Microbiología: Buenos Aires, Argentina, 2008; abstract number 69-22999; pp. 49–50. [Google Scholar]
- Mueller, J.E.; Bessaud, M.; Huang, S.; Martinez, L.C.; Barril, P.A.; Morel, V.; Balanant, J.; Bocacao, J.; Hewitt, J.; Gessner, B.D.; et al. Environmental Poliovirus Surveillance during Oral Poliovirus Vaccine and Inactivated Poliovirus Vaccine Use in Córdoba Province, Argentina. Appl. Environ. Microbiol. 2009, 75, 1395–1401. [Google Scholar] [CrossRef] [Green Version]
- Farías, A.A.; Mojsiejczuk, L.; Pisano, M.B.; Flores, F.S.; Aguilar, J.J.; Jean, A.N.; Yanes, L.A.; Masachessi, G.; Prez, V.E.; Isa, M.B.; et al. Environmental Surveillance of Enteroviruses in Central Argentina: First Detection and Evolutionary Analyses of E14. Food Environ. Virol. 2018, 10, 121–126. [Google Scholar] [CrossRef]
- Farías, A.A.; Mojsiejczuk, L.; Flores, F.S.; Aguilar, J.J.; Prez, V.E.; Masachessi, G.; Nates, S.V. Environmental Surveillance of Human Enteroviruses in Córdoba City, Argentina: Prevalence and Detection of Serotypes from 2009 to 2014. Food Environ. Virol. 2019, 11, 198–203. [Google Scholar] [CrossRef]
- Mistchenko, A.S.; Viegas, M.; Latta, M.P.; Barrero, P.R. Molecular and epidemiologic analysis of enterovirus B neurological infection in Argentine children. J. Clin. Virol. 2006, 37, 293–299. [Google Scholar] [CrossRef]
- Cisterna, D.M.; Palacios, G.; Rivero, K.; Girard, D.; Lema, C.; Freire, M.C. Epidemiology of enterovirus associated with neurologic diseases [in Spanish]. Medicina (B Aires) 2007, 67, 113–119. [Google Scholar] [PubMed]
- Wang, H.; Tao, Z.; Li, Y.; Lin, X.; Yoshida, H.; Song, L.; Zhang, Y.; Wang, S.; Cui, N.; Xu, W.; et al. Environmental Surveillance of Human Enteroviruses in Shandong Province, China, 2008 to 2012: Serotypes, Temporal Fluctuation, and Molecular Epidemiology. Appl. Environ. Microbiol. 2014, 80, 4683–4691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joffret, M.-L.; Polston, P.M.; Razafindratsimandresy, R.; Bessaud, M.; Heraud, J.-M.; Delpeyroux, F. Whole Genome Sequencing of Enteroviruses Species A to D by High-Throughput Sequencing: Application for Viral Mixtures. Front. Microbiol. 2018, 9, 2339. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, M.; Sharif, S.; Klapsa, D.; Wilton, T.; Alam, M.M.; Fernandez-Garcia, M.D.; Rehman, L.; Mujtaba, G.; McAllister, G.; Harvala, H.; et al. Environmental Surveillance Reveals Complex Enterovirus Circulation Patterns in Human Populations. Open Forum Infect. Dis. 2018, 5, ofy250. [Google Scholar] [CrossRef] [PubMed]
- Bisseux, M.; Debroas, D.; Mirand, A.; Archimbaud, C.; Peigue-Lafeuille, H.; Bailly, J.-L.; Henquell, C.; Didier, D.; Audrey, M.; Christine, A.; et al. Monitoring of enterovirus diversity in wastewater by ultra-deep sequencing: An effective complementary tool for clinical enterovirus surveillance. Water Res. 2020, 169, 115246. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Puchol, S.; Rusiñol, M.; Fernández-Cassi, X.; Timoneda, N.; Itarte, M.; Andrés, C.; Antón, A.; Abril, J.F.; Girones, R.; Bofill-Mas, S. Characterisation of the sewage virome: Comparison of NGS tools and occurrence of significant pathogens. Sci. Total. Environ. 2020, 713, 136604. [Google Scholar] [CrossRef]
- Masachessi, G.; Pisano, M.B.; Prez, V.E.; Martínez, L.C.; Michelena, J.F.; Martínez-Wassaf, M.; Giordano, M.O.; Isa, M.B.; Pavan, J.V.; Welter, A.; et al. Enteric Viruses in Surface Waters from Argentina: Molecular and Viable-Virus Detection. Appl. Environ. Microbiol. 2017, 84, e02327-17. [Google Scholar] [CrossRef] [Green Version]
- Instituto Nacional de Estadística y Censos de la República Argentina. Censo Nacional de Población, Hogares y Viviendas. Available online: https://www.indec.gob.ar/indec/web/Nivel4-Tema-2-41-135 (accessed on 13 January 2021).
- Nix, W.A.; Oberste, M.S.; Pallansch, M.A. Sensitive, Seminested PCR Amplification of VP1 Sequences for Direct Identification of All Enterovirus Serotypes from Original Clinical Specimens. J. Clin. Microbiol. 2006, 44, 2698–2704. [Google Scholar] [CrossRef] [Green Version]
- Illumina support web-site. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 14 December 2020).
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H.; Hasegawa, M. CONSEL: For assessing the confidence of phylogenetic tree selection. Bioinformatics 2001, 17, 1246–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate Likelihood-Ratio Test for Branches: A Fast, Accurate, and Powerful Alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequences alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sapia, E.Y.; Maroni, C.; Groisman, C.; Kromer, H.; Rojo, G.L.; Dastugue, M.; Valinotto, L. Genotipificación del virus de la enfermedad mano-pie-boca atípica en un hospital pediátrico de la Ciudad de Buenos Aires, Argentina. Arch. Argent. Pediatr. 2020, 118, e199–e203. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.A.; Maher, K.; Flemister, M.R.; Naraghi-Arani, P.; Uddin, M.; Oberste, M.S.; Pallansch, M.A. Resolving ambiguities in genetic typing of human enterovirus species C clinical isolates and identification of enterovirus 96, 99 and 102. J. Gen. Virol. 2009, 90, 1713–1723. [Google Scholar] [CrossRef]
- Adeniji, J.A.; Faleye, T.O.C. Isolation and Identification of Enteroviruses from Sewage and Sewage-Contaminated Water in Lagos, Nigeria. Food Environ. Virol. 2014, 6, 75–86. [Google Scholar] [CrossRef]
- Wieczorek, M.; Ciąćka, A.; Witek, A.; Kuryk, Ł.; Żuk-Wasek, A. Environmental Surveillance of Non-polio Enteroviruses in Poland, 2011. Food Environ. Virol. 2015, 7, 224–231. [Google Scholar] [CrossRef]
- Prez, V.; Martínez, L.; Victoria, M.; Giordano, M.; Masachessi, G.; Ré, V.; Pavan, J.; Colina, R.; Barril, P.; Nates, S. Tracking enteric viruses in green vegetables from central Argentina: Potential association with viral contamination of irrigation waters. Sci. Total. Environ. 2018, 665–671. [Google Scholar] [CrossRef]
- Pons-Salort, M.; Oberste, M.S.; Pallansch, M.A.; Abedi, G.R.; Takahashi, S.; Grenfell, B.T.; Grassly, N.C. The seasonality of nonpolio enteroviruses in the United States: Patterns and drivers. Proc. Natl. Acad. Sci. USA 2018, 115, 3078–3083. [Google Scholar] [CrossRef] [Green Version]
- Brinkman, N.E.; Fout, G.S.; Keely, S.P. Retrospective Surveillance of Wastewater To Examine Seasonal Dynamics of Enterovirus Infections. mSphere 2017, 2, e00099-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baicus, A.; Joffret, M.-L.; Bessaud, M.; Delpeyroux, F.; Oprisan, G. Reinforced poliovirus and enterovirus surveillance in Romania, 2015–2016. Arch. Virol. 2020, 165, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Chen, P.; Cui, N.; Lin, X.; Ji, F.; Liu, Y.; Xiong, P.; Zhang, L.; Xu, Q.; Song, Y.; et al. Detection of enteroviruses in urban sewage by next generation sequencing and its application in environmental surveillance. Sci. Total. Environ. 2020, 728, 138818. [Google Scholar] [CrossRef] [PubMed]
- Harvala, H.; Calvert, J.; Van Nguyen, D.; Clasper, L.; Gadsby, N.; Molyneaux, P.; Templeton, K.; Leitch, C.M.; Simmonds, P. Comparison of diagnostic clinical samples and environmental sampling for enterovirus and parechovirus surveillance in Scotland, 2010 to 2012. Eurosurveillance 2014, 19, 20772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndiaye, A.K.; Mbathio, P.A.; Diop, P.A.M. Environmental surveillance of poliovirus and non-polio enterovirus in urban sewage in Dakar, Senegal (2007-2013). Pan Afr. Med. J. 2014, 19, 243. [Google Scholar] [CrossRef]
- Brown, B.; Oberste, M.S.; Maher, K.; Pallansch, M.A. Complete Genomic Sequencing Shows that Polioviruses and Members of Human Enterovirus Species C Are Closely Related in the Noncapsid Coding Region. J. Virol. 2003, 77, 8973–8984. [Google Scholar] [CrossRef] [Green Version]
- Kew, O.; Pallansch, M. Breaking the Last Chains of Poliovirus Transmission: Progress and Challenges in Global Polio Eradication. Annu. Rev. Virol. 2018, 5, 427–451. [Google Scholar] [CrossRef]
- Jiang, P.; Faase, J.A.J.; Toyoda, H.; Paul, A.; Wimmer, E.; Gorbalenya, A.E. Evidence for emergence of diverse polioviruses from C-cluster coxsackie A viruses and implications for global poliovirus eradication. Proc. Natl. Acad. Sci. USA 2007, 104, 9457–9462. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Lin, X.; Ji, F.; Xiong, P.; Liu, Y.; Wang, S.; Chen, P.; Xu, Q.; Zhang, L.; Tao, Z.; et al. Prevalence and Bayesian Phylogenetics of Enteroviruses Derived From Environmental Surveillance Around Polio Vaccine Switch Period in Shandong Province, China. Food Environ. Virol. 2020, 12, 321–332. [Google Scholar] [CrossRef]
- Faleye, T.O.C.; Adeniji, J.A. Enterovirus Species B Bias of RD Cell Line and Its Influence on Enterovirus Diversity Landscape. Food Environ. Virol. 2015, 7, 390–402. [Google Scholar] [CrossRef]
- Bubba, L.; Broberg, E.K.; Jasir, A.; Simmonds, P.; Harvala, H.; Redlberger-Fritz, M.; Nikolaeva-Glomb, L.; Havlíčková, M.; Rainetova, P.; Fischer, T.K.; et al. Circulation of non-polio enteroviruses in 24 EU and EEA countries between 2015 and 2017: A retrospective surveillance study. Lancet Infect. Dis. 2020, 20, 350–361. [Google Scholar] [CrossRef]
- Savolainen-Kopra, C.; Paananen, A.; Blomqvist, S.; Klemola, P.; Simonen, M.-L.; Lappalainen, M.; Vuorinen, T.; Kuusi, M.; Lemey, P.; Roivainen, M. A large Finnish echovirus 30 outbreak was preceded by silent circulation of the same genotype. Virus Genes 2010, 42, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, O.E.; Yarmolskaya, M.S.; Eremeeva, T.P.; Babkina, G.M.; Baikova, O.Y.; Akhmadishina, L.V.; Krasota, A.Y.; Kozlovskaya, L.I.; Lukashev, A.N. Environmental Surveillance for Poliovirus and Other Enteroviruses: Long-Term Experience in Moscow, Russian Federation, 2004–2017. Viruses 2019, 11, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedmak, G.; Bina, D.; Macdonald, J. Assessment of an Enterovirus Sewage Surveillance System by Comparison of Clinical Isolates with Sewage Isolates from Milwaukee, Wisconsin, Collected August 1994 to December 2002. Appl. Environ. Microbiol. 2003, 69, 7181–7187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Z.; Song, Y.; Wang, H.; Zhang, Y.; Yoshida, H.; Ji, S.; Xu, A.; Song, L.; Liu, Y.; Cui, N.; et al. Intercity Spread of Echovirus 6 in Shandong Province, China: Application of Environmental Surveillance in Tracing Circulating Enteroviruses. Appl. Environ. Microbiol. 2012, 78, 6946–6953. [Google Scholar] [CrossRef] [Green Version]
- Harvala, H.; Broberg, E.; Benschop, K.; Berginc, N.; Ladhani, S.N.; Susi, P.; Christiansen, C.; McKenna, J.; Allen, D.; Makiello, P.; et al. Recommendations for enterovirus diagnostics and characterisation within and beyond Europe. J. Clin. Virol. 2018, 101, 11–17. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | No. Contigs * | No. Contigs Represented by Centroids (n) after Clusterization, and Quality Filtering | No. Contigs and Centroids (n) Mapping to CHED § | % of Mapped Contigs | No. Types Detected | Normalization (No. Types Detected/100,000 Contigs) |
|---|---|---|---|---|---|---|
| Jan-11 | 100,139 | 88,677 (15) | 88,624 (14) | 99.940 | 8 | 9 |
| Feb-11 | 94,803 | 84,740 (12) | 84,738 (11) | 99.998 | 9 | 11 |
| Mar-11 | 113,365 | 100,538 (11) | 100,509 (8) | 99.971 | 6 | 6 |
| Apr-11 | 109,599 | 95,364 (13) | 95,362 (12) | 99.998 | 8 | 8 |
| May-11 | 112,312 | 94,882 (12) | 94,880 (11) | 99.998 | 5 | 5 |
| Jun-11 | 108,516 | 94,081 (10) | 63,388 (9) | 67.376 | 6 | 9 |
| Jul-11 | 109,250 | 98,124 (11) | 98,039 (5) | 99.913 | 5 | 5 |
| Aug-11 | 110,859 | 96,682 (11) | 96,677 (10) | 99.995 | 6 | 6 |
| Sep-11 | 110,270 | 99,735 (5) | 99,735 (5) | 100 | 5 | 5 |
| Oct-11 | 63,293 | 50,828 (7) | 50,828 (7) | 100 | 5 | 10 |
| Nov-11 | 110,198 | 85,630 (11) | 85,630 (11) | 100 | 6 | 7 |
| Dec-11 | 101,611 | 79,770 (14) | 79,770 (14) | 100 | 10 | 13 |
| Jan-12 | 103,427 | 87,296 (18) | 87,264 (17) | 99.963 | 11 | 13 |
| Feb-12 | 100,802 | 83,832 (13) | 83,817 (11) | 99.982 | 7 | 8 |
| Apr-12 | 107,125 | 92,808 (17) | 92,806 (16) | 99.998 | 11 | 12 |
| Jun-12 | 110,616 | 94,899 (22) | 94,734 (18) | 99.826 | 10 | 11 |
| Jul-12 | 114,277 | 105,637 (7) | 105,610 (1) | 99.974 | 1 | 1 |
| Aug-12 | 111,862 | 102,504 (6) | 102,494 (3) | 99.990 | 3 | 3 |
| Nov-12 | 116,199 | 100,154 (11) | 100,154 (11) | 100 | 5 | 5 |
| Dec-12 | 118,027 | 97,677 (12) | 97,677 (12) | 100 | 5 | 5 |
| 2011–2012 | 106,328 (±11,680) | 91,693 (12) loss of 14% ± 4.1% | 90,137 (10) | 98.346 | 7 | 8 |
| Mar-17 | 204,856 | 179,282 (37) | 179,137 (35) | 99.919 | 13 | 7 |
| Apr-17 | 205,429 | 187,007 (29) | 143,597 (21) | 76.787 | 5 | 3 |
| May-17 | 160,517 | 119,872 (79) | 119,824 (76) | 99.960 | 16 | 13 |
| Jun-17 | 175,947 | 150,860 (33) | 148,737 (25) | 98.593 | 10 | 7 |
| Jul-17 | 155,278 | 128,309 (44) | 128,184 (42) | 99.903 | 11 | 9 |
| Aug-17 | 133,853 | 118,825 (25) | 118,811 (22) | 99.988 | 11 | 9 |
| Sep-17 | 179,041 | 165,549 (19) | 162,832 (11) | 98.359 | 8 | 5 |
| Oct-17 | 205,001 | 179,162 (27) | 179,156 (26) | 99.997 | 14 | 8 |
| Nov-17 | 300,971 | 259,183 (54) | 258,949 (49) | 99.910 | 14 | 5 |
| Dec-17 | 132,716 | 111,404 (24) | 110,687 (20) | 99.356 | 10 | 9 |
| Jan-18 | 158,591 | 130,304 (33) | 130,300 (31) | 99.997 | 7 | 5 |
| Feb-18 | 136,157 | 105,016 (28) | 105,012 (27) | 99.996 | 10 | 10 |
| 2017–2018 | 179,030 (± 46,968) | 152,898 (36) loss of 15% ± 5.3% | 148,769 (32) | 97.730 | 11 | 8 |
| Type | More Recent Reported Isolates | Year/Period | Location | Source | Accession Number | Reference |
|---|---|---|---|---|---|---|
| CVA2 | Just mentioned a | 1991–1998 | Unknown | AM | Unavailable | [23] |
| CVA4 | Unknown | --- | --- | --- | --- | --- |
| CVA6 | CVA6/BSAS/Arg001/2018 | 2018 | Buenos Aires | HFMD | MK867799 | [39] |
| CVA8 | Unknown | --- | --- | --- | --- | --- |
| CVA10 | Unknown | --- | --- | --- | --- | --- |
| CVA12 | Unknown | --- | --- | --- | --- | --- |
| CVA16 | 707F99 | Unknown | Unknown | Unknown | AF290092 | [11] |
| EVA71 | Just mentioned a | 1991–1998 | Unknown | AM & AFP | Unavailable | [23] |
| EVA76 | Unknown | --- | --- | --- | --- | --- |
| EVA90 | Unknown | --- | --- | --- | --- | --- |
| CVA9 | Cba_Mar_2014_2 | 2014 | Córdoba | Wastewater | MK435332 | [21] |
| CVB1 | Just mentioned a | 1991–1998 | Unknown | AFP | Unavailable | [23] |
| CVB2 | AR1530-3-03 | 2003 | Buenos Aires | Neurological disease | DQ282630 | [22] |
| CVB3 | AR1450-49-02 | 2002 | Buenos Aires | Neurological disease | DQ246722 | [22] |
| CVB4 | Cba_Mar_2010 | 2010 | Córdoba | Wastewater | MK435329 | [21] |
| CVB5 | AR875-48-01 | 2001 | Buenos Aires | Neurological disease | DQ246729 | [22] |
| E2 | AR1095-20-02 | 2002 | Buenos Aires | Neurological disease | DQ282634 | [22] |
| E3 | Unknown | --- | --- | --- | --- | --- |
| E5 | AR1617-8-03 | 2003 | Buenos Aires | Neurological disease | DQ282627 | [22] |
| E6 | Cba_Feb_2014_1 | 2014 | Córdoba | Wastewater | MK435318 | [21] |
| E9 | AR1669_12_03 | 2003 | Buenos Aires | Neurological disease | DQ246752 | [22] |
| E11 | Just mentioned a | 2005–2006 | Buenos Aires | Surface water | Unavailable | [18] |
| E13 | Cba_May_2013_1 | 2013 | Córdoba | Wastewater | MK435334 | [21] |
| E14 | Cba_Set_2014 | 2014 | Córdoba | Wastewater | MK435346 | [21] |
| E15 | Unknown | --- | --- | --- | --- | --- |
| E16 | Cba_Jun_2013 | 2013 | Córdoba | Wastewater | MK435326 | [21] |
| E17 | 875NE99 | Unknown | Unknown | Unknown | AF290905 | [11] |
| E18 | AR2109-44-03 | 2003 | Buenos Aires | Neurological disease | DQ246765 | [22] |
| E30 | CHA468-ARG12 | 2012 | Chaco | AM | MK410196 | [13] |
| E32 | Unknown | --- | --- | --- | --- | --- |
| CVA1 | Unknown | --- | --- | --- | --- | --- |
| CVA11 | Unknown | --- | --- | --- | --- | --- |
| CVA13 | ARG98-10613/ARG98-10614 | 1998 | Unknown | Unknown | DQ995636 | [40] |
| CVA19 | Unknown | --- | --- | --- | --- | --- |
| CVA22 | Unknown | --- | --- | --- | --- | --- |
| CVA24 | Cba_Aug_2013/Cba_Aug_2014 | 2013/2014 | Córdoba | Wastewater | MK435313 | [21] |
| EVC99 | Unknown | --- | --- | --- | --- | --- |
| EVC116 | Unknown | --- | --- | --- | --- | --- |
| Aseptic Meningitis Outbreak Buenos Aires ᵃ (Oct 2011–Mar 2012) | Two Aseptic Meningitis Sporadic Cases Córdoba (Mar 2012) | Aseptic Meningitis Outbreak Chaco ᵃ (Feb 2012–Apr 2012) | Other Echovirus 30 Strains Detected in 1998–2008 in Argentina | |
|---|---|---|---|---|
| E30/COR/ARG/Dec_2011 | 95.9–99.3% | 99% | 97.8–99.3% | 87.9–92.2% |
| E30/COR/ARG/Jan_2012 | 97.2–98.1% | 97.80% | 97.2–98.1% | 87.6–91.9% |
| E30/COR/ARG/Feb_2012 | 97.5–98.4% | 98.10% | 97.5–98.4% | 87.3–92.2% |
| E30/COR/ARG/Feb_2012 | 95.6–98.1% | 97.80% | 97.2–98.4% | 87.3–91.9% |
| E30/COR/ARG/Jun_2012 | 96.2–99.6% | 100% | 98.1–99.6% | 87.9–92.2% |
| E30/COR/ARG/Mar_2017 | 86.4–87.3% | 87.30% | 86.1–87.3% | 87.9–92.2% |
| E30/COR/ARG/Mar_2017 | 83.3–84.5% | 84.50% | 83.3–84.5% | 85.1–91.0% |
| E30/COR/ARG/Mar_2017 | 84.2–85.8% | 85.10% | 84.5–85.8% | 87.6–91.9% |
| E30/COR/ARG/Mar_2017 | 85.8–87.3% | 86.70% | 86.7–87.6% | 86.4–95.6% |
| E30/COR/ARG/Mar_2017 | 84.5–86.7% | 86.10% | 85.4–86.7% | 86.1–92.5% |
| E30/COR/ARG/Mar_2017 | 85.4–86.4% | 86.40% | 85.1–86.4% | 87.0–93.5% |
| E30/COR/ARG/Apr_2017 | 85.4–87.0% | 86.40% | 86.4–87.3% | 87.3–95.3% |
| E30/COR/ARG/May_2017 | 86.1–87.6% | 87.00% | 87.0–87.9% | 86.7–95.3% |
| E30/COR/ARG/Jun_2017 | 85.8–87.3% | 86.70% | 86.7–87.6% | 87.0–95.6% |
| E30/COR/ARG/Jul_2017 | 86.4–87.9% | 87.30% | 87.3–88.2% | 87.6–95.3% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lizasoain, A.; Mir, D.; Masachessi, G.; Farías, A.; Rodríguez-Osorio, N.; Victoria, M.; Nates, S.; Colina, R. Environmental Surveillance through Next-Generation Sequencing to Unveil the Diversity of Human Enteroviruses beyond the Reported Clinical Cases. Viruses 2021, 13, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010120
Lizasoain A, Mir D, Masachessi G, Farías A, Rodríguez-Osorio N, Victoria M, Nates S, Colina R. Environmental Surveillance through Next-Generation Sequencing to Unveil the Diversity of Human Enteroviruses beyond the Reported Clinical Cases. Viruses. 2021; 13(1):120. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010120
Chicago/Turabian StyleLizasoain, Andrés, Daiana Mir, Gisella Masachessi, Adrián Farías, Nélida Rodríguez-Osorio, Matías Victoria, Silvia Nates, and Rodney Colina. 2021. "Environmental Surveillance through Next-Generation Sequencing to Unveil the Diversity of Human Enteroviruses beyond the Reported Clinical Cases" Viruses 13, no. 1: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/v13010120