
In Vitro Characterization and In Vivo Efficacy Assessment in Galleria mellonella Larvae of Newly Isolated Bacteriophages against Escherichia coli K1

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phage Isolation and Propagation
2.3. Host Range and Efficiency of Plating
2.4. Transmission Electron Microscopy
2.5. DNA Sequencing and Phage Genome Annotation
2.6. Adsorption Time and Low MOI Phage Kinetics
2.7. Temperature and pH Stability
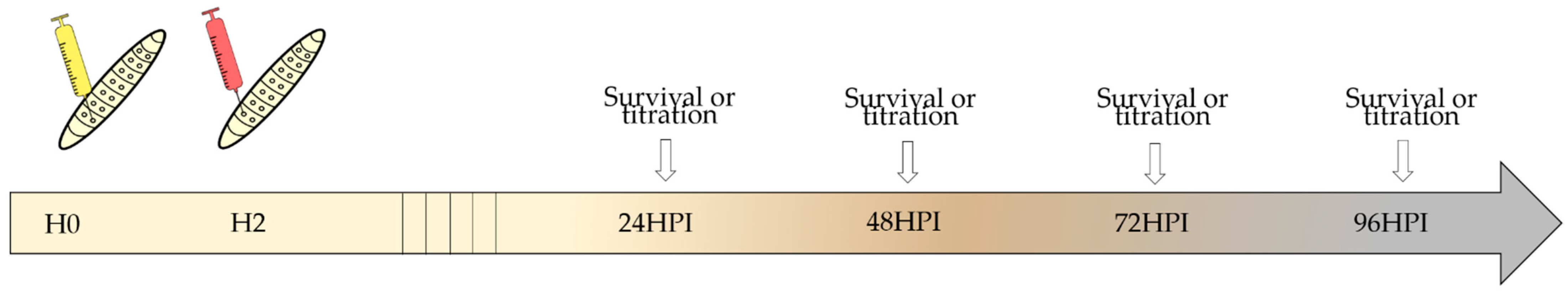
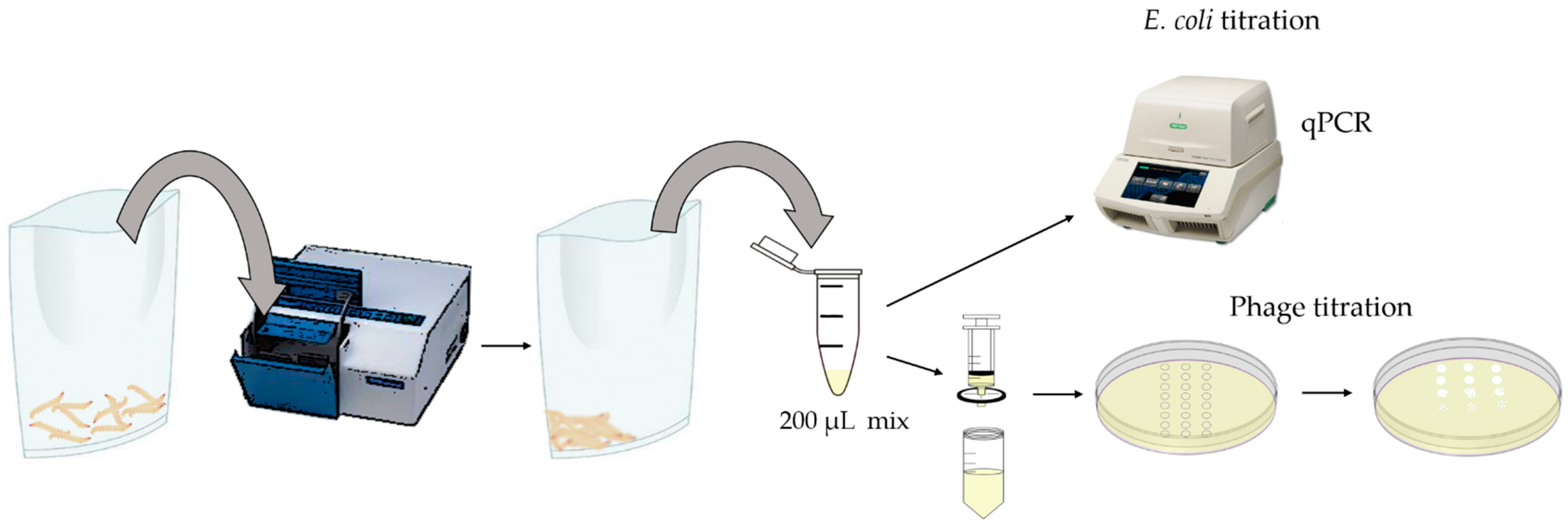
2.8. In Vivo Assay: Galleria Mellonella
3. Results
3.1. Phage Isolation
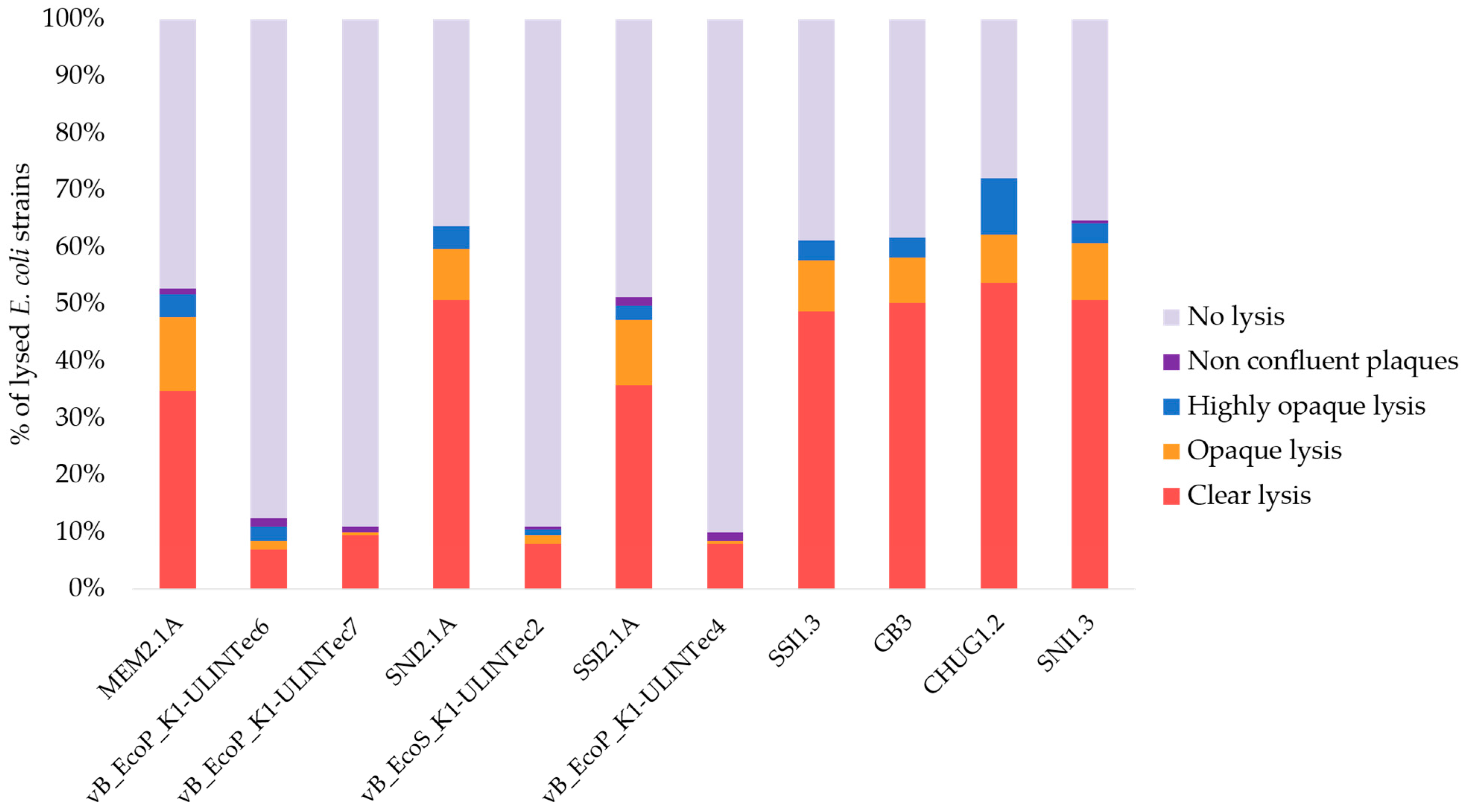
3.2. Host Range and Efficiency of Plating
3.3. Transmission Electron Microscopy
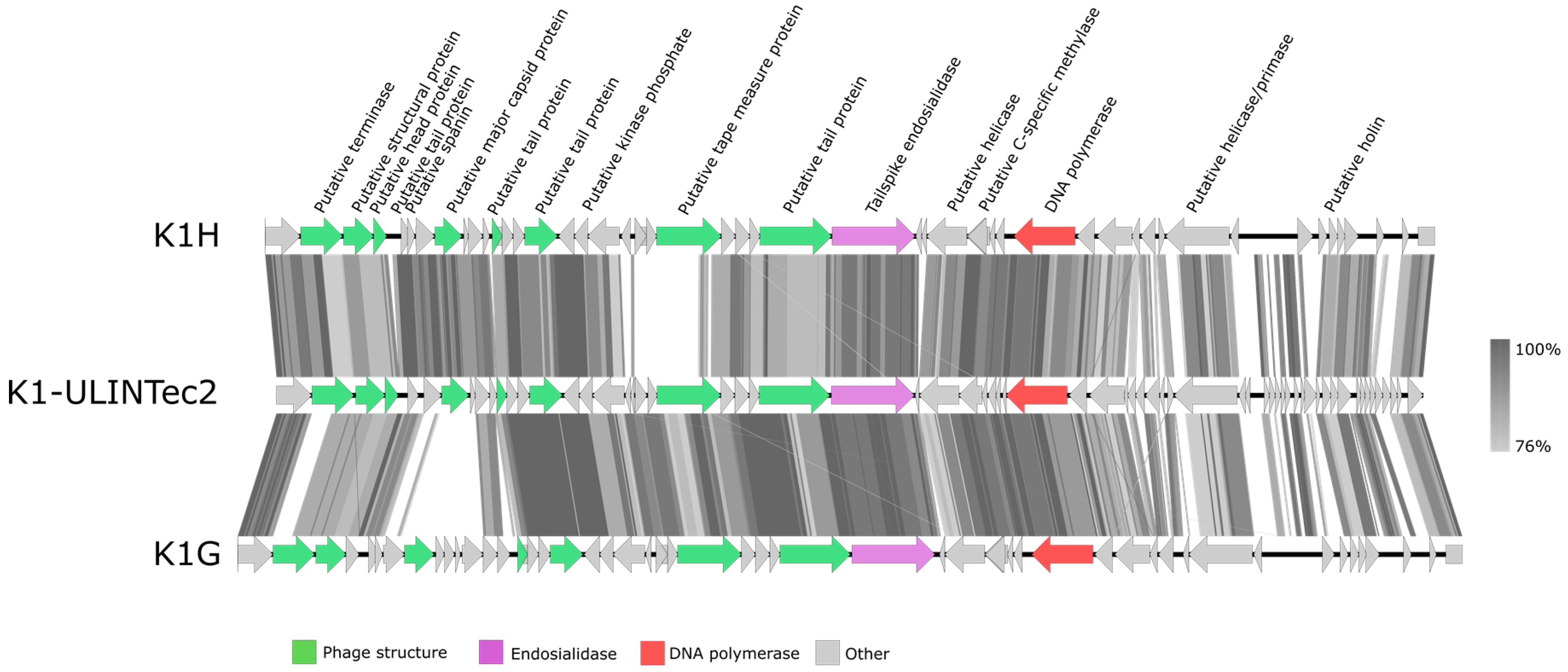
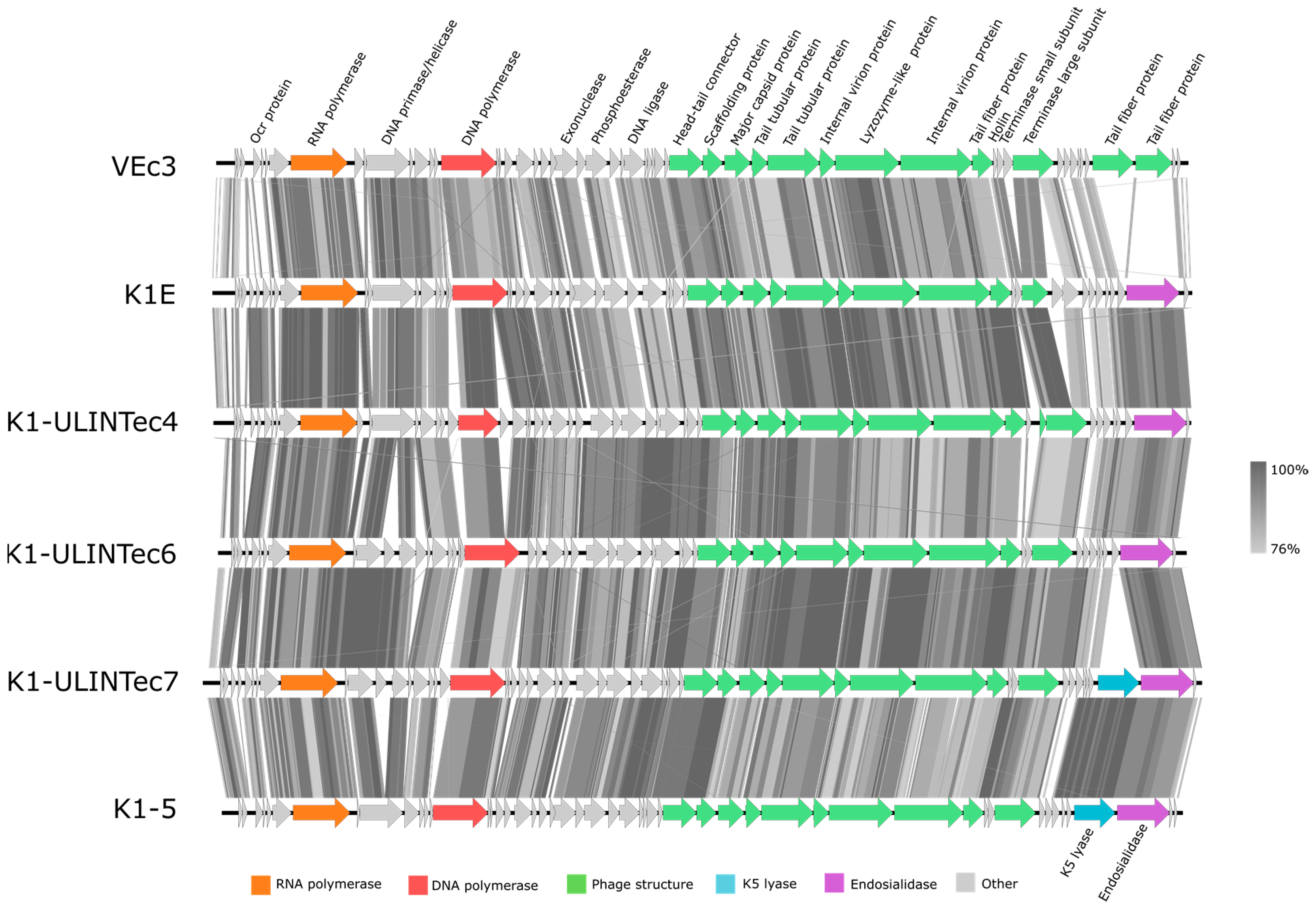
3.4. DNA Sequencing and Phage Genome Annotation
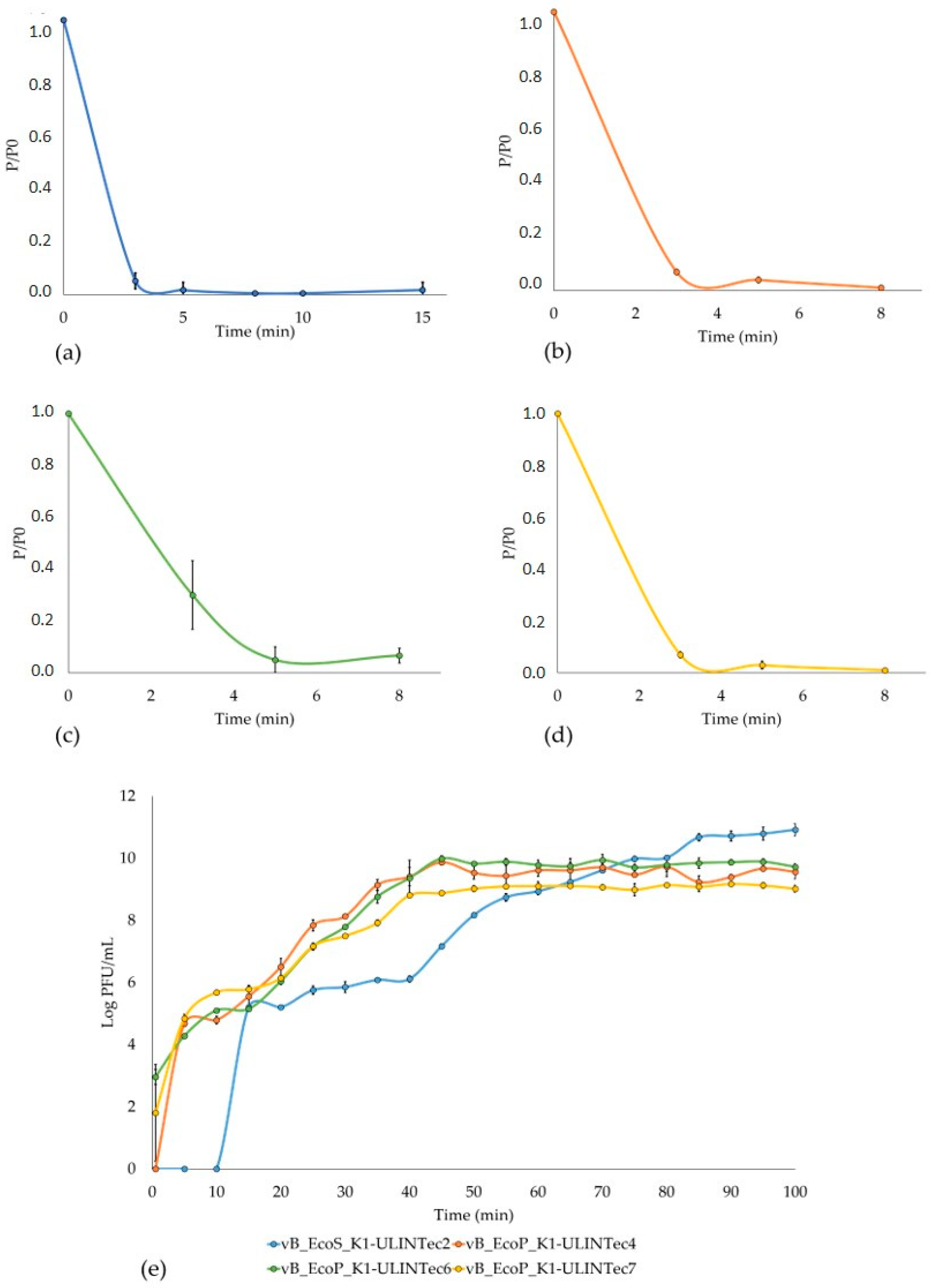
3.5. Adsorption Time and Low MOI Phage Kinetics
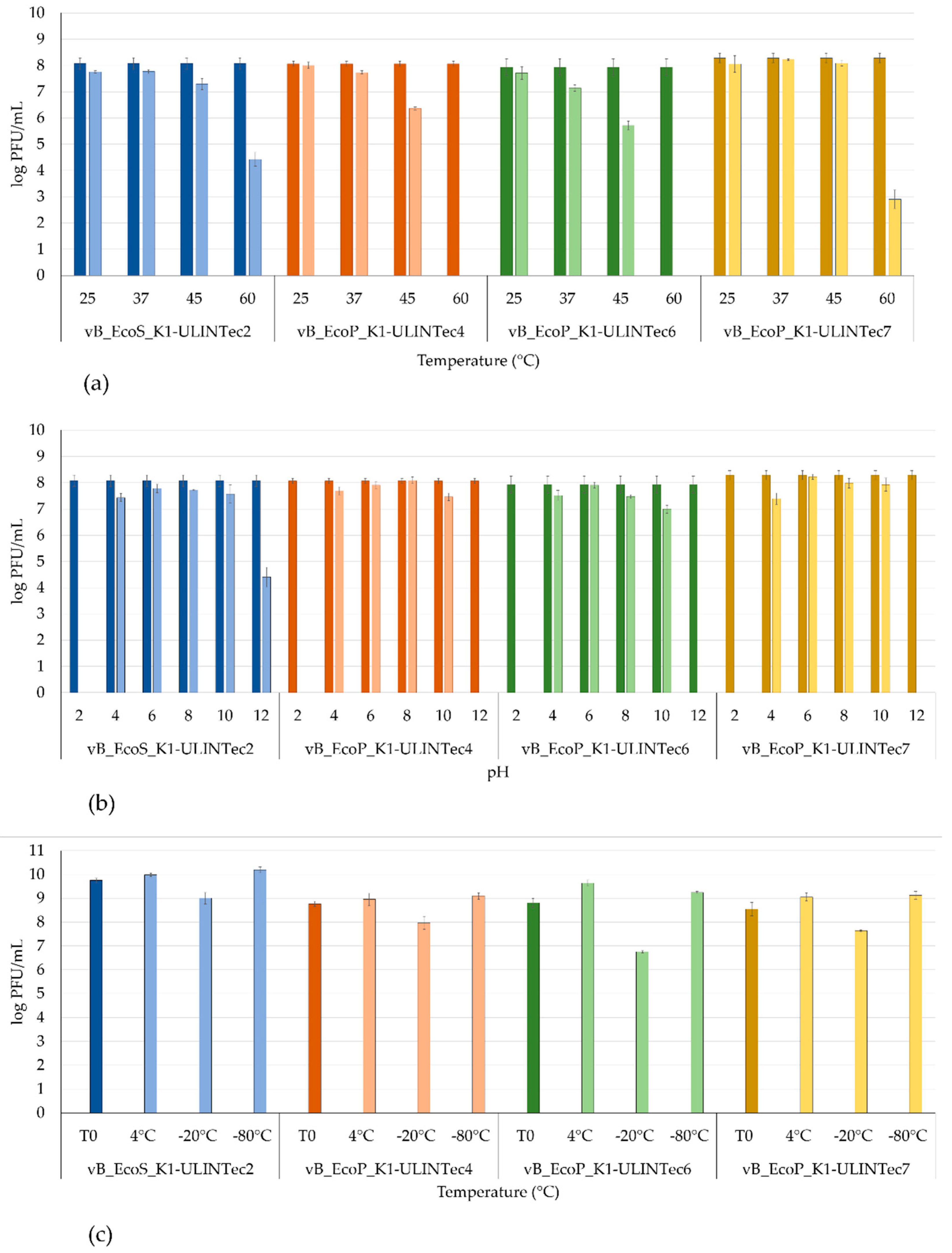
3.6. Temperature and pH Stability
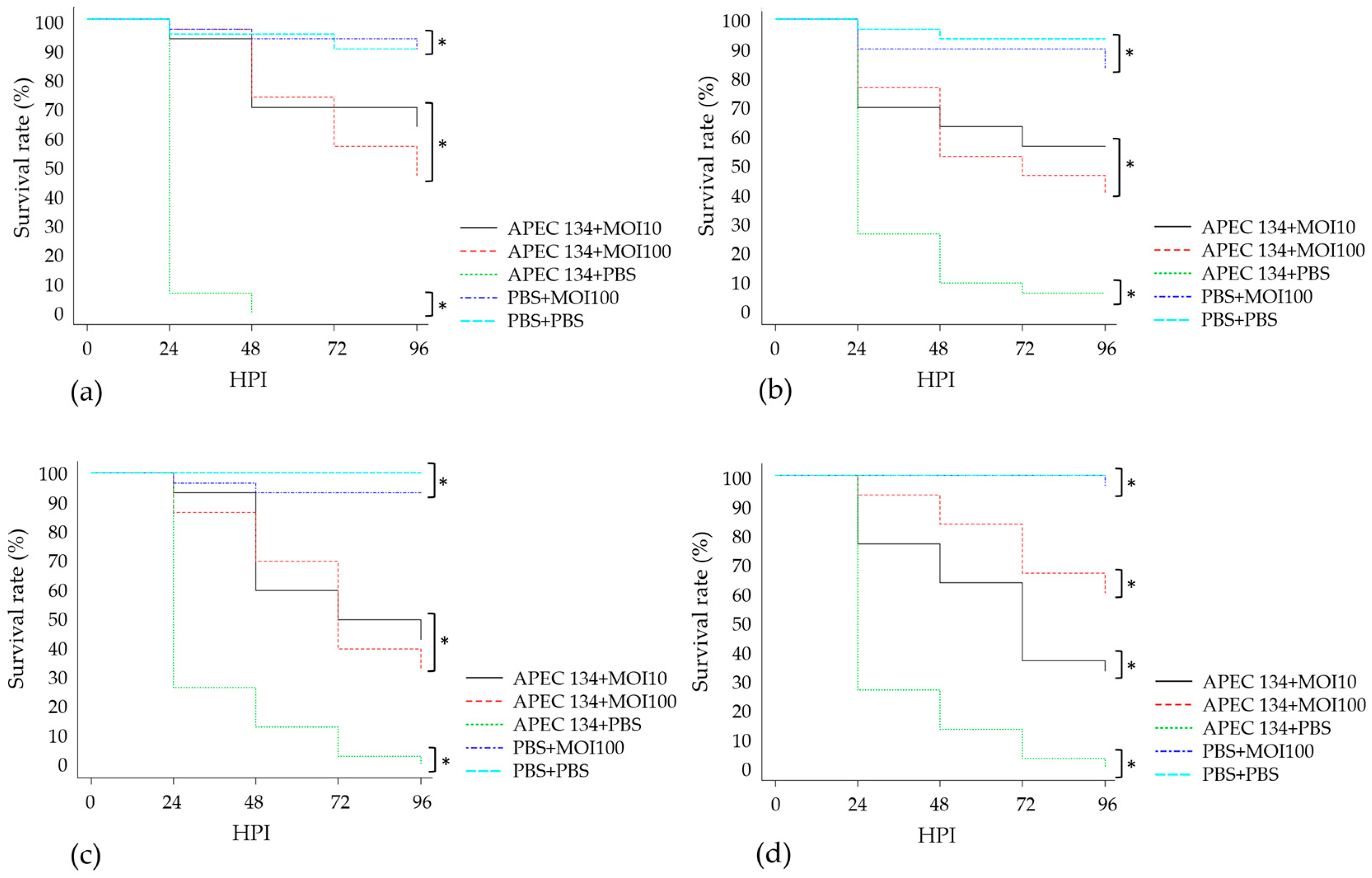
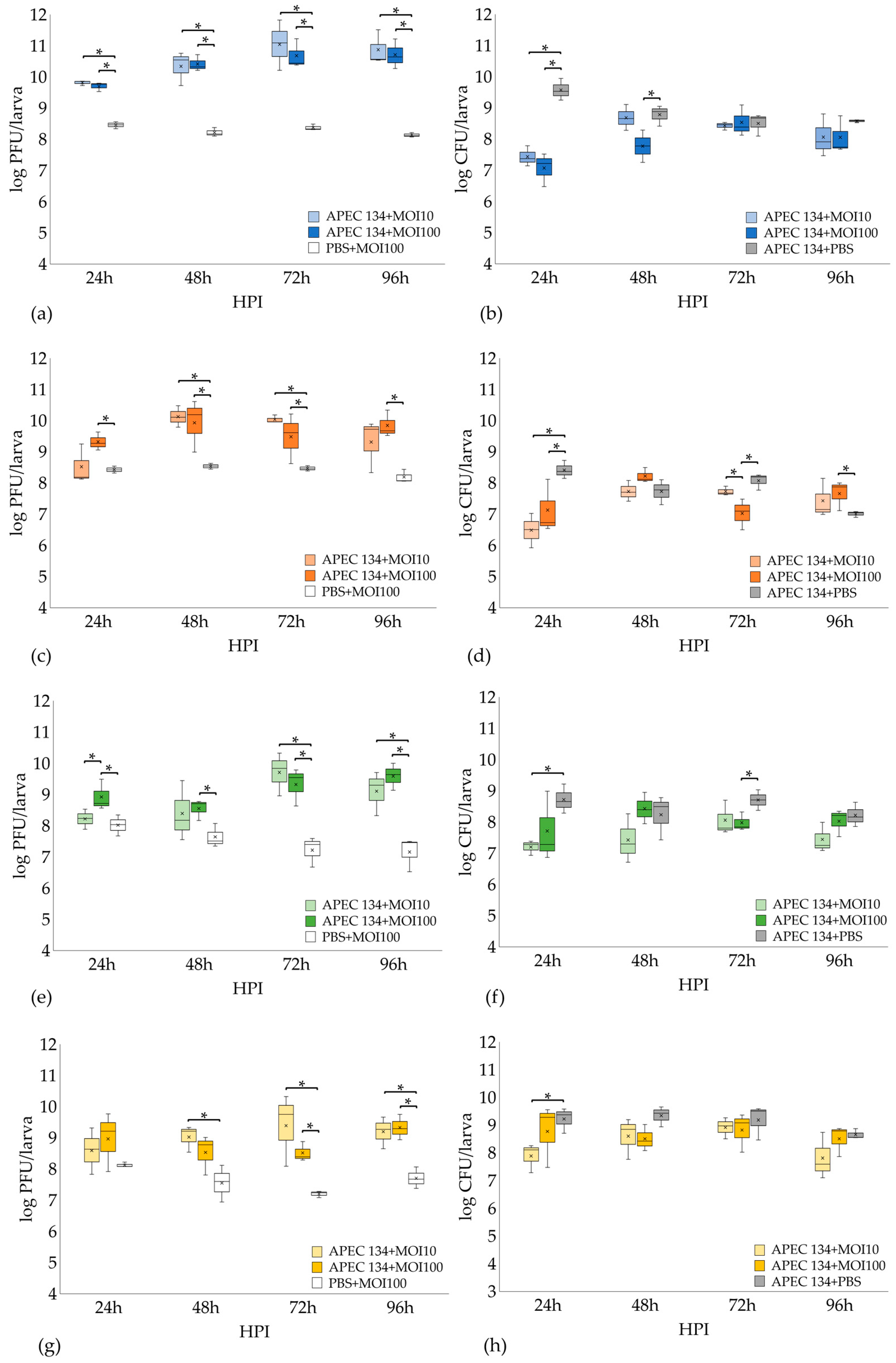
3.7. In Vivo Assay: Galleria Mellonella
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dale, A.P.; Woodford, N. Extra-Intestinal Pathogenic Escherichia coli (ExPEC): Disease, Carriage and Clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Delavari, P.; O’Bryan, T.T. Escherichia coli O18:K1:H7 Isolates from Patients with Acute Cystitis and Neonatal Meningitis Exhibit Common Phylogenetic Origins and Virulence Factor Profiles. J. Infect. Dis. 2001, 183, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, K.A.; Anderson-Berry, A.L.; Delair, S.F.; Davies, H.D. Early-Onset Neonatal Sepsis. Clin. Microbiol. Rev. 2014, 27, 21–47. [Google Scholar] [CrossRef] [Green Version]
- King, J.E.; Aal Owaif, H.A.; Jia, J.; Roberts, I.S. Phenotypic Heterogeneity in Expression of the K1 Polysaccharide Capsule of Uropathogenic Escherichia coli and Downregulation of the Capsule Genes during Growth in Urine. Infect. Immun. 2015, 83, 2605–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.S. Human Meningitis-Associated Escherichia coli. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.J.; Elliott, S.J.; Cello, F.D.; Stins, M.F.; Kim, K.S. The K1 Capsule Modulates Trafficking of E. coli-Containing Vacuoles and Enhances Intracellular Bacterial Survival in Human Brain Microvascular Endothelial Cells. Cell. Microbiol. 2003, 5, 245–252. [Google Scholar] [CrossRef]
- Xie, Y.; Kim, K.J.; Kim, K.S. Current Concepts on Escherichia coli K1 Translocation of the Blood–Brain Barrier. FEMS Immunol. Med. Microbiol. 2004, 42, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Sivanandan, S.; Soraisham, A.S.; Swarnam, K. Choice and Duration of Antimicrobial Therapy for Neonatal Sepsis and Meningitis. Int. J. Pediatr. 2011, 2011, 712150. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Dufendach, K.R.; Wellons, J.C.; Kuba, M.G.; Nickols, H.H.; Gómez-Duarte, O.G.; Wynn, J.L. Lethal Neonatal Meningoencephalitis Caused by Multi-Drug Resistant, Highly Virulent Escherichia coli. Infect. Dis. Lond. Engl. 2016, 48, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Jin, Y.; Duan, Y.; He, M.; Lin, Z.; Lin, J. Multi-Drug Resistant Escherichia Coli Causing Early-Onset Neonatal Sepsis—A Single Center Experience from China. Infect. Drug Resist. 2019, 12, 3695–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, T.A.; Munoz, F.M.; Troisi, C.L.; Nolan, M.S.; Hasbun, R.; Brown, E.L.; Murray, K.O. The Epidemiology of Meningitis in Infants under 90 Days of Age in a Large Pediatric Hospital. Microorganisms 2021, 9, 526. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Ding, Y.; Yao, K.; Gao, W.; Wang, Y. Antimicrobial Resistance Analysis of Clinical Escherichia coli Isolates in Neonatal Ward. Front. Pediatr. 2021, 9, 670470. [Google Scholar] [CrossRef]
- Johnson, J.R.; Stell, A.L.; Delavari, P. Canine Feces as a Reservoir of Extraintestinal Pathogenic Escherichia coli. Infect. Immun. 2001, 69, 1306–1314. [Google Scholar] [CrossRef] [Green Version]
- Moulin-Schouleur, M.; Schouler, C.; Tailliez, P.; Kao, M.R.; Brée, A.; Germon, P.; Oswald, E.; Mainil, J.; Blanco, M.; Blanco, J. Common Virulence Factors and Genetic Relationships between O18:K1:H7 Escherichia coli Isolates of Human and Avian Origin. J. Clin. Microbiol. 2006, 44, 3484–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassé, F.N.; Fairbrother, J.M.; Dubuc, J. Relationship between Escherichia coli Virulence Factors and Postpartum Metritis in Dairy Cows. J. Dairy Sci. 2016, 99, 4656–4667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mageiros, L.; Méric, G.; Bayliss, S.C.; Pensar, J.; Pascoe, B.; Mourkas, E.; Calland, J.K.; Yahara, K.; Murray, S.; Wilkinson, T.S.; et al. Genome Evolution and the Emergence of Pathogenicity in Avian Escherichia coli. Nat. Commun. 2021, 12, 765. [Google Scholar] [CrossRef]
- Sun, M.; Gao, X.; Zhao, K.; Ma, J.; Yao, H.; Pan, Z. Insight into the Virulence Related Secretion Systems, Fimbriae, and Toxins in O2:K1 Escherichia coli Isolated from Bovine Mastitis. Front. Vet. Sci. 2021, 8, 18. [Google Scholar] [CrossRef]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathog. Basel Switz. 2021, 10, 467. [Google Scholar] [CrossRef]
- Zhuge, X.; Zhou, Z.; Jiang, M.; Wang, Z.; Sun, Y.; Tang, F.; Xue, F.; Ren, J.; Dai, J. Chicken-Source Escherichia coli within Phylogroup F Shares Virulence Genotypes and Is Closely Related to Extraintestinal Pathogenic E. coli Causing Human Infections. Transbound. Emerg. Dis. 2021, 68, 880–895. [Google Scholar] [CrossRef]
- Tivendale, K.A.; Logue, C.M.; Kariyawasam, S.; Jordan, D.; Hussein, A.; Li, G.; Wannemuehler, Y.; Nolan, L.K. Avian-Pathogenic Escherichia coli Strains Are Similar to Neonatal Meningitis E. coli Strains and Are Able to Cause Meningitis in the Rat Model of Human Disease. Infect. Immun. 2010, 78, 3412–3419. [Google Scholar] [CrossRef] [Green Version]
- Mellata, M.; Johnson, J.R.; Curtiss, R. Escherichia coli Isolates from Commercial Chicken Meat and Eggs Cause Sepsis, Meningitis and Urinary Tract Infection in Rodent Models of Human Infections. Zoonoses Public Health 2018, 65, 103–113. [Google Scholar] [CrossRef]
- Liu, C.M.; Stegger, M.; Aziz, M.; Johnson, T.J.; Waits, K.; Nordstrom, L.; Gauld, L.; Weaver, B.; Rolland, D.; Statham, S.; et al. Escherichia coli ST131-H22 as a Foodborne Uropathogen. mBio 2018, 9, e00470-18. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, S.L.; Stegger, M.; Kudirkiene, E.; Lilje, B.; Poulsen, L.L.; Ronco, T.; Pires Dos Santos, T.; Kiil, K.; Bisgaard, M.; Pedersen, K.; et al. Diversity and Population Overlap between Avian and Human Escherichia coli Belonging to Sequence Type 95. mSphere 2019, 4, e00333-18. [Google Scholar] [CrossRef] [Green Version]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage Therapy: A Renewed Approach to Combat Antibiotic-Resistant Bacteria. Cell Host Microbe 2019, 25, 219–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Gross, R.J.; Cheasty, T.; Rowe, B. Isolation of Bacteriophages Specific for the K1 Polysaccharide Antigen of Escherichia coli. J. Clin. Microbiol. 1977, 6, 548–550. [Google Scholar] [CrossRef]
- Smith, H.W.; Huggins, M.B. Successful Treatment of Experimental Escherichia coli Infections in Mice Using Phage: Its General Superiority over Antibiotics. J. Gen. Microbiol. 1982, 128, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Scholl, D.; Rogers, S.; Adhya, S.; Merril, C.R. Bacteriophage K1-5 Encodes Two Different Tail Fiber Proteins, Allowing It to Infect and Replicate on Both K1 and K5 Strains of Escherichia coli. J. Virol. 2001, 75, 2509–2515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bull, J.J.; Levin, B.R.; DeRouin, T.; Walker, N.; Bloch, C.A. Dynamics of Success and Failure in Phage and Antibiotic Therapy in Experimental Infections. BMC Microbiol. 2002, 2, 35. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Otto, G.; Molineux, I.J. In Vivo Growth Rates Are Poorly Correlated with Phage Therapy Success in a Mouse Infection Model. Antimicrob. Agents Chemother. 2012, 56, 949–954. [Google Scholar] [CrossRef] [Green Version]
- Desbois, A.P.; Coote, P.J. Utility of Greater Wax Moth Larva (Galleria Mellonella) for Evaluating the Toxicity and Efficacy of New Antimicrobial Agents. Adv. Appl. Microbiol. 2012, 78, 25–53. [Google Scholar] [CrossRef]
- Cutuli, M.A.; Petronio Petronio, G.; Vergalito, F.; Magnifico, I.; Pietrangelo, L.; Venditti, N.; Di Marco, R. Galleria Mellonella as a Consolidated In Vivo Model Hosts: New Developments in Antibacterial Strategies and Novel Drug Testing. Virulence 2019, 10, 527–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kling, J. Get a Look at Galleria. Lab Anim. 2020, 49, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Lavine, M.D.; Strand, M.R. Insect Hemocytes and Their Role in Immunity. Insect Immunol. 2002, 32, 25–47. [Google Scholar] [CrossRef]
- Casanova-Torres, Á.M.; Goodrich-Blair, H. Immune Signaling and Antimicrobial Peptide Expression in Lepidoptera. Insects 2013, 4, 320–338. [Google Scholar] [CrossRef] [Green Version]
- Wojda, I. Immunity of the Greater Wax Moth Galleria Mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Dongeun, Y. Two Novel Bacteriophages Improve Survival in Galleria Mellonella Infection and Mouse Acute Pneumonia Models Infected with Extensively Drug-Resistant Pseudomonas Aeruginosa. Appl. Environ. Microbiol. 2019, 85, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Thiry, D.; Passet, V.; Danis-Wlodarczyk, K.; Lood, C.; Wagemans, J.; De Sordi, L.; van Noort, V.; Dufour, N.; Debarbieux, L.; Mainil, J.G.; et al. New Bacteriophages against Emerging Lineages ST23 and ST258 of Klebsiella Pneumoniae and Efficacy Assessment in Galleria Mellonella Larvae. Viruses 2019, 11, 411. [Google Scholar] [CrossRef] [Green Version]
- Antoine, C.; Laforêt, F.; Blasdel, B.; Glonti, T.; Kutter, E.; Pirnay, J.P.; Mainil, J.; Delcenserie, V.; Thiry, D. Efficacy Assessment of PEV2 Phage on Galleria Mellonella Larvae Infected with a Pseudomonas Aeruginosa Dog Otitis Isolate. Res. Vet. Sci. 2021, 136, 598–601. [Google Scholar] [CrossRef]
- Ngassam-Tchamba, C.; Duprez, J.N.; Fergestad, M.; De Visscher, A.; L’Abee-Lund, T.; De Vliegher, S.; Wasteson, Y.; Touzain, F.; Blanchard, Y.; Lavigne, R.; et al. In Vitro and In Vivo Assessment of Phage Therapy against Staphylococcus Aureus Causing Bovine Mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 762–770. [Google Scholar] [CrossRef]
- Manohar, P.; Nachimuthu, R.; Lopes, B.S. The Therapeutic Potential of Bacteriophages Targeting Gram-Negative Bacteria Using Galleria Mellonella Infection Model. BMC Microbiol. 2018, 18, 97. [Google Scholar] [CrossRef] [Green Version]
- Van Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil. Methods Mol. Biol. Clifton NJ 2009, 501, 15–21. [Google Scholar] [CrossRef]
- Mast, J.; Demeestere, L. Electron Tomography of Negatively Stained Complex Viruses: Application in Their Diagnosis. Diagn. Pathol. 2009, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A Modular and Extensible Implementation of the RAST Algorithm for Building Custom Annotation Pipelines and Annotating Batches of Genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Chang, H.-Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro Protein Families and Domains Database: 20 Years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef]
- Meier, A.; Söding, J. Automatic Prediction of Protein 3D Structures by Probabilistic Multi-Template Homology Modeling. PLoS Comput. Biol. 2015, 11, e1004343. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A Genome Comparison Visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Vandenheuvel, D.; Kropinski, A.M.; Mast, J.; Vos, D.D.; Verbeken, G.; Noben, J.-P.; Lavigne, R.; Vaneechoutte, M.; Pirnay, J.-P. Characterization of Newly Isolated Lytic Bacteriophages Active against Acinetobacter Baumannii. PLoS ONE 2014, 9, e104853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Menéndez, E.; Fernández, L.; Gutiérrez, D.; Rodríguez, A.; Martínez, B.; García, P. Comparative Analysis of Different Preservation Techniques for the Storage of Staphylococcus Phages Aimed for the Industrial Development of Phage-Based Antimicrobial Products. PLoS ONE 2018, 13, e0205728. [Google Scholar] [CrossRef]
- Using the R Commander. Available online: https://socialsciences.mcmaster.ca/jfox/Books/RCommander/ (accessed on 23 August 2021).
- Delannoy, S.; Beutin, L.; Mariani-Kurkdjian, P.; Fleiss, A.; Bonacorsi, S.; Fach, P. The Escherichia coli Serogroup O1 and O2 Lipopolysaccharides Are Encoded by Multiple O-Antigen Gene Clusters. Front. Cell. Infect. Microbiol. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bacchetti De Gregoris, T.; Aldred, N.; Clare, A.S.; Burgess, J.G. Improvement of Phylum- and Class-Specific Primers for Real-Time PCR Quantification of Bacterial Taxa. J. Microbiol. Methods 2011, 86, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at Its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Lysis from without. Bacteriophage 2011, 1, 46–49. [Google Scholar] [CrossRef]
- Bull, J.J.; Vimr, E.R.; Molineux, I.J. A Tale of Tails: Sialidase Is Key to Success in a Model of Phage Therapy against K1-Capsulated Escherichia coli. Virology 2010, 398, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baig, A.; Colom, J.; Barrow, P.; Schouler, C.; Moodley, A.; Lavigne, R.; Atterbury, R.; Atterbury, R.; Barrow, P.; Baig, A.; et al. Biology and Genomics of an Historic Therapeutic Escherichia coli Bacteriophage Collection. Front. Microbiol. 2017, 8, 1652. [Google Scholar] [CrossRef] [PubMed]
- Scholl, D.; Adhya, S.; Merril, C. Escherichia coli K1′s Capsule Is a Barrier to Bacteriophage T7. Appl. Environ. Microbiol. 2005, 71, 4872–4874. [Google Scholar] [CrossRef] [Green Version]
- Leiman, P.G.; Battisti, A.J.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; Gerardy-Schahn, R.; Scholl, D.; Molineux, I.J. The Structures of Bacteriophages K1E and K1-5 Explain Processive Degradation of Polysaccharide Capsules and Evolution of New Host Specificities. J. Mol. Biol. 2007, 371, 836–849. [Google Scholar] [CrossRef]
- Mushtaq, N.; Redpath, M.B.; Luzio, J.P.; Taylor, P.W. Prevention and Cure of Systemic Escherichia coli K1 Infection by Modification of the Bacterial Phenotype. Antimicrob. Agents Chemother. 2004, 48, 1503–1508. [Google Scholar] [CrossRef] [Green Version]
- Schneider, G.; Szentes, N.; Horváth, M.; Dorn, Á.; Cox, A.; Nagy, G.; Doffkay, Z.; Maróti, G.; Rákhely, G.; Kovács, T. Kinetics of Targeted Phage Rescue in a Mouse Model of Systemic Escherichia coli K1. BioMed Res. Int. 2018, 2018, 7569645. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | 1st Injection | 2nd Injection | |
|---|---|---|---|
| 1 | APEC 134 + phage 1 MOI 100 | APEC 134: 106 CFU/10 µL | Phage 1: 108 PFU/10 µL |
| 2 | APEC 134 + phage 1 MOI 10 | APEC 134: 106 CFU/10 µL | Phage 1: 107 PFU/10 µL |
| 3 | APEC 134 + PBS | APEC 134: 106 CFU/10 µL | PBS: 10 µL |
| 4 | PBS + phage 1 MOI 100 | PBS: 10 µL | Phage 1: 108 PFU/10 µL |
| 5 | PBS + PBS | PBS: 10 µL | PBS: 10 µL |
| Phages | |||||
|---|---|---|---|---|---|
| vB_EcoS_K1-ULINTec2 | vB_EcoP_K1-ULINTec4 | vB_EcoP_K1-ULINTec6 | vB_EcoP_K1-ULINTec7 | ||
| E. coli strains | 45 * | 1.000 | 1.000 | 1.000 | 1.000 |
| 30 * | 0.971 | 0.886 | 1.103 | 0.838 | |
| 79 * | NP | NP | / | NP | |
| 134 * | 0.765 | 1.143 | 1.205 | 2.137 | |
| 161 * | 0.535 | <0.001 | <0.001 | NP | |
| 486 * | 1.059 | NP | NP | NP | |
| 591 * | NP | 0.829 | 0.692 | 0.906 | |
| 592 * | NP | 1.000 | 0.405 | 1.086 | |
| 1073 * | NP | 0.120 | 0.024 | 0.044 | |
| 1343 * | NP | NP | 0.641 | NP | |
| 151.1 ** | / | / | / | 1.145 | |
| 143.1 * | NP | 1.143 | 0.487 | 0.350 | |
| O1 * | NP | 0.343 | NP | 0.256 | |
| O2 * | NP | 0.629 | / | 0.197 | |
| O7 * | NP | 0.943 | 1.923 | 0.855 | |
| O10 ** | / | / | / | 0.940 | |
| O12 ** | / | / | / | 1.453 | |
| O16 * | NP | 0.005 | NP | NP | |
| O45 * | NP | <0.001 | 0.564 | 2.821 | |
| O166 * | NP | 0.111 | NP | NP | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antoine, C.; Laforêt, F.; Blasdel, B.; Fall, A.; Duprez, J.-N.; Mainil, J.; Delcenserie, V.; Thiry, D. In Vitro Characterization and In Vivo Efficacy Assessment in Galleria mellonella Larvae of Newly Isolated Bacteriophages against Escherichia coli K1. Viruses 2021, 13, 2005. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102005
Antoine C, Laforêt F, Blasdel B, Fall A, Duprez J-N, Mainil J, Delcenserie V, Thiry D. In Vitro Characterization and In Vivo Efficacy Assessment in Galleria mellonella Larvae of Newly Isolated Bacteriophages against Escherichia coli K1. Viruses. 2021; 13(10):2005. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102005
Chicago/Turabian StyleAntoine, Céline, Fanny Laforêt, Bob Blasdel, Abdoulaye Fall, Jean-Noël Duprez, Jacques Mainil, Véronique Delcenserie, and Damien Thiry. 2021. "In Vitro Characterization and In Vivo Efficacy Assessment in Galleria mellonella Larvae of Newly Isolated Bacteriophages against Escherichia coli K1" Viruses 13, no. 10: 2005. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102005