
Feline Calicivirus Virulent Systemic Disease: Clinical Epidemiology, Analysis of Viral Isolates and In Vitro Efficacy of Novel Antivirals in Australian Outbreaks

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Data and Definitions
2.2. Sample Collection
2.3. Virus Isolation
2.4. Histopathology and Immunohistochemistry (IHC)
2.5. RNA Extraction RT-PCR
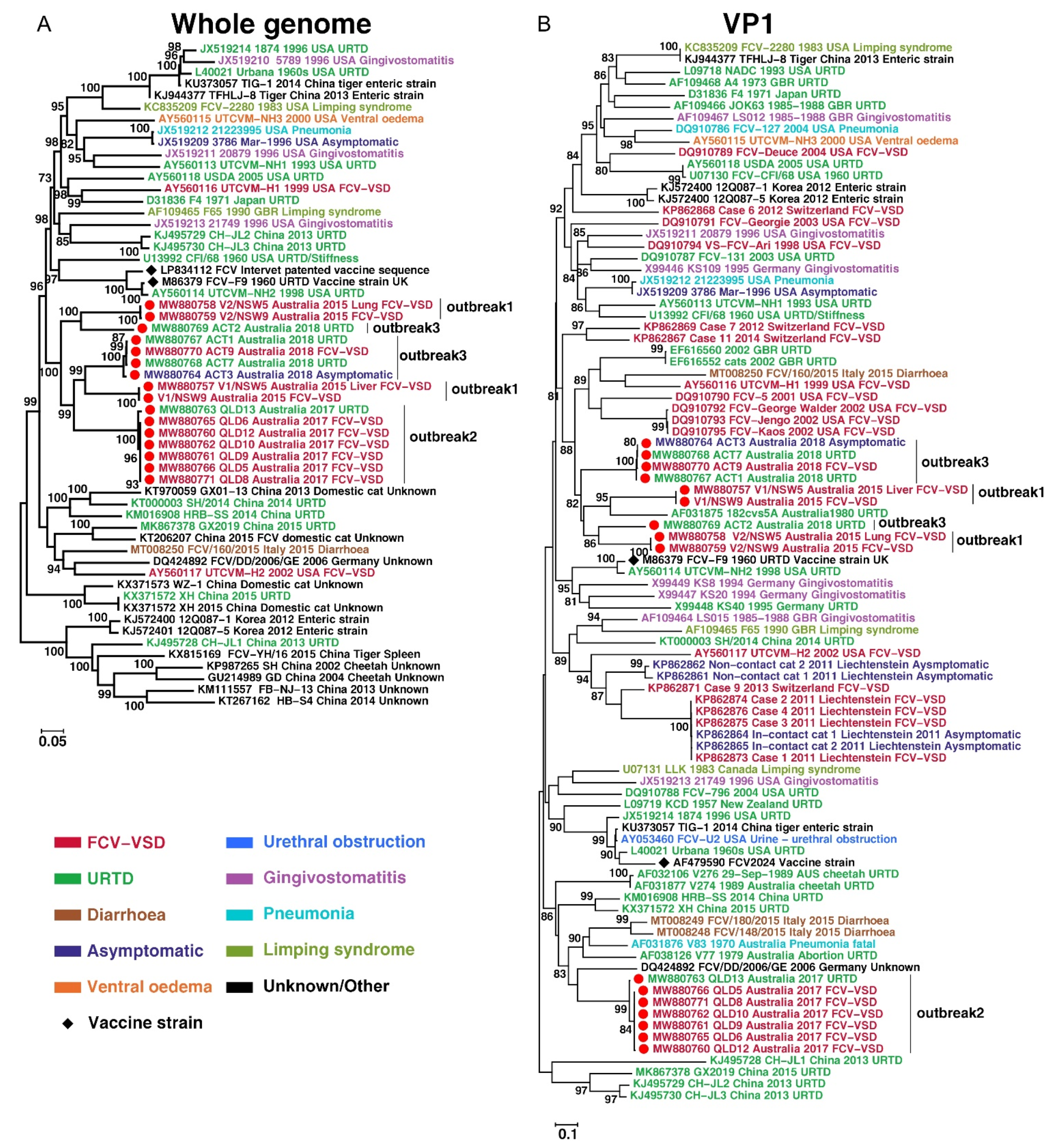
2.6. Bioinformatic and Phylogenetic Analyses
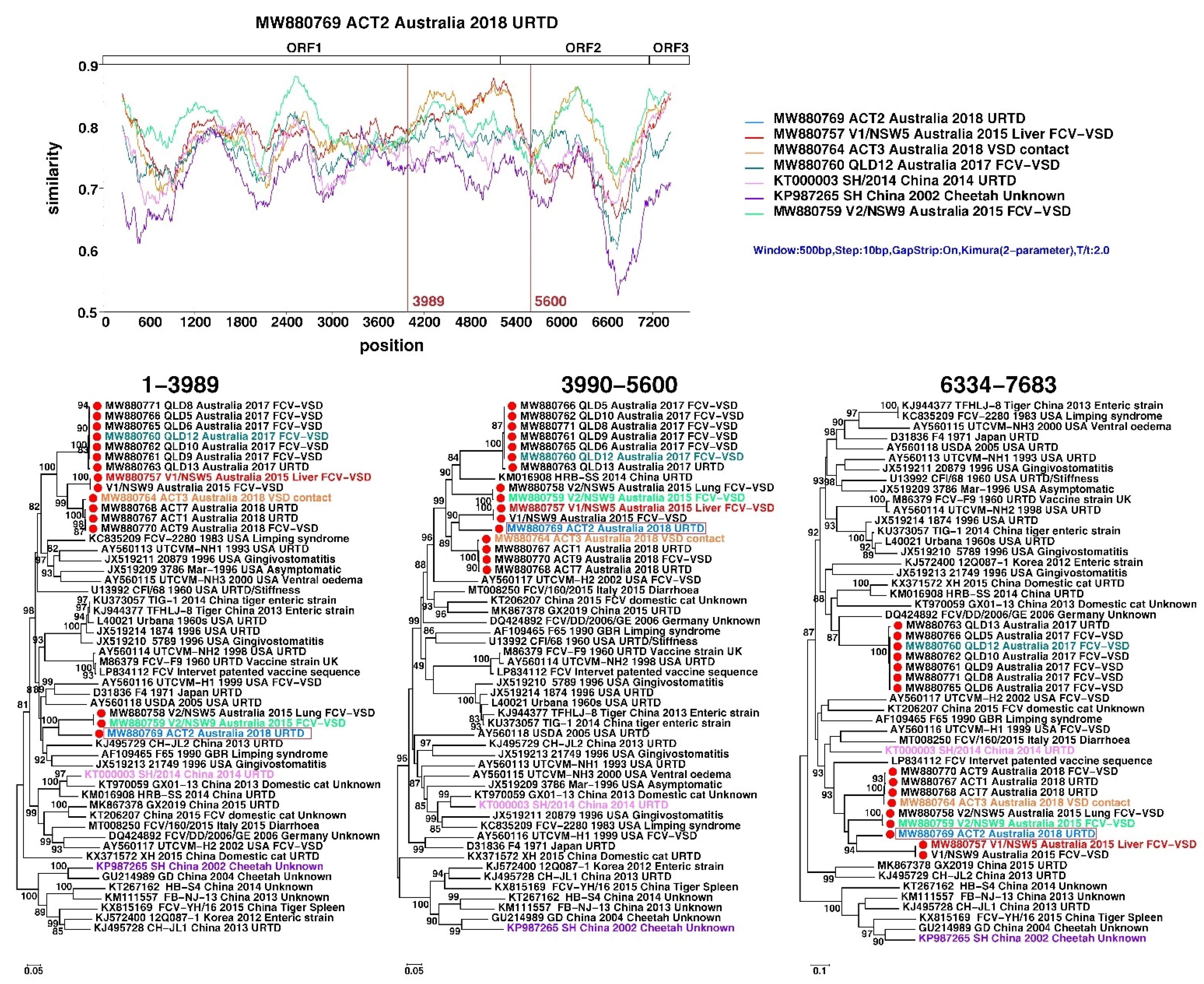
2.7. Recombination Analysis
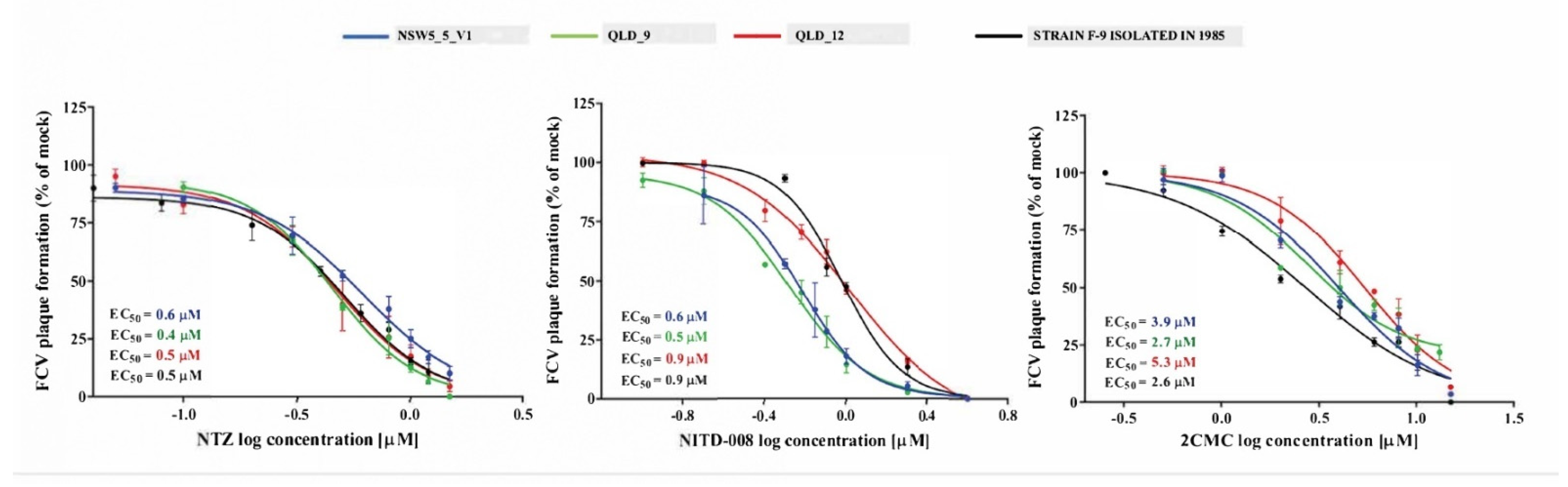
2.8. Evaluation of Antivirals in Plaque-Reduction Assays
3. Results
3.1. Clinical and Epidemiological Descriptions of Outbreaks
3.2. Histology and Immunohistochemistry
3.3. Virus Isolation and Genome Assembly
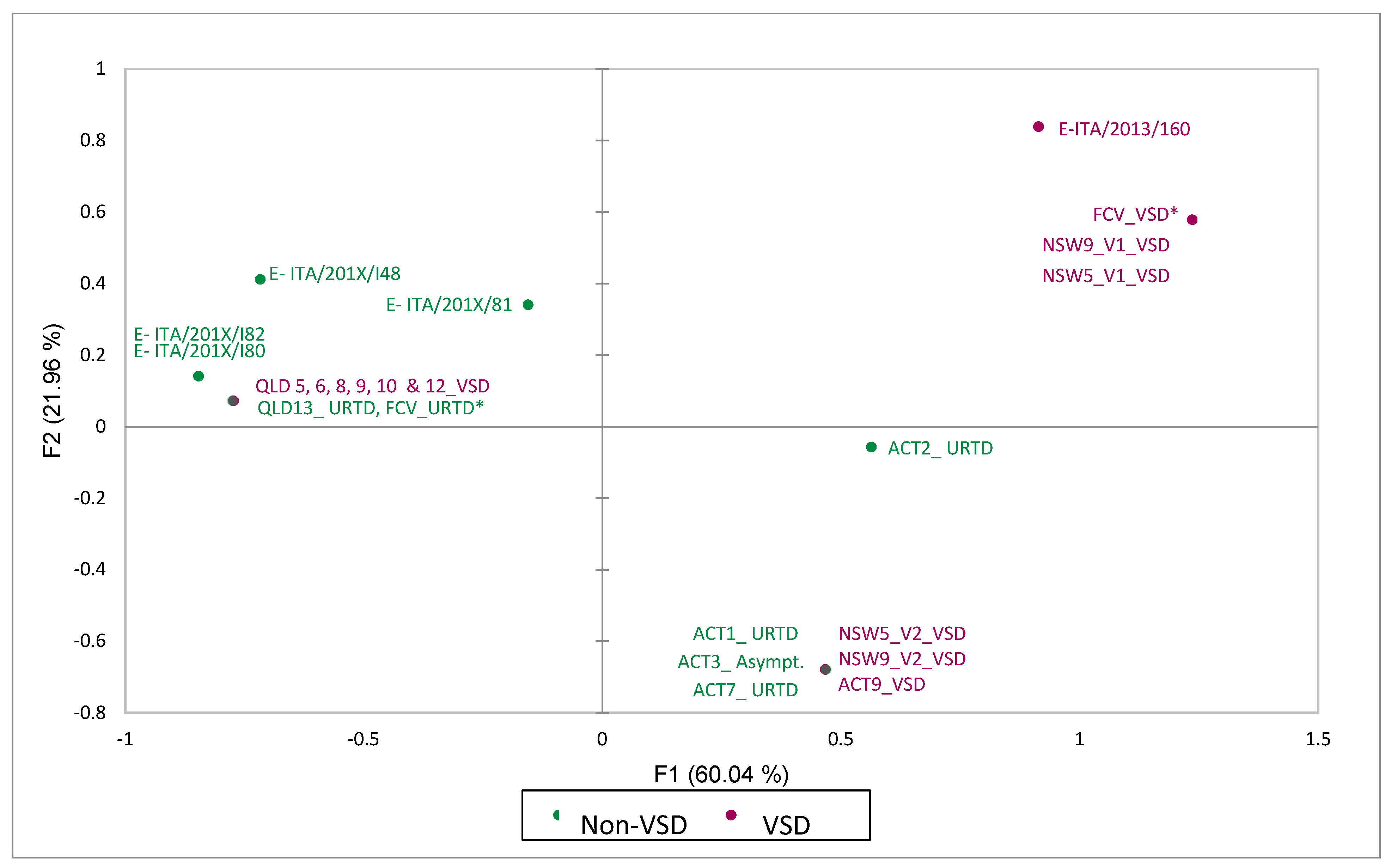
3.4. Metagenomic, Amino-Acid and Recombination Analyses
3.5. Antiviral Compound Efficacy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herbert, T.; Brierley, I.; Brown, T. Identification of a protein linked to the genomic and subgenomic mRNAs of feline calicivirus and its role in translation. J. Gen. Virol. 1997, 78, 1033–1040. [Google Scholar] [CrossRef]
- Neill, J.D.; Mengeling, W.L. Further characterization of the virus-specific RNAs in feline calicivirus infected cells. Virus Res. 1988, 11, 59–72. [Google Scholar] [CrossRef]
- Schaffer, F.; Ehresmann, D.; Fretz, M.; Soergel, M. A protein, VPg, covalently linked to 36S calicivirus RNA. J. Gen. Virol. 1980, 47, 215–220. [Google Scholar] [CrossRef]
- Mitra, T.; Sosnovtsev, S.V.; Green, K.Y. Mutagenesis of tyrosine 24 in the VPg protein is lethal for feline calicivirus. J. Virol. 2004, 78, 4931–4935. [Google Scholar] [CrossRef] [Green Version]
- Pesavento, P.A.; Chang, K.-O.; Parker, J.S. Molecular virology of feline calicivirus. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, N.C.; Elliott, J.B.; Glasgow, A.; Poland, A.; Keel, K. An isolated epizootic of hemorrhagic-like fever in cats caused by a novel and highly virulent strain of feline calicivirus. Vet. Microbiol. 2000, 73, 281–300. [Google Scholar] [CrossRef]
- Schorr-Evans, E.; Poland, A.; Johnson, W.; Pedersen, N.C. An epizootic of highly virulent feline calicivirus disease in a hospital setting in New England. J. Feline Med. Surg. 2003, 5, 217–226. [Google Scholar] [CrossRef]
- Reynolds, B.S.; Poulet, H.; Pingret, J.L.; Jas, D.; Brunet, S.; Lemeter, C.; Etievant, M.; Boucraut-Baralon, C. A nosocomial outbreak of feline calicivirus associated virulent systemic disease in France. J. Feline Med. Surg. 2009, 11, 633–644. [Google Scholar] [CrossRef]
- Battilani, M.; Vaccari, F.; Carelle, M.S.; Morandi, F.; Benazzi, C.; Kipar, A.; Dondi, F.; Scagliarini, A. Virulent feline calicivirus disease in a shelter in Italy: A case description. Res. Vet. Sci. 2013, 95, 283–290. [Google Scholar] [CrossRef]
- Prikhodko, V.G.; Sandoval-Jaime, C.; Abente, E.J.; Bok, K.; Parra, G.I.; Rogozin, I.B.; Ostlund, E.N.; Green, K.Y.; Sosnovtsev, S.V. Genetic characterization of feline calicivirus strains associated with varying disease manifestations during an outbreak season in Missouri (1995–1996). Virus Genes 2014, 48, 96–110. [Google Scholar] [CrossRef]
- Caringella, F.; Elia, G.; Decaro, N.; Martella, V.; Lanave, G.; Varello, K.; Catella, C.; Diakoudi, G.; Carelli, G.; Colaianni, M.L. Feline calicivirus infection in cats with virulent systemic disease, Italy. Res. Vet. Sci. 2019, 124, 46–51. [Google Scholar] [CrossRef]
- Radford, A.D.; Coyne, K.P.; Dawson, S.; Porter, C.J.; Gaskell, R.M. Feline calicivirus. Vet. Res. 2007, 38, 319–335. [Google Scholar] [CrossRef] [Green Version]
- Pesavento, P.A.; MacLachlan, N.J.; Dillard-Telm, L.; Grant, C.K.; Hurley, K.F. Pathologic, immunohistochemical, and electron microscopic findings in naturally occurring virulent systemic feline calicivirus infection in cats. Vet. Pathol. 2004, 41, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Fumian, T.M.; Tuipulotu, D.E.; Netzler, N.E.; Lun, J.H.; Russo, A.G.; Yan, G.J.; White, P.A. Potential therapeutic agents for feline calicivirus infection. Viruses 2018, 10, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enosi Tuipulotu, D.; Fumian, T.M.; Netzler, N.E.; Mackenzie, J.M.; White, P.A. The adenosine analogue NITD008 has potent antiviral activity against human and animal caliciviruses. Viruses 2019, 11, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, J.; Hurley, K.; Pesavento, P.A.; Poland, A.; Pedersen, N.C. Virulent systemic feline calicivirus infection: Local cytokine modulation and contribution of viral mutants. J. Feline Med. Surg. 2006, 8, 55–61. [Google Scholar] [CrossRef]
- Brunet, S. Multiple Correspondence Analysis on amino acids properties within the variable region of the capsid protein shows differences between classical and virulent systemic Feline Calicivirus strains. Viruses 2019, 11, 1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyne, K.P.; Gaskell, R.M.; Dawson, S.; Porter, C.J.; Radford, A.D. Evolutionary mechanisms of persistence and diversification of a calicivirus within endemically infected natural host populations. J. Virol. 2007, 81, 1961–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, B.; Lanave, G.; Di Profio, F.; Melegari, I.; Marsilio, F.; Camero, M.; Catella, C.; Capozza, P.; Bányai, K.; Barrs, V.R.; et al. Identification of feline calicivirus in cats with enteritis. Transbound Emerg. Dis. 2020, 67, 2579–2588. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, D.N.; Baker, K.D. Sudden death in kittens associated with a feline picornavirus. Aust. Vet. J. 1972, 48, 643. [Google Scholar] [CrossRef] [PubMed]
- Love, D.N.; Zuber, R.M. Feline calicivirus associated with pyrexia, profound anorexia and oral and perianal ulceration in a cat. Aust. Vet. Pract. 1987, 17, 136–137. [Google Scholar]
- Ellis, T.M. Jaundice in a Siamese cat with in utero feline calicivirus infection. Aust. Vet. J. 1981, 57, 383–385. [Google Scholar] [CrossRef]
- Cooper, L.M.; Sabine, M. Paw and mouth disease in a cat. Aust. Vet. J. 1972, 48, 644. [Google Scholar] [CrossRef]
- Hurley, K.F.; Pesavento, P.A.; Pedersen, N.C.; Poland, A.M.; Wilson, E.; Foley, J.E. An outbreak of virulent systemic feline calicivirus disease. J. Am. Vet. Med. Assoc. 2004, 224, 241–249. [Google Scholar] [CrossRef]
- Deschamps, J.Y.; Topie, E.; Roux, F. Nosocomial feline calicivirus-associated virulent systemic disease in a veterinary emergency and critical care unit in France. JFMS Open Rep. 2015, 1, 2055116915621581. [Google Scholar] [CrossRef] [Green Version]
- Coyne, K.; Jones, B.; Kipar, A.; Chantrey, J.; Porter, C.; Barber, P.; Dawson, S.; Gaskell, R.; Radford, A. Lethal outbreak of disease associated with feline calicivirus infection in cats. Vet. Rec. 2006, 158, 544–550. [Google Scholar] [CrossRef]
- Ossiboff, R. Feline caliciviruses (FCVs) isolated from cats with virulent systemic disease possess in vitro phenotypes distinct from those of other FCV isolates. J. Gen. Virol. 2007, 88, 506–517. [Google Scholar] [CrossRef]
- Coyne, K.P.; Dawson, S.; Radford, A.D.; Cripps, P.J.; Porter, C.J.; McCracken, C.M.; Gaskell, R.M. Long-Term analysis of feline calicivirus prevalence and viral shedding patterns in naturally infected colonies of domestic cats. Vet. Microbiol. 2006, 118, 12–25. [Google Scholar] [CrossRef]
- Willi, B.; Spiri, A.M.; Meli, M.L.; Samman, A.; Hoffmann, K.; Sydler, T.; Cattori, V.; Graf, F.; Diserens, K.A.; Padrutt, I.; et al. Molecular characterization and virus neutralization patterns of severe, non-epizootic forms of feline calicivirus infections resembling virulent systemic disease in cats in Switzerland and in Liechtenstein. Vet. Microbiol. 2016, 182, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Kershaw, O.; Klopfleisch, R. Feline calicivirus-associated virulent systemic disease: Not necessarily a local epizootic problem. Vet. Rec. 2011, 168, 589. [Google Scholar] [CrossRef]
- Brunet, S.; Jas, D.; David, F.; Bublot, M.; Poulet, H. Feline calicivirus: Vaccinations against virulent strains. In Proceedings of the Conference of the European Society of Veterinary Virology 2005: Comparative and Emerging Virus Infections of Dogs and Cats, Liverpool, UK, 20–22 June 2005. [Google Scholar]
- Rocha-Pereira, J.; Jochmans, D.; Dallmeier, K.; Leyssen, P.; Cunha, R.; Costa, I.; Nascimento, M.; Neyts, J. Inhibition of norovirus replication by the nucleoside analogue 2′-C-methylcytidine. Biochem. Biophys. Res. Commun. 2012, 427, 796–800. [Google Scholar] [CrossRef]
- Yin, Z.; Chen, Y.-L.; Schul, W.; Wang, Q.-Y.; Gu, F.; Duraiswamy, J.; Kondreddi, R.R.; Niyomrattanakit, P.; Lakshminarayana, S.B.; Goh, A. An adenosine nucleoside inhibitor of dengue virus. Proc. Natl. Acad. Sci. USA 2009, 106, 20435–20439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussar, D.A. New drugs of 2003. J. Am. Pharm. Assoc. 2004, 44, 168–210. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.F. Nitazoxanide in the treatment of acquired immune deficiency syndrome-related cryptosporidiosis: Results of the United States compassionate use program in 365 patients. Aliment. Pharmacol. Ther. 2006, 24, 887–894. [Google Scholar] [CrossRef]
- Stachulski, A.V.; Rossignol, J.F.; Pate, S.; Taujanskas, J.; Robertson, C.M.; Aerts, R.; Pascal, E.; Piacentini, S.; Frazia, S.; Santoro, M.G.; et al. Synthesis, antiviral activity, preliminary pharmacokinetics and structural parameters of thiazolide amine salts. Future Med. Chem. 2021. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Li, D.; Xie, Y.; Wang, K.; Zhang, Y.; Li, G.; Zhang, Q.; Chen, X.; Teng, Y.; Zhao, S. Nitazoxanide protects cats from feline calicivirus infection and acts synergistically with mizoribine in vitro. Antivir. Res. 2020, 182, 104827. [Google Scholar] [CrossRef] [PubMed]
- Gookin, J.L.; Levy, M.G.; Law, J.M.; Papich, M.G.; Poore, M.F.; Breitschwerdt, E.B. Experimental infection of cats with Tritrichomonas foetus. Am. J. Vet. Res. 2001, 62, 1690–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Date in 2015 | Breed, Sex | Age | Origin | Vacc. Status | Exposure History and Clinical Signs | Disease Phenotype | CPEs | Duration of Illness | Outcome |
|---|---|---|---|---|---|---|---|---|---|---|
| NSW_1 Index case | 15/11 | DSH F | 1.5 m | MCH | Unvacc. | Index case URT signs, lameness. | URTD | N/A | 2 d | R |
| NSW_2 Index case | 15/11 | DSH F | 1.5 m | MCH | Unvacc. | Index case URT signs, lameness. | URTD | N/A | 4 d | R |
| NSW_3 | 02/12 | DSH F | 4 m | SCH | Unknown | Pyrexia, lethargy, bilateral forelimb oedema, jaundice. Onset of CS 2 days after a surgical procedure (hindlimb amputation). | VSD | N/A | 4 d | D |
| NSW_4 | 06/12 | DSH M | 1.25 m | SCH | Unvacc. | Pyrexia, lethargy, facial and forelimb oedema. Onset of CS 7 days after a surgical procedure (abscess drainage). | VSD | N/A | 5 d | D |
| NSW_5 1,2,3 | 07/12 | DSH FN | 6 y | SCH | Incomp. | Pyrexia, multiple limb oedema, jaundice. Onset of CS 4 days after a surgical procedure (abscess drainage). | VSD | 24 h | 7 d | D |
| NSW_6 | 09/12 | DSH MN | 1 y | SCH | Complete | Pyrexia, forelimb oedema. Onset of CS 4 days after a surgical procedure (abscess drainage). | VSD | N/A | 9 d | R |
| NSW_7 | 14/12 | DSH M | 2 y | MCH | Complete | Pyrexia, facial and forelimb oedema, oral ulcers. | VSD | N/A | 2 d | E |
| NSW_8 | 14/12 | DSH FN | 3 y | SCH | Complete | Pyrexia, facial oedema. Onset of CS 5 days after a surgical procedure (jaw fracture repair). | VSD | N/A | 1 d | E |
| NSW_9 1,2 | 18/12 | DSH F | 1.5 m | MCH | Unknown | Pyrexia, facial oedema. Onset of CS 4 days after a surgical procedure (for intestinal intussusception). | VSD | 24 h | 1 d | E |
| Case | Date in 2017 | Breed, Sex | Age | Origin | Vacc. Status | Exposure History and Clinical Signs (CS) | Disease Phenotype | CPEs | Duration of Illness | Outcome |
|---|---|---|---|---|---|---|---|---|---|---|
| QLD_1 | 28/08 | DSH FN | 3 y | MCH | Complete | Acute respiratory effort, jaundice. Onset of CS 5 days after a surgical procedure (neutering). | VSD | N/A | 2 d | D |
| QLD_2 | 30/08 | DSH FN | 1 y | SCH | Complete | URT signs, facial oedema, oral ulcers. Onset of CS 5 days after a surgical procedure (neutering). | VSD | N/A | 9 d | R |
| QLD_3 | 06/09 | DSH MN | 4 y | SCH | Complete | Pyrexia, jaundice, facial/limb oedema. Onset of CS 7 days after a surgical procedure (abscess drainage). | VSD | N/A | 13 d | R |
| QLD_4 | 25/9 | DSH FN | 10 m | SCH | Complete | Pyrexia, facial/limb oedema, jaundice, dyspnoea. Onset of CS 4 days after a surgical procedure (neutering). | VSD | N/A | 6 d | E |
| QLD_5 *,1,2 | 05/10 | DSH MN | 1 y | MCH | Complete | Pyrexia, facial/limb oedema, jaundice, oral/skin ulcers. Onset of CS 9 days after a surgical procedure (limb amputation). | VSD | 48 h | 6 d | R |
| QLD_6 **,1,2 | 06/10 | DSH FN | 1 y | MCH | Complete | Lameness, limb oedema, oral ulcers, elevated bilirubin. Onset of CS 7 days after a surgical procedure (neutering). | VSD | 48 h | 6 d | R |
| QLD_7 **,1 | 09/10 | DSH FN | 1 y | MCH | Complete | Pyrexia, inappetence, limb oedema, elevated bilirubin. Onset of CS 10 days after a surgical procedure (neutering). | VSD | 72 h | 10 d | R |
| QLD_8 *,1,2 | 09/10 | DSH MN | 2 y | MCH | Complete | Pyrexia, inappetence, facia/limb oedema, oral ulcers, nasal discharge, sneezing. Household contact of QLD_5. | VSD | 48 h | 7 d | R |
| QLD_9 ***,1,2,3 | 09/10 | DSH FN | 1 y | MCH | Complete | Pyrexia, anorexia, limb oedema, nasal discharge, dyspnoea, jaundice. Onset of CS 11 days after a surgical procedure (neutering). | VSD | 36 h | 8 d | R |
| QLD_10 ***,1,2 | 09/10 | DSH FN | 1 y | MCH | Complete | Pyrexia, nasal discharge, facial/limb oedema. Onset of CS 11 days after a surgical procedure (neutering). | VSD | 36 h | 6 d | R |
| QLD_11 ***,1 | 09/10 | DSH FN | 3 y | MCH | Complete | Pyrexia, facial/limb oedema, jaundice, nasal discharge, hypothermia. Household contact of QLD_9 and QLD_10. | VSD | 72 h | 6 d | E |
| QLD_12 1,2,3 | 03/10 | DSH FN | 6 y | MCH | Complete | Pyrexia, anorexia, limb oedema, elevated bilirubin. Onset of CS 5 days after treatment for brown snake envenomation. | VSD | 36 h | 4 d | R |
| QLD_13 1,2 | 15/10 | Unkn. | N/A | MCH | Unkn. | Adult stray cat from a colony, oral ulcers. | URTD | N/A | Unkn. | Unkn. |
| Case | Date in 2018 | Breed, Sex | Age | Origin | Vacc. Status | Exposure History and Clinical Signs (CS) | Disease Phenotype | CPEs | Duration of Illness | Outcome |
|---|---|---|---|---|---|---|---|---|---|---|
| ACT_1 **,1,2 Index case | 22/01 | DSH FN | 4 m | MCH | Incomp. | Fever, inappetence, lameness, polyarthropathy. Adopted from a shelter. | URTD | 36 h | 14 d | R |
| ACT_2 **,1,2 Index case | 22/01 | DSH MN | 4 m | MCH | Incomp. | Fever, lethargy, sneezing. Adopted from a shelter. | URTD | 36 h | 14 d | R |
| ACT_3 ****,1,2 | 23/01 | DSH M | 10 y | MCH | Unvacc. | No CS. Adopted from a rescue society. Presented on 23/01 for a dental procedure. | Asympt. | 60 h | N/A | N/A |
| ACT_4 **** | 30/01 | DSH FN | 5 y | MCH | Complete | Fever, pain on abdominal palpation, limb oedema, myopathy, creatine kinase 20,206 U/L (RR < 261); AST 487 U/L, (RR < 60), myoglobinuria. Indoor cat co-housed with ACT_3. Onset of CS 7 days after ACT_3 had a dental procedure. | VSD | N/A | 9 d | R |
| ACT_5 | 02/02 | DSH MN | 8 y | MCH | Complete | Fever, inappetence, hypersalivation, jaundice, lumbar muscle pain, facial/limb oedema, ulcerated nasal planum. Onset of CS 4 days after a dental procedure. | VSD | N/A | 21 d | R |
| ACT_6 *** | 07/02 | DSH FN | 5 y | MCH | Complete | Vomiting, fever, painful kidneys on abdominal palpation, marked facial and limb oedema (all limbs), subcutaneous oedema of flanks, oral ulcers, swollen nose. Onset of CS 14 days after dental check-up. | VSD | N/A | 10 d | R |
| ACT_7 *,1,2 | 08/02 | Maine Coon MN | 10 m | MCH | Complete | Fever, inappetence, oral ulcers. Onset of CS 8 days after ACT_8 had a dental procedure. | URTD | 48 h | 7 d | R |
| ACT_8 * | 08/02 | DSH MN | 15 y | MCH | Unknown | Fever, inappetence, nasal planum ulcers. Onset of CS 8 days after being admitted to the hospital for a dental procedure. | URTD | 12 d | R | |
| ACT_9 ***,1,2 | 14/02 | Ragdoll cross MN | 4 y | MCH | Comp. | Fever, inappetence, lethargy, facial/limb oedema, swollen nose. Indoor cat co-housed with ACT-6. | VSD | 36 h | 7 d | R |
| Strain | Amino-Acid Residues and Physico-Chemical Properties Associated with VSD Pathotype | ||||||
|---|---|---|---|---|---|---|---|
| 438 Hydrophobic, Aliphatic ILV | 440 Non-Small EFHKIL MQRWY | 448 Polar, Positive Charge HKR | 452 Non-Small EFHKIL MQRWY | 455 Non-Negative All Except DE | 465 Polar DEHKN QRSTWI | 492 Small ADGNP STV | |
| FCV-VSD | V9T7 (9) | Q6G4E4SK (11) | K7A2E27 (7) | E11D6 (11) | T6D3M2I2NES (12) | S14G3 (14) | V16R (16) |
| FCV-URTD | T37V2I | G22S6Q4R2A2END | A30P4G3K3 | D36E3N | D28T5S3G2VR | G26S14 | V17L8I6R5K2 |
| E-ITA/2013/160 | V | Q | R | E | T | G | V |
| E-FCV ITA others | T3V11 | G2SR | A4 | D4 | D3E | G3S | L3V2 |
| E-ITA/201X/81 | V | S | A | D | E | S | V |
| E-ITA/201X/I48 | T | R | A | D | D | G | L |
| E-ITA/201X/I80 | T | G | A | D | D | G | L |
| E-ITA/201X/I82 | T | G | A | D | D | G | L |
| ACT7_URTD | T | G | K | E | D | S | V |
| ACT9_VSD | T | G | K | E | D | S | V |
| ACT1_URTD | T | G | K | E | D | S | V |
| ACT3_Asympt. | T | G | K | E | D | S | V |
| ACT2_URTD | T | G | R | E | N | S | I |
| QLD13_URTD | T | G | A | D | D | G | V |
| QLD8_VSD | T | G | A | D | D | G | V |
| QLD 5_VSD | T | G | A | D | D | G | V |
| QLD6_VSD | T | G | A | D | D | G | V |
| QLD9_VSD | T | G | A | D | D | G | V |
| QLD10_VSD | T | G | A | D | D | G | V |
| QLD12_VSD | T | G | A | D | D | G | V |
| NSW5_V1_VSD | V | Q | K | E | I | S | V |
| NSW9_V1_VSD | V | Q | K | E | I | S | V |
| NSW5_V2_VSD | T | S | K | E | D | S | V |
| NSW9_V2_VSD | T | S | K | E | D | S | V |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bordicchia, M.; Fumian, T.M.; Van Brussel, K.; Russo, A.G.; Carrai, M.; Le, S.-J.; Pesavento, P.A.; Holmes, E.C.; Martella, V.; White, P.; et al. Feline Calicivirus Virulent Systemic Disease: Clinical Epidemiology, Analysis of Viral Isolates and In Vitro Efficacy of Novel Antivirals in Australian Outbreaks. Viruses 2021, 13, 2040. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102040
Bordicchia M, Fumian TM, Van Brussel K, Russo AG, Carrai M, Le S-J, Pesavento PA, Holmes EC, Martella V, White P, et al. Feline Calicivirus Virulent Systemic Disease: Clinical Epidemiology, Analysis of Viral Isolates and In Vitro Efficacy of Novel Antivirals in Australian Outbreaks. Viruses. 2021; 13(10):2040. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102040
Chicago/Turabian StyleBordicchia, Matteo, Tulio Machado Fumian, Kate Van Brussel, Alice G. Russo, Maura Carrai, Shi-Jia Le, Patricia A. Pesavento, Edward C. Holmes, Vito Martella, Peter White, and et al. 2021. "Feline Calicivirus Virulent Systemic Disease: Clinical Epidemiology, Analysis of Viral Isolates and In Vitro Efficacy of Novel Antivirals in Australian Outbreaks" Viruses 13, no. 10: 2040. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102040