
The Expression Level of HIV-1 Vif Is Optimized by Nucleotide Changes in the Genomic SA1D2prox Region during the Viral Adaptation Process

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
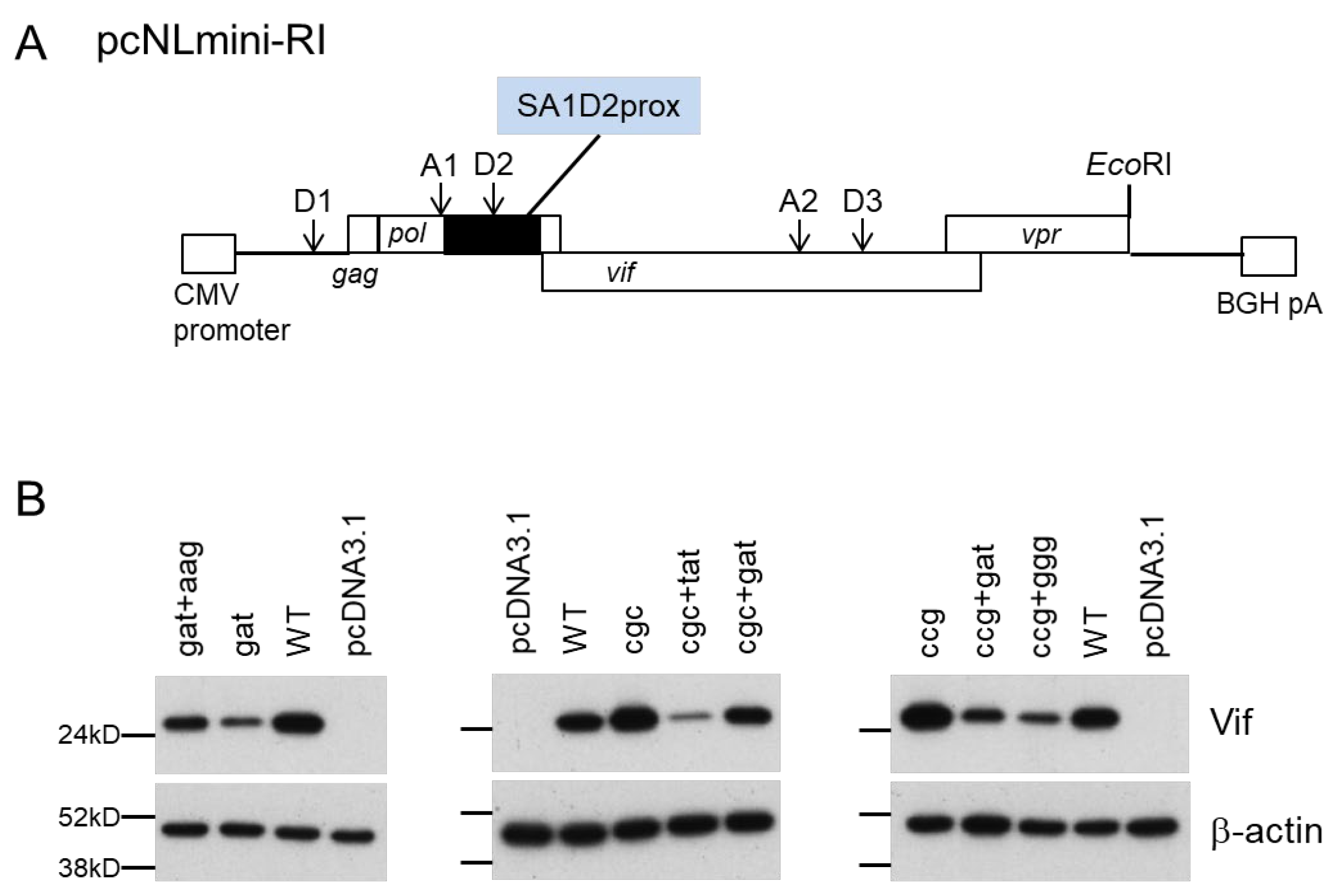
2.1. Plasmids
2.2. Cells
2.3. Adaptation Experiments
2.4. Infection Experiments
2.5. Western Blotting Analysis
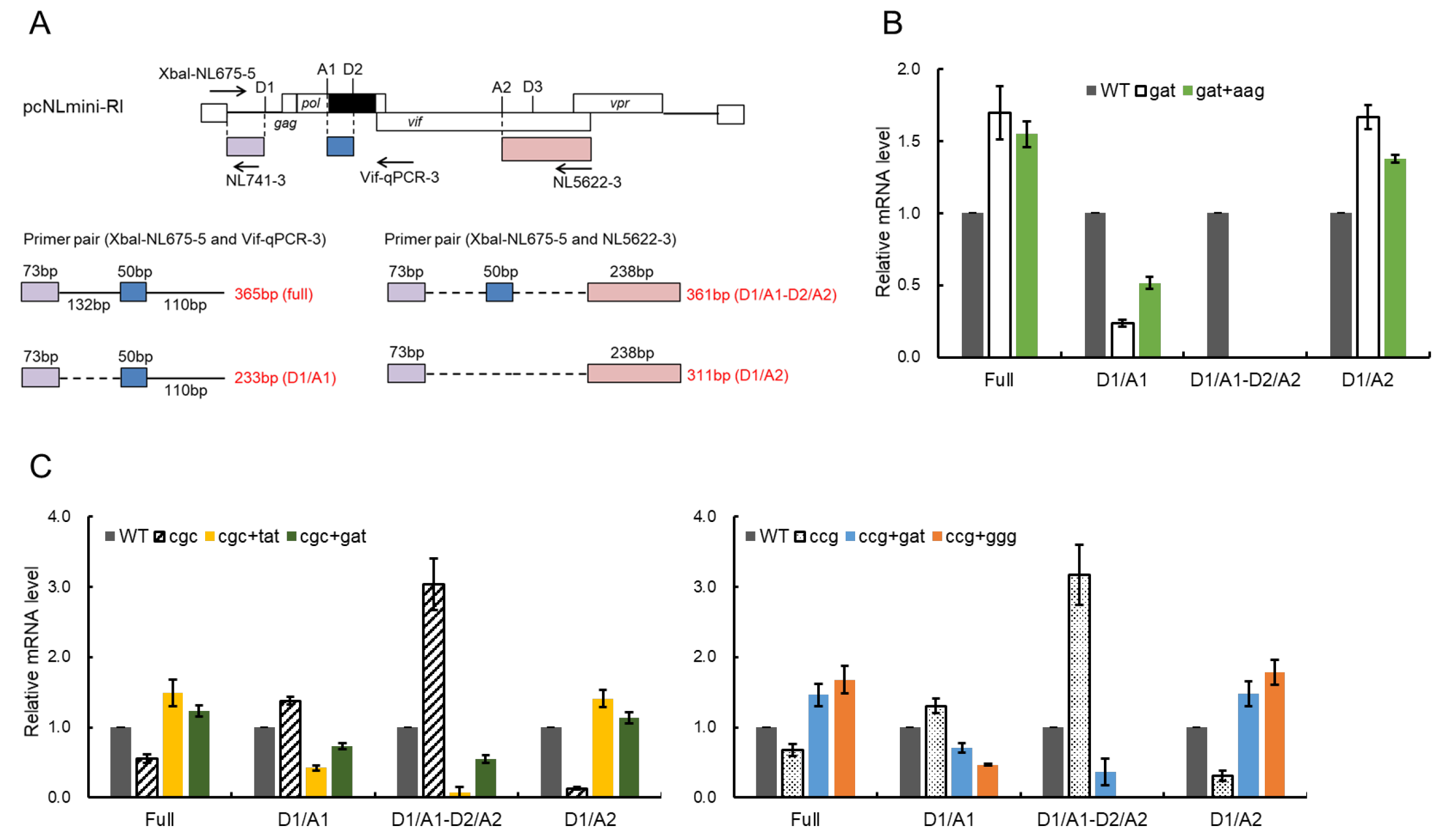
2.6. Semiquantitative RT-PCR Analysis of Splicing Products
3. Results
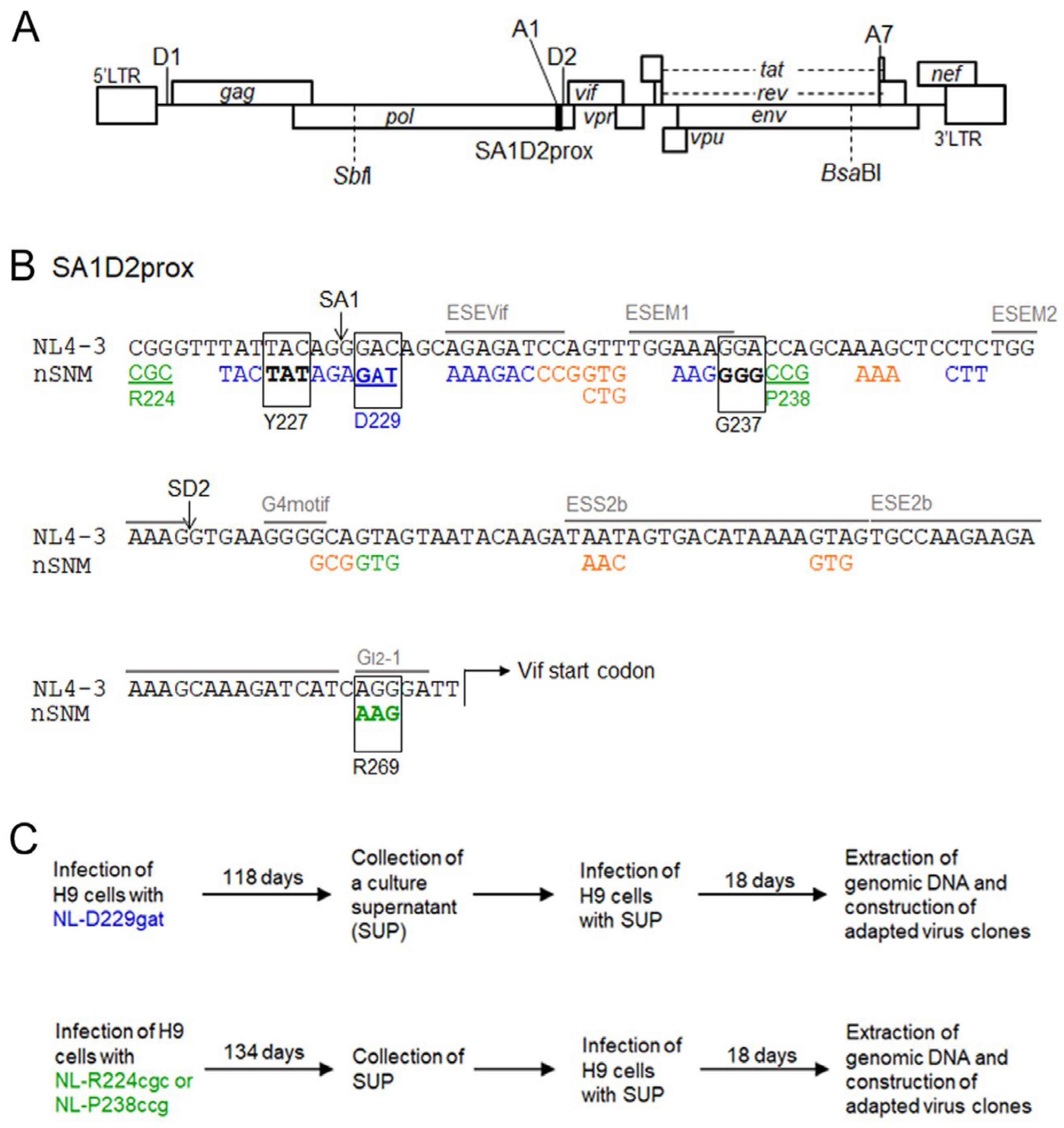
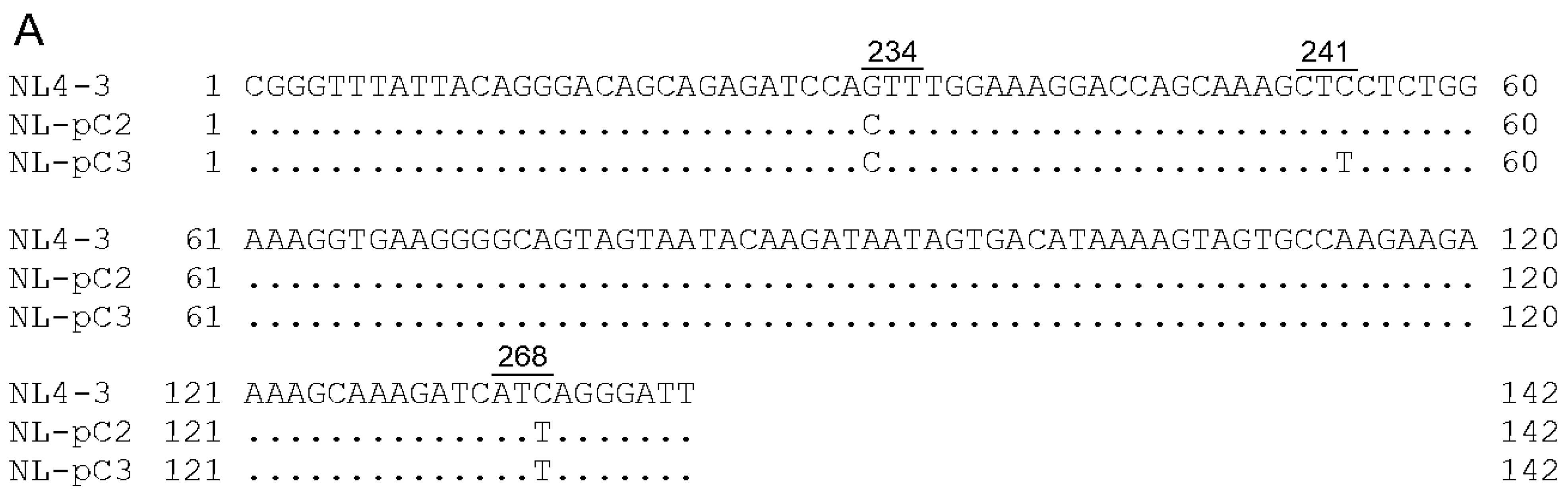
3.1. Viral Clones Used for the Adaptation Experiments
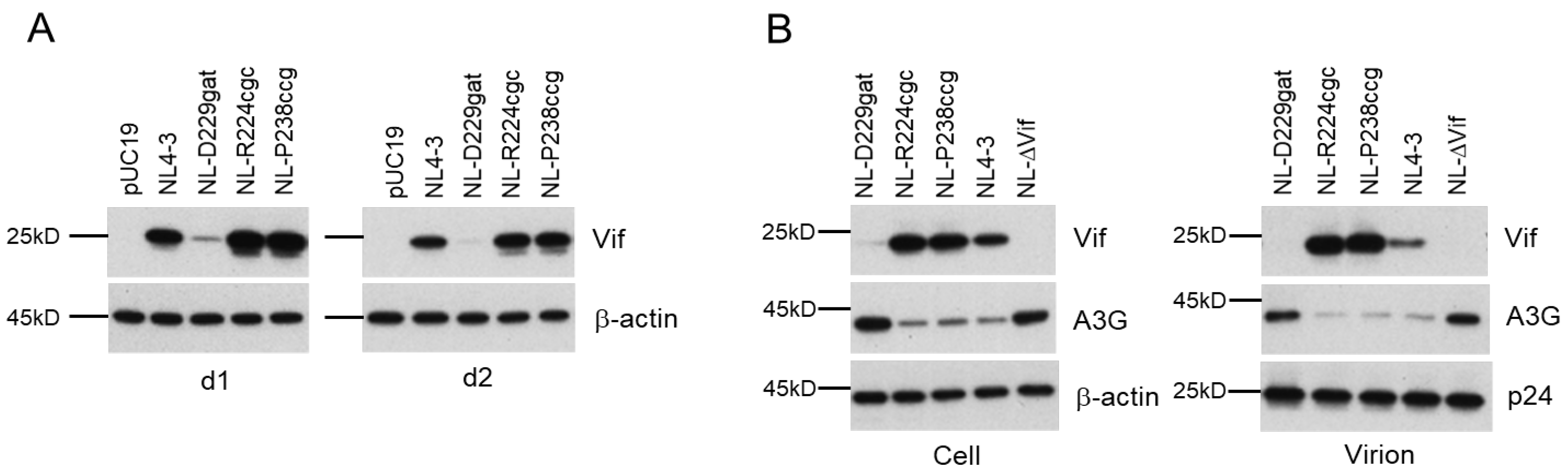
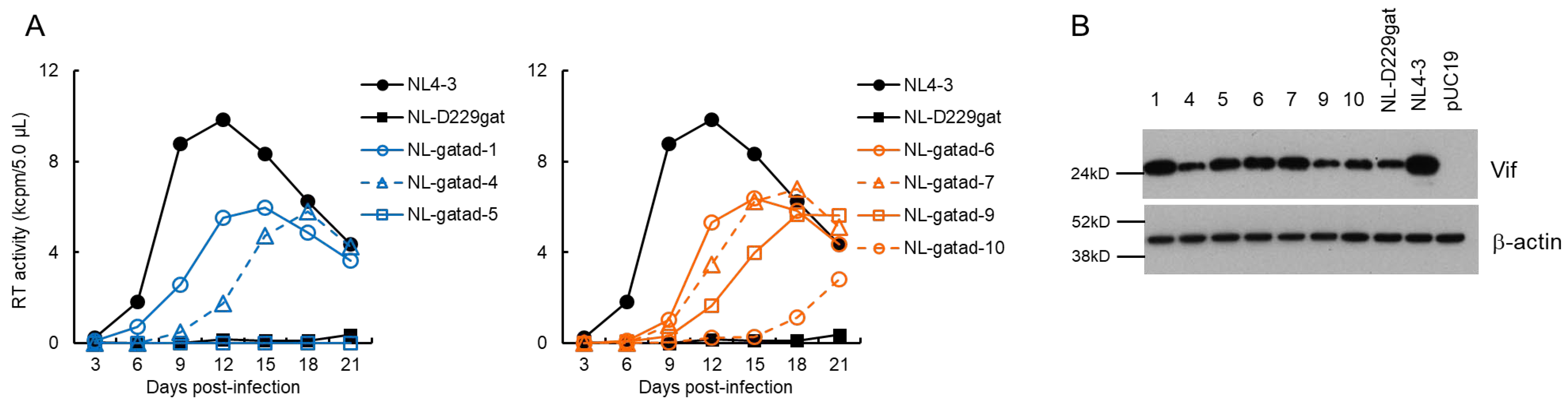
3.2. Characteristics of Viral Clones Obtained from Low-Vif Type NL-D229gat Adaptation
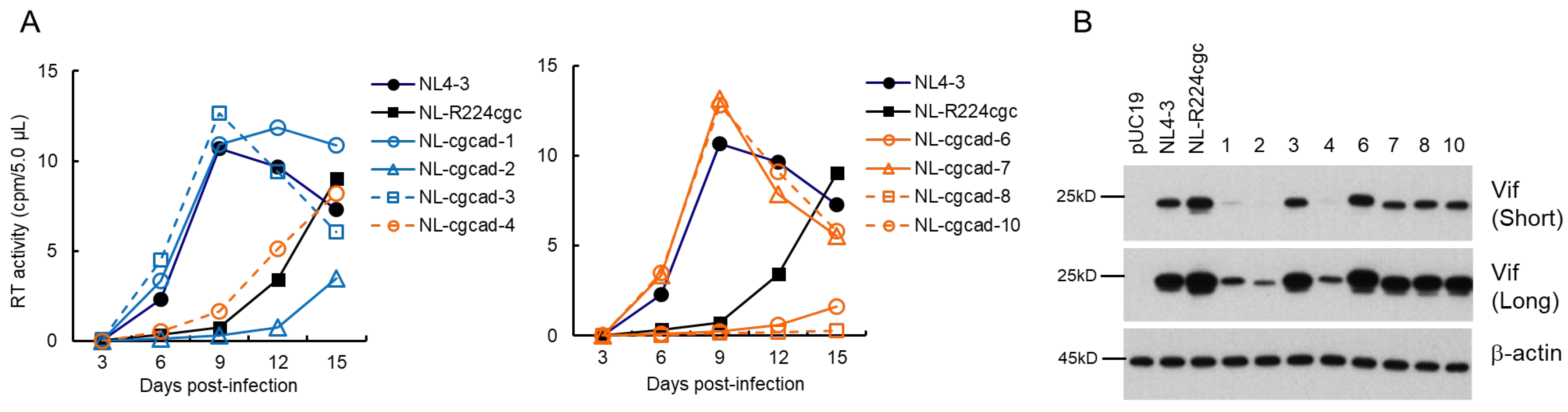
3.3. Characteristics of Viral Clones Obtained from Excessive-Vif Type NL-R224cgc Adaptation
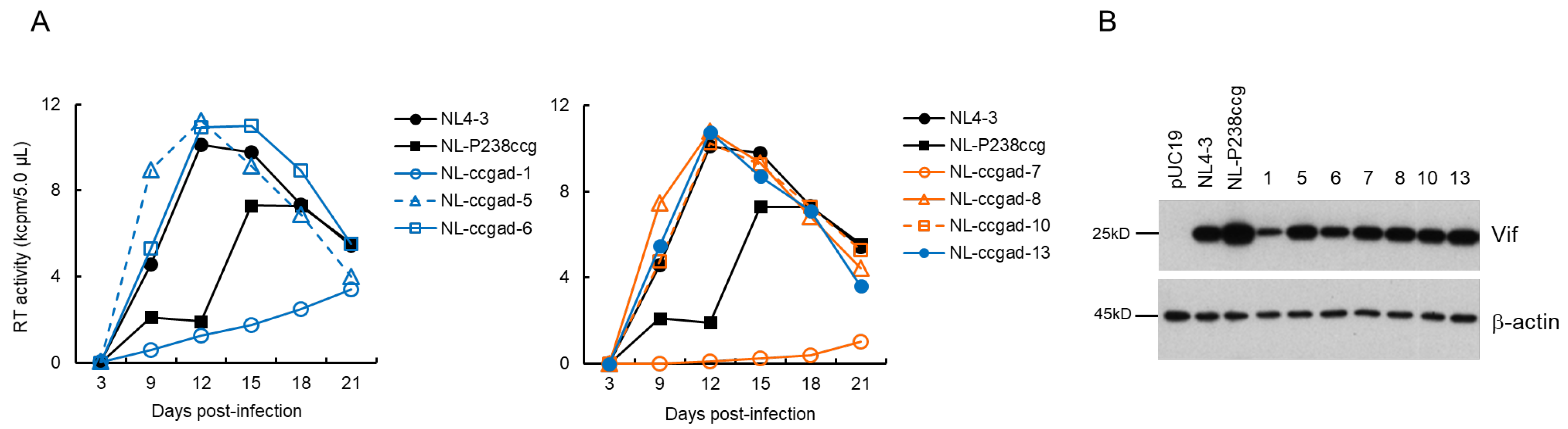
3.4. Characteristics of Viral Clones Obtained from Excessive-Vif Type NL-P238ccg Adaptation
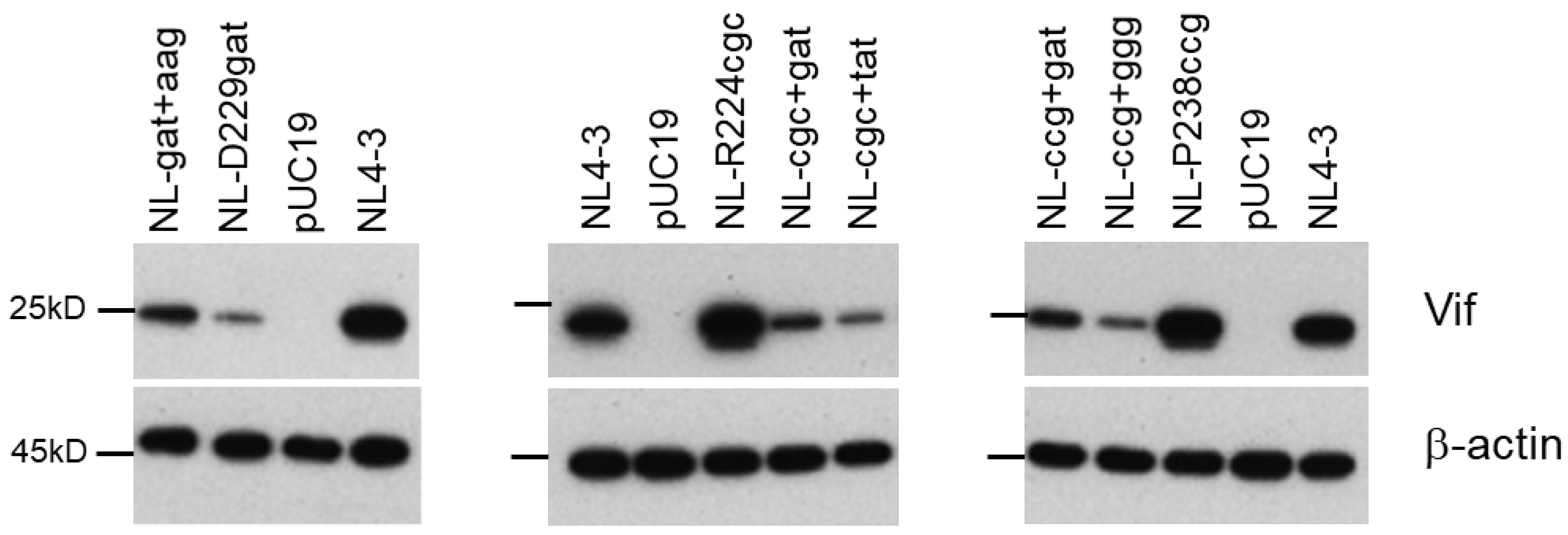
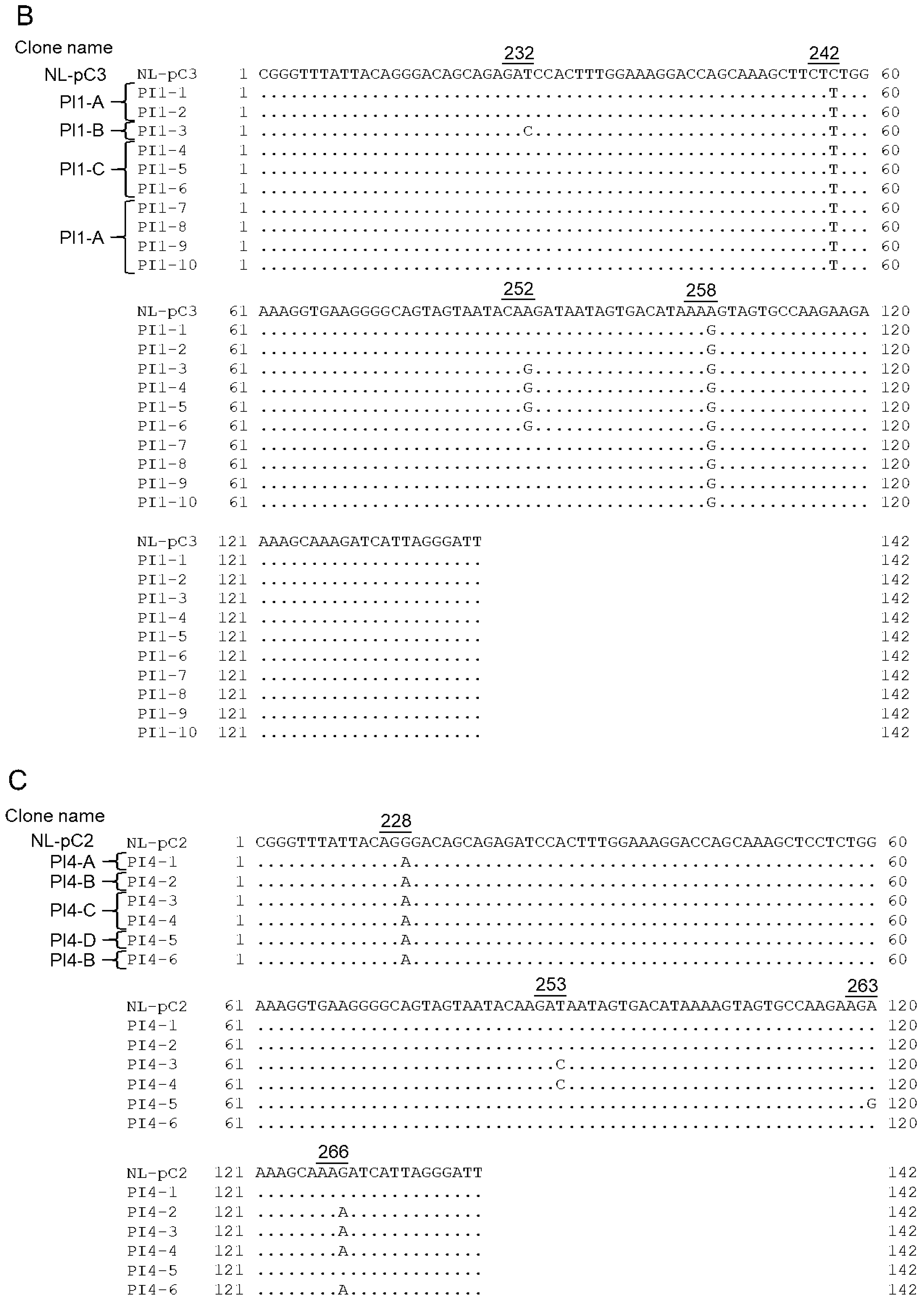
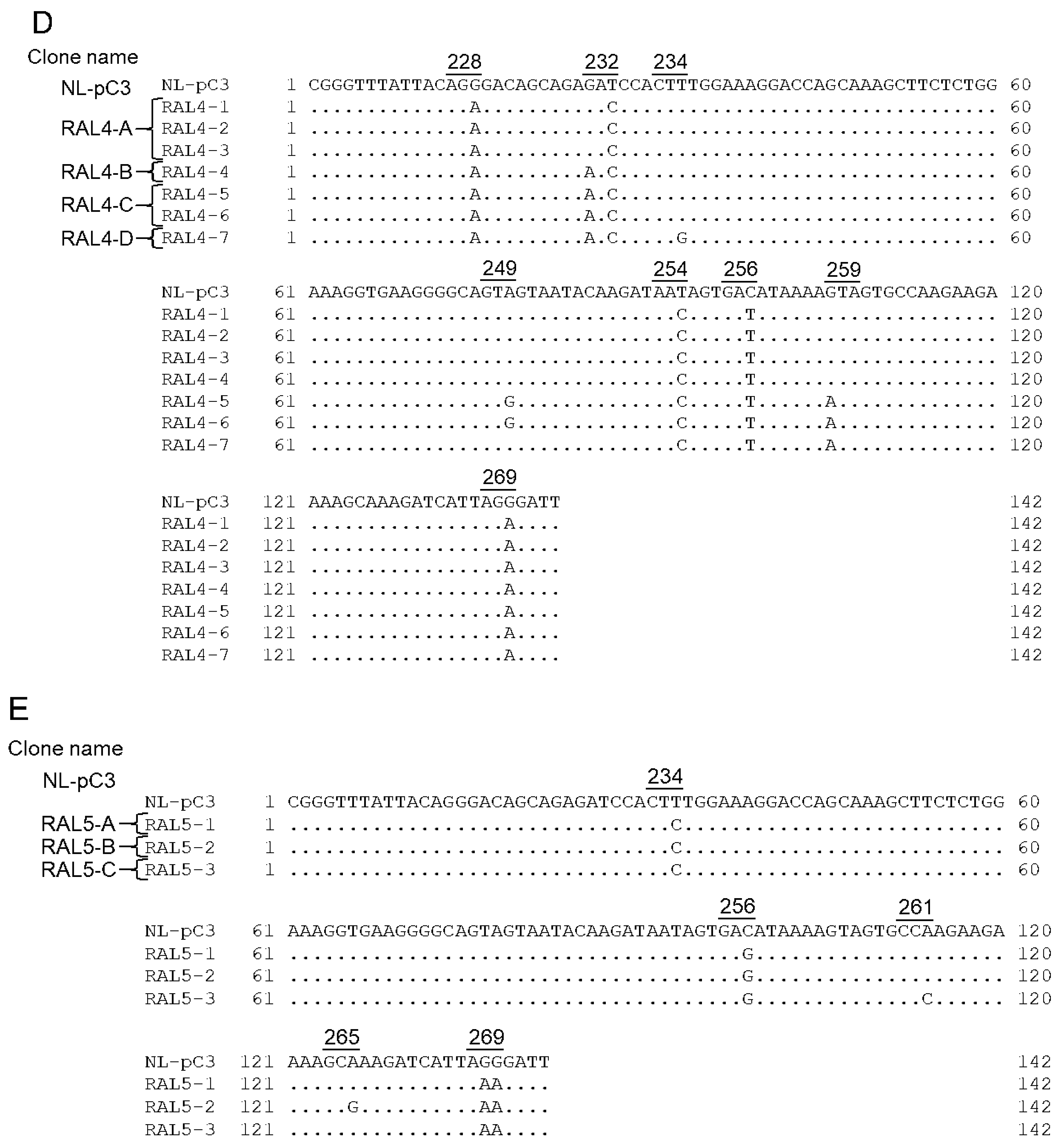
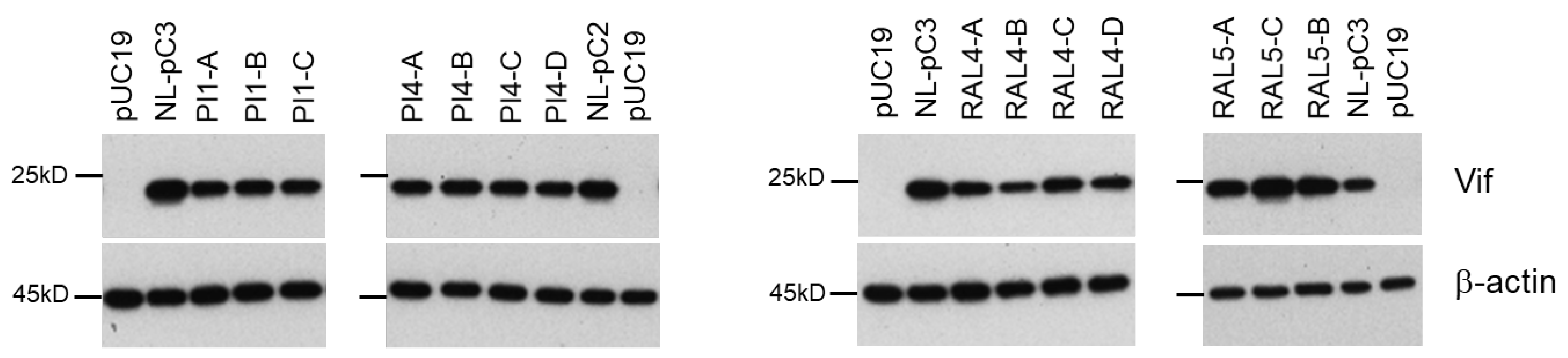
3.5. Identification of Adaptive Mutations Associated with the Variation in Vif Expression Level
3.6. Effect of Adaptive Mutations within the SA1D2prox Region on the Splicing Pattern
3.7. Chronological Changes in the Vif Expression Levels by Mutations within SA1D2prox in Infected Individuals
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Malim, M.H.; Emerman, M. HIV-1 accessory proteins—Ensuring viral survival in a hostile environment. Cell Host Microbe 2008, 3, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.S.; Hultquist, J.F.; Evans, D.T. The restriction factors of human immunodeficiency virus. J. Biol. Chem. 2012, 287, 40875–40883. [Google Scholar] [CrossRef] [Green Version]
- Malim, M.H.; Bieniasz, P.D. HIV restriction factors and mechanisms of evasion. Cold Spring Harb. Perspect. Med. 2012, 2, a006940. [Google Scholar] [CrossRef] [PubMed]
- Aydin, H.; Taylor, M.W.; Lee, J.E. Structure-guided analysis of the human APOBEC3-HIV restrictome. Structure 2014, 22, 668–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desimmie, B.A.; Delviks-Frankenberrry, K.A.; Burdick, R.C.; Qi, D.; Izumi, T.; Pathak, V.K. Multiple APOBEC3 restriction factors for HIV-1 and one Vif to rule them all. J. Mol. Biol. 2014, 426, 1220–1245. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Baig, T.T.; Love, R.P.; Chelico, L. Suppression of APOBEC3-mediated restriction of HIV-1 by Vif. Front. Microbiol. 2014, 5, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, A.; Iwatani, Y. APOBEC3G-mediated G-to-A hypermutation of the HIV-1 genome: The missing link in antiviral molecular mechanisms. Front. Microbiol. 2016, 7, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupfler, B.; Verriez, C.; Gallois-Montbrun, S.; Marquet, R.; Paillart, J.C. Degradation-independent inhibition of APOBEC3G by the HIV-1 Vif protein. Viruses 2021, 13, 617. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Chertova, E.; Chen, J.; Ott, D.E.; Roser, J.D.; Hu, W.S.; Pathak, V.K. Stoichiometry of the antiviral protein APOBEC3G in HIV-1 virions. Virology 2007, 360, 247–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Browne, E.P.; Allers, C.; Landau, N.R. Restriction of HIV-1 by APOBEC3G is cytidine deaminase-dependent. Virology 2009, 387, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Felber, B.K.; Benko, D.M.; Fenyö, E.M.; Pavlakis, G.N. Cloning and functional analysis of multiply spliced mRNA species of human immunodeficiency virus type 1. J. Virol. 1990, 64, 2519–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, D.F.J.; Martin, M.A. Alternative splicing of human immunodeficiency virus type 1 mRNA modulates viral protein expression, replication, and infectivity. J. Virol. 1993, 67, 6365–6378. [Google Scholar] [CrossRef] [Green Version]
- Amendt, B.A.; Si, Z.H.; Stoltzfus, C.M. Presence of exon splicing silencers within human immunodeficiency virus type 1 tat exon 2 and tat-rev exon 3: Evidence for inhibition mediated by cellular factors. Mol. Cell. Biol. 1995, 15, 4606–4615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emery, A.; Zhou, S.; Pollom, E.; Swanstrom, R. Characterizing HIV-1 splicing by using next-generation sequencing. J. Virol. 2017, 91, e02515-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bevilacqua, P.C.; Ritchey, L.E.; Su, Z.; Assmann, S.M. Genome-wide analysis of RNA secondary structure. Annu. Rev. Genet. 2016, 50, 235–266. [Google Scholar] [CrossRef] [Green Version]
- Sertznig, H.; Hillebrand, F.; Erkelenz, S.; Schaal, H.; Widera, M. Behind the scenes of HIV-1 replication: Alternative splicing as the dependency factor on the quiet. Virology 2018, 516, 176–188. [Google Scholar] [CrossRef]
- Emery, A.; Swanstrom, R. HIV-1: To splice or not to splice, that is the question. Viruses 2021, 13, 181. [Google Scholar] [CrossRef]
- Kammler, S.; Otte, M.; Hauber, I.; Kjems, J.; Hauber, J.; Schaal, H. The strength of the HIV-1 30 splice sites affects Rev function. Retrovirology 2006, 3, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exline, C.M.; Feng, Z.; Stoltzfus, C.M. Negative and positive mRNA splicing elements act competitively to regulate human immunodeficiency virus type 1 vif gene expression. J. Virol. 2008, 82, 3921–3931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brillen, A.L.; Walotka, L.; Hillebrand, F.; Müller, L.; Widera, M.; Theiss, S.; Schaal, H. Analysis of competing HIV-1 splice donor sites uncovers a tight cluster of splicing regulatory elements within exon 2/2b. J. Virol. 2017, 91, e00389-17. [Google Scholar] [CrossRef] [Green Version]
- Widera, M.; Erkelenz, S.; Hillebrand, F.; Krikoni, A.; Widera, D.; Kaisers, W.; Deenen, R.; Gombert, M.; Dellen, R.; Pfeiffer, T.; et al. An intronic G run within HIV-1 intron 2 is critical for splicing regulation of vif mRNA. J. Virol. 2013, 87, 2707–2720. [Google Scholar] [CrossRef] [Green Version]
- Mansky, L.M.; Temin, H.M. Lower in vivo mutation rate of human immunodeficiency virus type 1 than that predicted from the fidelity of purified reverse transcriptase. J. Virol. 1995, 69, 5087–5094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D.N.; Aldrovandi, G.M.; Kutsch, O.; Shaw, G.M. Dynamics of HIV-1 recombination in its natural target cells. Proc. Natl. Acad. Sci. USA 2004, 101, 4204–4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffin, J.M. HIV population dynamics in vivo: Implications for genetic variation, pathogenesis, and therapy. Science 1995, 267, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malim, M.H.; Emerman, M. HIV-1 sequence variation: Drift, shift, and attenuation. Cell 2001, 104, 469–472. [Google Scholar] [CrossRef] [Green Version]
- van Opijnen, T.; Berkhout, B. The host environment drives HIV-1 fitness. Rev. Med. Virol. 2005, 15, 219–233. [Google Scholar] [CrossRef]
- Carlson, J.M.; Le, A.Q.; Shahid, A.; Brumme, Z.L. HIV-1 adaptation to HLA: A window into virus-host immune interactions. Trends Microbiol. 2015, 23, 212–224. [Google Scholar] [CrossRef] [PubMed]
- Nijhuis, M.; van Maarseveen, N.M.; Lastere, S.; Schipper, P.; Coakley, E.; Glass, B.; Rovenska, M.; de Jong, D.; Chappey, C.; Goedegebuure, I.W.; et al. A novel substrate-based HIV-1 protease inhibitor drug resistance mechanism. PLoS Med. 2007, 4, e36. [Google Scholar] [CrossRef]
- Parry, C.M.; Kolli, M.; Myers, R.E.; Cane, P.A.; Schiffer, C.; Pillay, D. Three residues in HIV-1 matrix contribute to protease inhibitor susceptibility and replication capacity. Antimicrob. Agents Chemother. 2011, 55, 1106–1113. [Google Scholar] [CrossRef] [Green Version]
- Rabi, S.A.; Laird, G.M.; Durand, C.M.; Laskey, S.; Shan, L.; Bailey, J.R.; Chioma, S.; Moore, R.D.; Siliciano, R.F. Multi-step inhibition explains HIV-1 protease inhibitor pharmacodynamics and resistance. J. Clin. Investig. 2013, 123, 3848–3860. [Google Scholar] [CrossRef]
- Blanch-Lombarte, O.; Santos, J.R.; Peña, R.; Jiménez-Moyano, E.; Clotet, B.; Paredes, R.; Prado, J.G. HIV-1 Gag mutations alone are sufficient to reduce darunavir susceptibility during virological failure to boosted PI therapy. J. Antimicrob. Chemother. 2020, 75, 2535–2546. [Google Scholar] [CrossRef]
- Hikichi, Y.; Van Duyne, R.; Pham, P.; Groebner, J.L.; Wiegand, A.; Mellors, J.W.; Kearney, M.F.; Freed, E.O. Mechanistic analysis of the broad antiretroviral resistance conferred by HIV-1 envelope glycoprotein mutations. mBio 2021, 12, e03134-20. [Google Scholar] [CrossRef]
- Ikeda, T.; Symeonides, M.; Albin, J.S.; Li, M.; Thali, M.; Harris, R.S. HIV-1 adaptation studies reveal a novel Env-mediated homeostasis mechanism for evading lethal hypermutation by APOBEC3G. PLoS Pathog. 2018, 14, e1007010. [Google Scholar] [CrossRef] [Green Version]
- Haché, G.; Abbink, T.E.; Berkhout, B.; Harris, R.S. Optimal translation initiation enables Vif-deficient human immunodeficiency virus type 1 to escape restriction by APOBEC3G. J. Virol. 2009, 83, 5956–5960. [Google Scholar] [CrossRef] [Green Version]
- Miyagi, E.; Kao, S.; Fumitaka, M.; Buckler-White, A.; Plishka, R.; Strebel, K. Long-term passage of Vif-null HIV-1 in CD4+ T cells expressing sub-lethal levels of APOBEC proteins fails to develop APOBEC resistance. Virology 2017, 504, 1–11. [Google Scholar] [CrossRef]
- Nomaguchi, M.; Doi, N.; Fujiwara, S.; Saito, A.; Akari, H.; Nakayama, E.E.; Shioda, T.; Yokoyama, M.; Sato, H.; Adachi, A. Systemic biological analysis of the mutations in two distinct HIV-1mt genomes occurred during replication in macaque cells. Microbes Infect. 2013, 15, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Nomaguchi, M.; Miyake, A.; Doi, N.; Fujiwara, S.; Miyazaki, Y.; Tsunetsugu-Yokota, Y.; Yokoyama, M.; Sato, H.; Masuda, T.; Adachi, A. Natural single-nucleotide polymorphisms in the 3’ region of the HIV-1 pol gene modulate viral replication ability. J. Virol. 2014, 88, 4145–4160. [Google Scholar] [CrossRef] [Green Version]
- Nomaguchi, M.; Doi, N.; Sakai, Y.; Ode, H.; Iwatani, Y.; Ueno, T.; Matsumoto, Y.; Miyazaki, Y.; Masuda, T.; Adachi, A. Natural single-nucleotide variations in the HIV-1 genomic SA1prox region can alter viral replication ability by regulating Vif expression levels. J. Virol. 2016, 90, 4563–4578. [Google Scholar] [CrossRef] [Green Version]
- Nomaguchi, M.; Doi, N.; Yoshida, T.; Koma, T.; Adachi, S.; Ode, H.; Iwatani, Y.; Yokoyama, M.; Sato, H.; Adachi, A. Production of HIV-1 vif mRNA is modulated by natural nucleotide variations and SLSA1 RNA structure in SA1D2prox genomic region. Front. Microbiol. 2017, 8, 2542. [Google Scholar] [CrossRef] [PubMed]
- Doi, N.; Koma, T.; Adachi, A.; Nomaguchi, M. Expression level of HIV-1 Vif can be fluctuated by natural nucleotide variations in the vif-coding and regulatory SA1D2prox sequences of the proviral genome. Front. Microbiol. 2019, 10, 2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, A.; Gendelman, H.E.; Koenig, S.; Folks, T.; Willey, R.; Rabson, A.; Martin, M.A. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol. 1986, 59, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Ono, N.; Sakai, H.; Ogawa, K.; Shibata, R.; Kiyomasu, T.; Masuike, H.; Ueda, S. Generation and characterization of the human immunodeficiency virus type 1 mutants. Arch. Virol. 1991, 117, 45–58. [Google Scholar] [CrossRef]
- Yamashita, T.; Kamada, K.; Hatcho, K.; Adachi, A.; Nomaguchi, M. Identification of amino acid residues in HIV-1 Vif critical for binding and exclusion of APOBEC3G/F. Microbes Infect. 2008, 10, 1142–1149. [Google Scholar] [CrossRef]
- Yamashita, T.; Nomaguchi, M.; Miyake, A.; Uchiyama, T.; Adachi, A. Status of APOBEC3G/F in cells and progeny virions modulated by Vif determines HIV-1 infectivity. Microbes Infect. 2010, 12, 166–171. [Google Scholar] [CrossRef]
- Lebkowski, J.S.; Clancy, S.; Calos, M.P. Simian virus 40 replication in adenovirus-transformed human cells antagonizes gene expression. Nature 1985, 317, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Kamada, K.; Igarashi, T.; Martin, M.A.; Khamsri, B.; Hatcho, K.; Yamashita, T.; Fujita, M.; Uchiyama, T.; Adachi, A. Generation of HIV-1 derivatives that productively infect macaque monkey lymphoid cells. Proc. Natl. Acad. Sci. USA 2006, 103, 16959–16964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomaguchi, M.; Yokoyama, M.; Kono, K.; Nakayama, E.E.; Shioda, T.; Doi, N.; Fujiwara, S.; Saito, A.; Akari, H.; Miyakawa, K.; et al. Generation of rhesus macaque-tropic HIV-1 clones that are resistant to major anti-HIV-1 restriction factors. J. Virol. 2013, 87, 11447–11461. [Google Scholar] [CrossRef] [Green Version]
- Willey, R.L.; Smith, D.H.; Lasky, L.A.; Theodore, T.S.; Earl, P.L.; Moss, B.; Capon, D.J.; Martin, M.A. In vitro mutagenesis identifies a region within the envelope gene of the human immunodeficiency virus that is critical for infectivity. J. Virol. 1988, 62, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akari, H.; Fujita, M.; Kao, S.; Khan, M.A.; Shehu-Xhilaga, M.; Adachi, A.; Strebel, K. High level expression of human immunodeficiency virus type-1 Vif inhibits viral infectivity by modulating proteolytic processing of the Gag precursor at the p2/nucleocapsid processing site. J. Biol. Chem. 2004, 279, 12355–12362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, J.M.; Stoltzfus, C.M. A suboptimal 5′ splice site downstream of HIV-1 splice site A1 is required for unspliced viral mRNA accumulation and efficient virus replication. Retrovirology 2006, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Dang, Y.; Wang, X.; Esselman, W.J.; Zheng, Y.H. Identification of APOBEC3DE as another antiretroviral factor from the human APOBEC family. J. Virol. 2006, 80, 10522–10533. [Google Scholar] [CrossRef] [Green Version]
- Piroozmand, A.; Yamamoto, Y.; Khamsri, B.; Fujita, M.; Uchiyama, T.; Adachi, A. Generation and characterization of APOBEC3G-positive 293T cells for HIV-1 Vif study. J. Med. Investig. 2007, 54, 154–158. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Saenz, D.T.; Fadel, H.J.; Walker, W.; Peretz, M.; Poeschla, E.M. The HIV-1 central polypurine tract functions as a second line of defense against APOBEC3G/F. J. Virol. 2010, 84, 11981–11993. [Google Scholar] [CrossRef] [Green Version]
- Widera, M.; Hillebrand, F.; Erkelenz, S.; Vasudevan, A.A.; Münk, C.; Schaal, H. A functional conserved intronic G run in HIV-1 intron 3 is critical to counteract APOBEC3G-mediated host restriction. Retrovirology 2014, 11, 72. [Google Scholar] [CrossRef]
- Ode, H.; Matsuda, M.; Matsuoka, K.; Hachiya, A.; Hattori, J.; Kito, Y.; Yokomaku, Y.; Iwatani, Y.; Sugiura, W. Quasispecies analyses of the HIV-1 near-full-length genome with Illumina MiSeq. Front. Microbiol. 2015, 6, 1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takata, M.A.; Soll, S.J.; Emery, A.; Blanco-Melo, D.; Swanstrom, R.; Bieniasz, P.D. Global synonymous mutagenesis identifies cis-acting RNA elements that regulate HIV-1 splicing and replication. PLoS Pathog. 2018, 14, e1006824. [Google Scholar] [CrossRef] [Green Version]
- Pagani, F.; Raponi, M.; Baralle, F.E. Synonymous mutations in CFTR exon 12 affect splicing and are not neutral in evolution. Proc. Natl. Acad. Sci. USA 2005, 102, 6368–6372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presnyak, V.; Alhusaini, N.; Chen, Y.H.; Martin, S.; Morris, N.; Kline, N.; Olson, S.; Weinberg, D.; Baker, K.E.; Graveley, B.R.; et al. Codon optimality is a major determinant of mRNA stability. Cell 2015, 160, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, J.B.; Kudla, G. Synonymous but not the same: The causes and consequences of codon bias. Nat. Rev. Genet. 2011, 12, 32–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan-Paiz, A.; Franco, S.; Martínez, M.A. Impact of synonymous genome recoding on the HIV life cycle. Front. Microbiol. 2021, 12, 606087. [Google Scholar] [CrossRef]
- Moris, A.; Murray, S.; Cardinaud, S. AID and APOBECs span the gap between innate and adaptive immunity. Front. Microbiol. 2014, 5, 534. [Google Scholar] [CrossRef] [Green Version]
- Pollom, E.; Dang, K.K.; Potter, E.L.; Gorelick, R.J.; Burch, C.L.; Weeks, K.M.; Swanstrom, R. Comparison of SIV and HIV-1 genomic RNA structures reveals impact of sequence evolution on conserved and non-conserved structural motifs. PLoS Pathog. 2013, 9, e1003294. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NL-gatad-4 | NL-gatad-9 | ||||
|---|---|---|---|---|---|
| Nt Change | Region | NS/S Change in the Region | Nt Change | Region | NS/N Change in the Region |
| c4916t | Pol-IN (SA1D2prox) | D229gat | c4916t | Pol-IN (SA1D2prox) | D229gat |
| t5039c | Pol-IN (SA1D2prox) | D270gac | |||
| g5389a | Vif | E117K | g5389a | Vif | E117K |
| g6280a | Vpu | A74T | g6280a | Vpu | A74T |
| Env | M20I | Env | M20I | ||
| g7016a | Env | E266K | |||
| g7032a | Env | R271K | |||
| NL-Gatad-1 | NL-Gatad-6 | NL-Gatad-7 | ||||||
|---|---|---|---|---|---|---|---|---|
| Nt Change | Region | NS/S Change in the Region | Nt Change | Region | NS/N Change in the Region | Nt Change | Region | NS/N Change in the Region |
| g3301a | Pol-RT | S251N | g3301a | Pol-RT | S251N | g3301a | Pol-RT | S251N |
| g3341a | Pol-RT | L264tta | g3341a | Pol-RT | L264tta | g3341a | Pol-RT | L264tta |
| g3545a | Pol-RT | Q332caa | g3545a | Pol-RT | Q332caa | g3545a | Pol-RT | Q332caa |
| g4220a | Pol-RT | R557aga | ||||||
| g4374a | Pol-IN | A49T | g4374a | Pol-IN | A49T | g4374a | Pol-IN | A49T |
| c4916t | Pol-IN (SA1D2prox) | D229gat | c4916t | Pol-IN (SA1D2prox) | D229gat | c4916t | Pol-IN (SA1D2prox) | D229gat |
| g5035a | Pol-IN (SA1D2prox) | R269Kaag | g5035a | Pol-IN (SA1D2prox) | R269Kaag | g5035a | Pol-IN (SA1D2prox) | R269Kaag |
| a5702g | Vpr | E48gag | a5702g | Vpr | E48gag | a5702g | Vpr | E48gag |
| g6218a | Vpu | G53D | ||||||
| g6280a | Vpu | A74T | g6280a | Vpu | A74T | g6280a | Vpu | A74T |
| Env | M20I | Env | M20I | Env | M20I | |||
| g6603a | Env | S128N | g6603a | Env | S128N | |||
| 7388-7402del | Env | 390-394del (NSTWFdel) | 7388-7402del | Env | 390-394del (NSTWFdel) | |||
| NL-cgcad-1 | NL-cgcad-2 | NL-cgcad-4 | ||||||
|---|---|---|---|---|---|---|---|---|
| Nt Change | Region | NS/S Change in the Region | Nt Change | Region | NS/N Change in the Region | Nt Change | Region | NS/N Change in the Region |
| a3153t | Pol-RT | I202L | ||||||
| g4901c | Pol-IN (SA1D2prox) | R224cgc | g4901c | Pol-IN (SA1D2prox) | R224cgc | g4901c | Pol-IN (SA1D2prox) | R224cgc |
| c4910t | Pol-IN (SA1D2prox) | Y227tat | c4910t | Pol-IN (SA1D2prox) | Y227tat | |||
| c4916t | Pol-IN (SA1D2prox) | D229gat | ||||||
| c5030t | Pol-IN (SA1D2prox) | I267att | ||||||
| t5132a | Vif | I31N | ||||||
| t5223c | Vif | D61gac | t5223c | Vif | D61gac | |||
| g5310a | Vif | R90aga | ||||||
| g5593del | *1 | g5593del | 1 | |||||
| *2 | 2 | |||||||
| a5613g | *2 | |||||||
| 5632-5717del | *2 | |||||||
| g5865a | Tat | K12aaa | ||||||
| c6633t | Env | T138I | c6633t | Env | T138I | |||
| t6643c | Env | N141aac | ||||||
| c6652t | Env | S144agt | ||||||
| c6830a | Env | P204T | c6830a | Env | P204T | |||
| g6938a | Env | V240I | g6938a | Env | V240I | g6938a | Env | V240I |
| c7531t | Env | I437att | ||||||
| NL-ccgad-1 | NL-ccgad-6 | ||||
|---|---|---|---|---|---|
| Nt Change | Region | NS/S Change in the Region | Nt Change | Region | NS/N Change in the Region |
| g2923a | Pol-RT | R125K | |||
| g4071a | Pol-RT | A508T | |||
| g4566a | Pol-IN | V313I | |||
| c4916t | Pol-IN (SA1D2prox) | D229gat | |||
| a4940g | Pol-IN (SA1D2prox) | G237ggg | |||
| a4943g | Pol-IN (SA1D2prox) | P238ccg | a4943g | Pol-IN (SA1D2prox) | P238ccg |
| g5090a | Pol-IN | E287gaa | |||
| Vif | R17K | ||||
| a5242g | Vif | T68A | a5242g | Vif | T68A |
| a5315g | Vif | K92R | |||
| t5379c | Vif | D113gac | |||
| a5602g | Vif | T188A | |||
| Vpr | Y15C | ||||
| 5627-5651del | *1 | ||||
| g5797a | *1 | ||||
| a5820g | *1 | a5820g | Vpr | R88G | |
| g6625a | Env | K135aag | g6625a | Env | K135aag |
| g7032a | Env | R271K | g7032a | Env | R271K |
| a7180c | Env | K320N | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koma, T.; Doi, N.; Takemoto, M.; Watanabe, K.; Yamamoto, H.; Nakashima, S.; Adachi, A.; Nomaguchi, M. The Expression Level of HIV-1 Vif Is Optimized by Nucleotide Changes in the Genomic SA1D2prox Region during the Viral Adaptation Process. Viruses 2021, 13, 2079. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102079
Koma T, Doi N, Takemoto M, Watanabe K, Yamamoto H, Nakashima S, Adachi A, Nomaguchi M. The Expression Level of HIV-1 Vif Is Optimized by Nucleotide Changes in the Genomic SA1D2prox Region during the Viral Adaptation Process. Viruses. 2021; 13(10):2079. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102079
Chicago/Turabian StyleKoma, Takaaki, Naoya Doi, Mai Takemoto, Kyosuke Watanabe, Hideki Yamamoto, Satoshi Nakashima, Akio Adachi, and Masako Nomaguchi. 2021. "The Expression Level of HIV-1 Vif Is Optimized by Nucleotide Changes in the Genomic SA1D2prox Region during the Viral Adaptation Process" Viruses 13, no. 10: 2079. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102079