
Transmission of Grapevine leafroll-associated virus-1 (Ampelovirus) and Grapevine virus A (Vitivirus) by the Cottony Grape Scale, Pulvinaria vitis (Hemiptera: Coccidae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Viruses and Insects
2.2. Recipient Grapevines
2.3. Infectivity Experiments
2.4. Controlled Transmission Experiments
- First instar nymphs (L1)
- Second instar nymphs (L2)
- Inoculation access period (IAP)
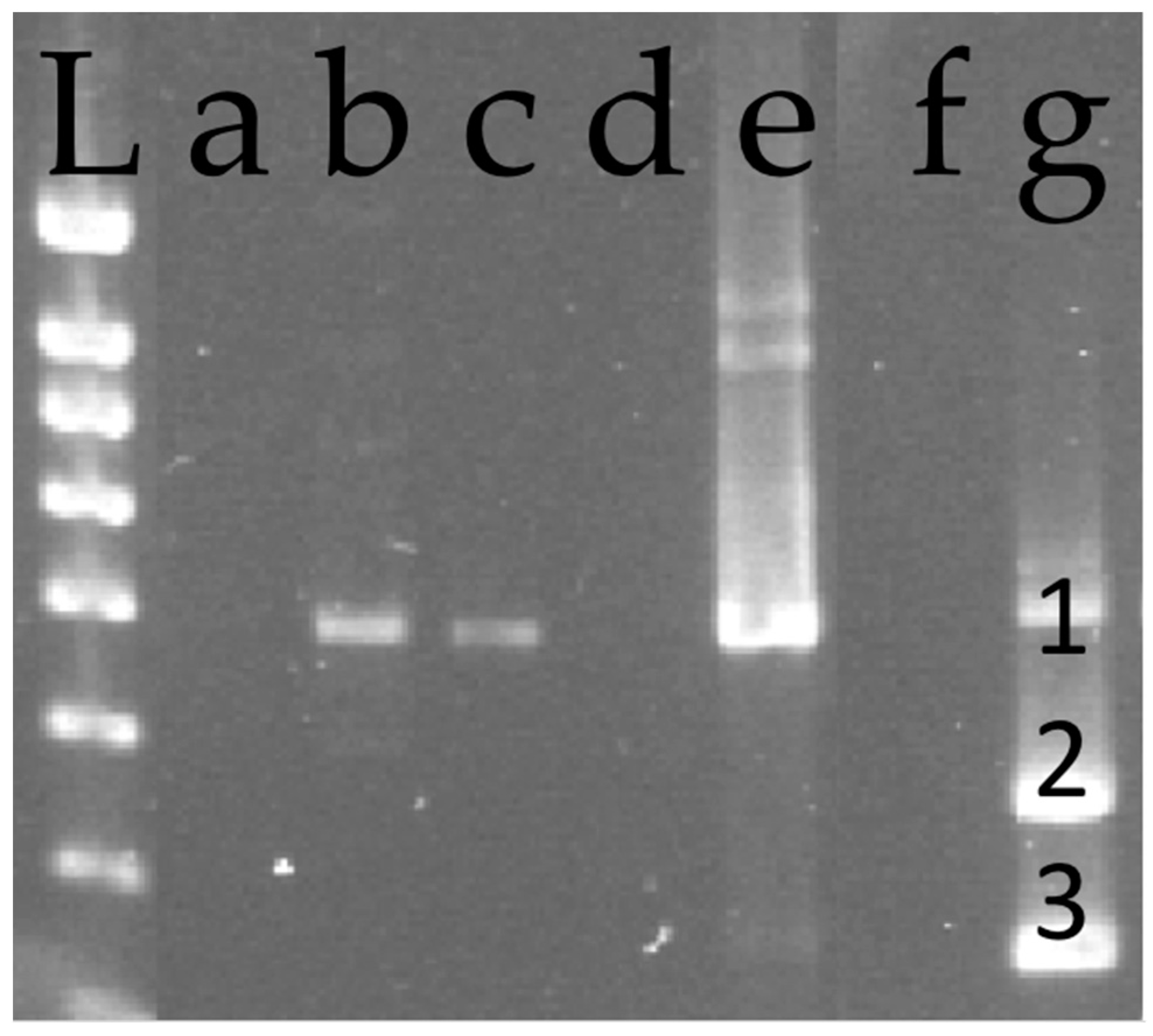
2.5. Virus Detection
3. Results
3.1. Infectivity of Natural Populations
3.2. Controlled Transmission Experiments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Balachowsky, A.; Mesnil, L. Les Insectes Nuisibles aux Plantes Cultivées. Leurs Mœurs. Leur Destruction; Établissements Busson: Paris, France, 1935; 1137p. [Google Scholar]
- Duschin, I. Investigations on the bioecology and control of the woolly vine scale (Pulvinaria vitis L.). Cercet. Agron. Mold. 1986, 19, 55–59. [Google Scholar]
- Pellizzari, G. Coccid pests of important crops: Grapevine. In World Crop Pests. Soft Scale Insects, Their Biology, Natural Enemies and Control; Ben-Dov, Y., Hodgson, C.J., Eds.; Elsevier Sciences B.V.: Amsterdam, The Netherlands; New York, NY, USA, 1997; Volume 7B, pp. 323–331. [Google Scholar]
- Masten-Milek, T.; Šimala, M.; Korić, B.; Bjeliš, M. Status of scale insects (Coccoidea), family Coccidae, on grapes in 2006 in Croatia with emphasis on rarity of second generation of Parthenolecanium corni (Bouche) and Parthenolecanium persicae (Fabricius). In Proceedings of the 8th Slovenian Conference on Plant Protection, Radenci, Slovenia, 6–7 March 2007; Plant Protection Society of Slovenia: Ljubljana, Slovenia, 2007; pp. 326–329. [Google Scholar]
- Phillips, J.H.H. Life history of Pulvinaria vitis (L.) (Hemiptera Coccoidea), the cottony scale attacking peach in Ontario. Can. Entomol. 1963, 95, 372–407. [Google Scholar] [CrossRef]
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database 2016, 2016, bav118. [Google Scholar] [CrossRef] [PubMed]
- Foldi, I.; Soria, S.J. Les cochenilles nuisibles à la vigne en Amérique du Sud (Coccoidea, Coccidae). Ann. Soc. Entomol. Fr. 1989, 25, 411–430. [Google Scholar]
- Ben-Dov, Y. A Systematic Catalogue of the Soft Scale Insects of the World (Homoptera: Coccoidea: Coccidae); Sandhill Crane Press: Gainesville, FL, USA, 1993; 536p. [Google Scholar]
- Granara de Willink, M.C. Las cochinillas blandas de la República Argentina (Homoptera: Coccoidea: Coccidae). Contrib. Entomol. Int. 1999, 3, 1–183. [Google Scholar]
- Hodgson, C.J.; Henderson, R.C. Fauna of New Zealand, No. 41. Coccidae (Insecta: Hemiptera: Coccoidea); Manaaki Whenua Press Lincoln: Canterbury, New Zealand, 2000; 259p. [Google Scholar]
- Charles, J.; Cole, L.; Walker, J.; Mundy, D. Soft Scales in New Zealand Vineyards; Research Report Commissioned by Winegrowers New Zealand Ltd. Hort Research Client Report No. 15419; Winegrowers New Zealand Ltd.: Auckland, New Zealand, 2005; 4p. [Google Scholar]
- Herrbach, E.; Le Maguet, J.; Hommay, G. Virus transmission by mealybugs and soft scales (Hemiptera, Coccoidea). In Vector-Mediated Transmission of Plant Pathogens; Brown, J.K., Ed.; American Phytopathological Society Press: St. Paul, MN, USA, 2016; pp. 147–161. [Google Scholar] [CrossRef]
- Herrbach, E.; Alliaume, A.; Prator, C.A.; Daane, K.M.; Cooper, M.L.; Almeida, R.P.P. Vector transmission of grapevine-leafroll associated viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B.Z., Martelli, G.P., Fuchs, M., Golino, D., Eds.; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017; pp. 483–503. [Google Scholar] [CrossRef]
- Belli, G.; Fortusini, A.; Casati, P.; Belli, L.; Bianco, P.A.; Prati, S. Transmission of a grapevine leafroll associated closterovirus by the scale insect Pulvinaria vitis L. Riv. Patol. Veg. 1994, 4, 105–108. [Google Scholar]
- Fortusini, A.G.; Scattini, G.; Prati, S.; Cinquanta, S.; Belli, G. Transmission of grapevine leafroll virus 1 (GLRaV-1) and grapevine virus A (GVA) by scale insects. In Proceedings of the 12th Meeting of ICVG, Lisbon, Portugal, 29 September–2 October 1997; Department of Plant Pathology, EAN/INIA: Oeiras, Portugal, 1997; pp. 121–122. [Google Scholar]
- Zorloni, A.; Prati, S.; Chiesa, S.; Bianco, P. Transmission of Grapevine leafroll-associated virus 3 by the soft scale insect Neopulvinaria innumerabilis. J. Plant Pathol. 2006, 88, S61. [Google Scholar]
- Wallingford, A.K.; Fuchs, M.F.; Martinson, T.; Hesler, S.; Loeb, G.M. Slowing the spread of Grapevine leafroll-associated viruses in commercial vineyards with insecticide control of the vector, Pseudococcus maritimus (Hemiptera: Pseudococcidae). J. Insect Sci. 2015, 15, 112. [Google Scholar] [CrossRef] [PubMed]
- Greif, C.; Walter, B. The European reference collection of grapevine virus diseases. In Sanitary Selection of the Grapevine, Proceedings of the AIR Network Conference, Colmar, France, 9 October 1997, INRA Editions; Walter, B., Ed.; INRA: Paris, France, 1997; pp. 171–181. [Google Scholar]
- Alliaume, A.; Reinbold, C.; Erhardt, M.; Beuve, M.; Hily, J.M.; Lemaire, O.; Herrbach, E. Virus preparations from the mixed-infected P70 Pinot Noir accession exhibit GLRaV-1/GVA ‘end-to-end’ particles. Arch. Virol. 2018, 163, 3149–3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beuve, M.; Hily, J.M.; Alliaume, A.; Reinbold, C.; Le Maguet, J.; Candresse, T.; Herrbach, E.; Lemaire, O. A complex virome unveiled by deep sequencing analysis of RNAs from a French Pinot Noir grapevine exhibiting strong leafroll symptoms. Arch. Virol. 2018, 163, 2937–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Maguet, J.; Beuve, M.; Herrbach, E.; Lemaire, O. Transmission of five ampeloviruses and two vitiviruses to grapevine by Phenacoccus aceris (Signoret). Phytopathology 2012, 102, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, D.; Bass, P.; Legin, R.; Walter, B. Characterization and serological detection of four closterovirus-like particles associated with leafroll disease on grapevine. J. Phytopath. 1990, 130, 205–218. [Google Scholar] [CrossRef]
- Beuve, M.; Moury, B.; Spilmont, A.S.; Sempé-Ignatovic, L.; Hemmer, C.; Lemaire, O. Viral sanitary status of declining grapevine Syrah clones and genetic diversity of Grapevine Rupestris stem pitting-associated virus. Eur. J. Plant Pathol. 2013, 135, 439–452. [Google Scholar] [CrossRef]
- Hommay, G.; Komar, V.; Lemaire, O.; Herrbach, É. Grapevine virus A transmission by larvae of Parthenolecanium corni. Eur. J. Plant Pathol. 2008, 121, 185–188. [Google Scholar] [CrossRef]
- Hommay, G.; Wiss, L.; Chadoeuf, J.; Le Maguet, J.; Beuve, M.; Herrbach, É. Gone with the wind: Aerial dispersal of Parthenolecanium corni crawlers in a newly planted grapevine plot. Ann. Appl. Biol. 2019, 174, 372–387. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Locality | Bennwihr | Kienheim | Kienheim | Nothalten | Turckheim | |
|---|---|---|---|---|---|---|
| Latitude | 48°08′18″ N | 48°41′33″ N | 48°41′37″ N | 48°21′32″ N | 48°05′42″ N | |
| Longitude | 7°19′07″ E | 7°35′32″ E | 7°35′33″ E | 7°24′40″ E | 7°16′34″ E | |
| Variety | Pinot noir | Gewurztraminer | Pinot gris | Riesling | Sylvaner | |
| source vine viruses | GLRaV−1 | 1 | 1 | 3 | ||
| GLRaV−3 | 3 | |||||
| GLRaV−1; GVA | 9 | 1 | ||||
| GLRaV−1,−3 | 1 | 2 | ||||
| GLRaV−3; GVA | 1 | |||||
| GLRaV−1,−3; GVA | 2 | 10 | ||||
| GLRaV−1,−2,−3; GVA | 1 | |||||
| number of vines tested | 6 | 1 | 1 | 26 | 1 |
| Cultivar and Accession | Transmission Rates | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soft Scale | Mean No. | Vine | Viruses in | GLRaV−1 | GLRaV−3 | GLRaV−4 Strains | GVA | Test | ||||
| Origin | /Stage | for AAP | Origin | Source Vines | 4 | 5 | 6 | 9 | Vines | |||
| Kienheim | 100 L1 | Liali Bidona Y258 | Armenia | GLRaV−1, −3; GVA | 3/4 | 0/4 | - | - | - | - | 1/4 | P115 |
| Nothalten | 18 L2 | Yshouhali Y318 | Iran | GLRaV−1, −3; GVA | 0/10 | 0/10 | - | - | - | - | 0/10 | P115 |
| Bennwihr | 50 L1 | Pinot noir P70 | France | GLRaV−1; GVA | 2/4 | - | - | - | - | - | 0/4 | P115 |
| Furdenheim | 12 L2 | Pinot noir P70 | 0/10 | - | - | - | - | - | 0/10 | P115 | ||
| Kienheim | 23 L2 | Pinot noir P70 | 7/13 | - | - | - | - | - | 5/13 | P115 | ||
| Nothalten | 100 L1 | Koudsi Y252 | Israel | GLRaV−4; GVA | - | - | 0/10 | - | - | - | nt | MO |
| Nothalten | 100 L1 | Black Monukka Y199 | USA | GLRaV−5, −6; GVA | - | - | - | 0/11 | 0/11 | - | nt | MO |
| Nothalten | 100 L1 | White Emperor Y217 | USA | GLRaV−5; GVA | - | - | - | 0/10 | - | - | nt | MO |
| Nothalten | 100 L1 | Cabernet Sauvignon Cab 119-66 | France | GLRaV−9; GVA | - | - | - | - | - | 0/11 | nt | MO |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hommay, G.; Alliaume, A.; Reinbold, C.; Herrbach, E. Transmission of Grapevine leafroll-associated virus-1 (Ampelovirus) and Grapevine virus A (Vitivirus) by the Cottony Grape Scale, Pulvinaria vitis (Hemiptera: Coccidae). Viruses 2021, 13, 2081. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102081
Hommay G, Alliaume A, Reinbold C, Herrbach E. Transmission of Grapevine leafroll-associated virus-1 (Ampelovirus) and Grapevine virus A (Vitivirus) by the Cottony Grape Scale, Pulvinaria vitis (Hemiptera: Coccidae). Viruses. 2021; 13(10):2081. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102081
Chicago/Turabian StyleHommay, Gérard, Antoine Alliaume, Catherine Reinbold, and Etienne Herrbach. 2021. "Transmission of Grapevine leafroll-associated virus-1 (Ampelovirus) and Grapevine virus A (Vitivirus) by the Cottony Grape Scale, Pulvinaria vitis (Hemiptera: Coccidae)" Viruses 13, no. 10: 2081. https://0-doi-org.brum.beds.ac.uk/10.3390/v13102081