
African Swine Fever Re-Emerging in Estonia: The Role of Seropositive Wild Boar from an Epidemiological Perspective

 , , , , and
, , , , and
Abstract
:1. Introduction
- Persistence: ASFV was present in wild boar or the environment throughout the period with no ASFV detections, but at a low prevalence (≤1% or 2%, respectively).
- Virus carrier-hypothesis: seropositive wild boar could remain infectious and, thus, kept the epidemic going.
- Reintroduction: the virus was absent for 1.5 years and newly introduced.
2. Materials and Methods
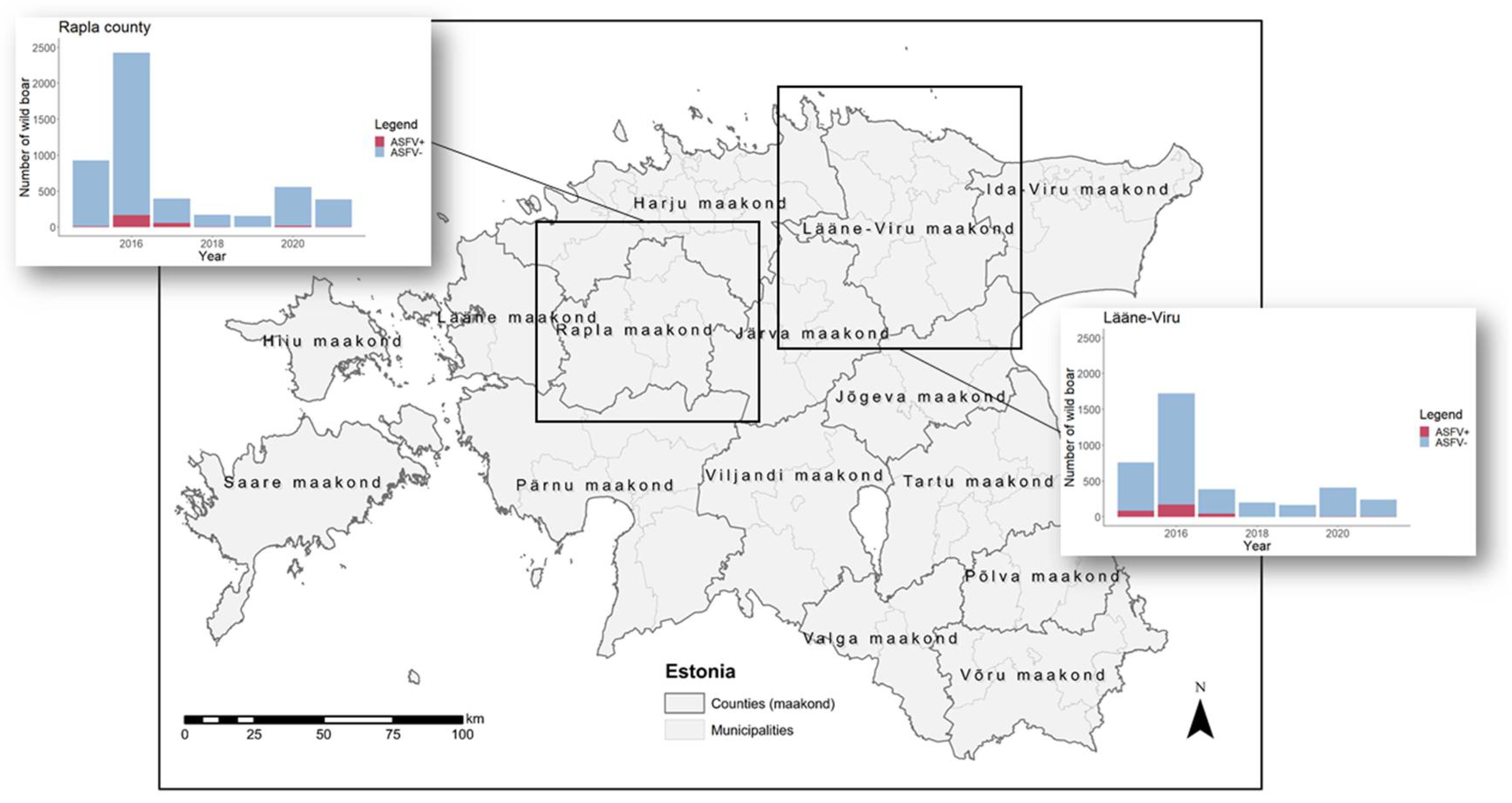
2.1. Study Areas and Data
2.2. Persistence
2.2.1. Sensitivity of the Surveillance System
- Relative risk. This measure describes the relative risk of ASF-infected wild boar in the high-risk group, relative to the risk of ASF-infected wild boar in the low-risk group. Based on the surveillance data, the risk that wild boar in the high-risk group are ASFV-infected was estimated to be 60 times higher compared to the risk that hunted wild boar were ASFV-positive (low-risk group).
- Test characteristics. The sensitivity of the test performed on individual wild boar (i.e., of the virological laboratory test) was assumed as 0.999. The specificity of the surveillance system was assumed equal to 1, expecting that all positive wild boar were followed up by a confirmatory test to ensure that they were not false positive.
- Number of animals tested in the high- and low-risk group. The surveillance sensitivity was calculated for 2018, 2019, and 2020. However, in each year, only those months during which no ASFV-positive wild boar was detected in the two study areas, were included in the calculations. The number of animals tested in the high- and low-risk group was, thus, calculated for the considered time periods, based on the surveillance data.
2.2.2. Confidence in Freedom from ASF
2.3. Virus Carrier Hypothesis
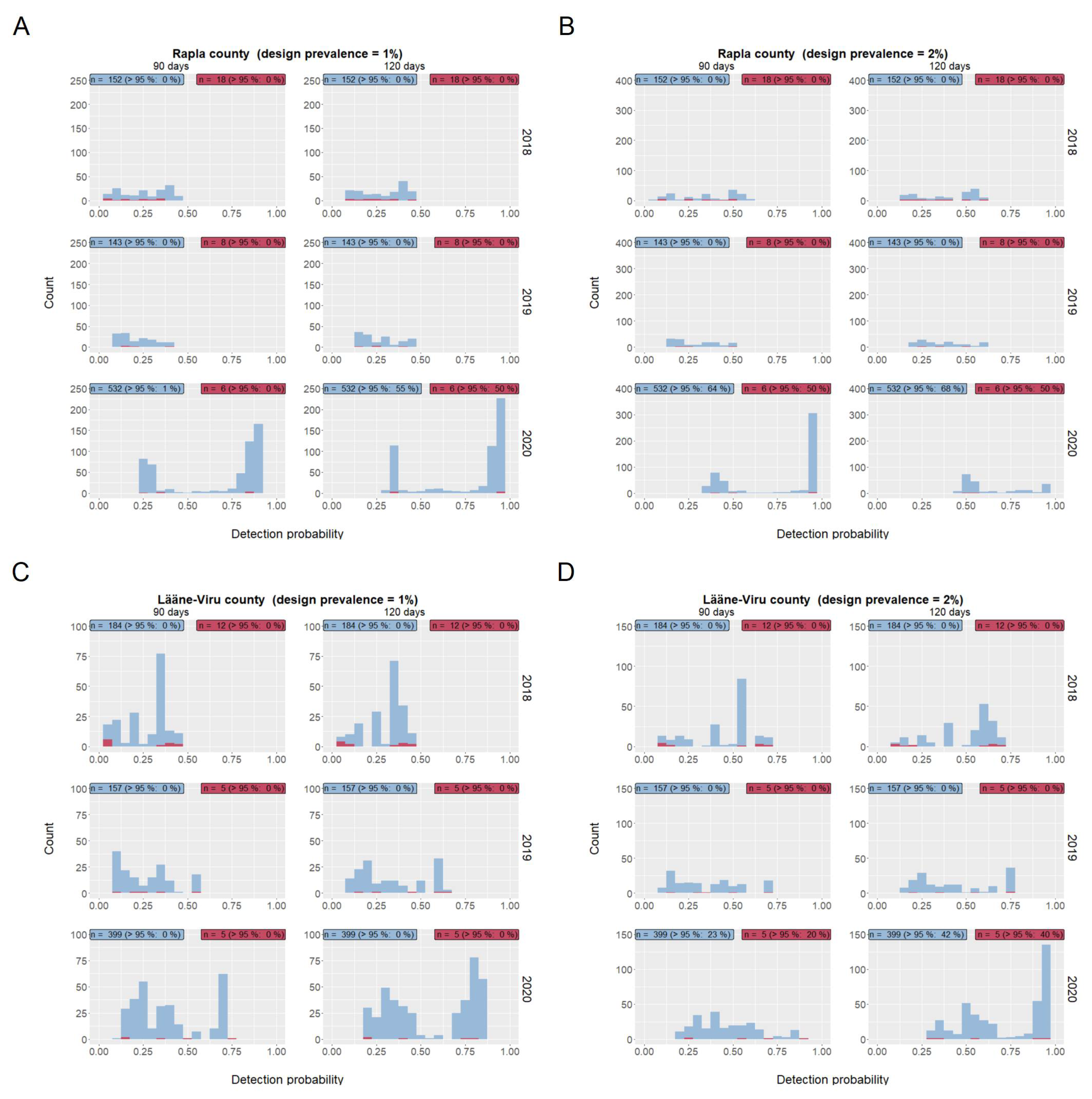
2.3.1. Detection Probability
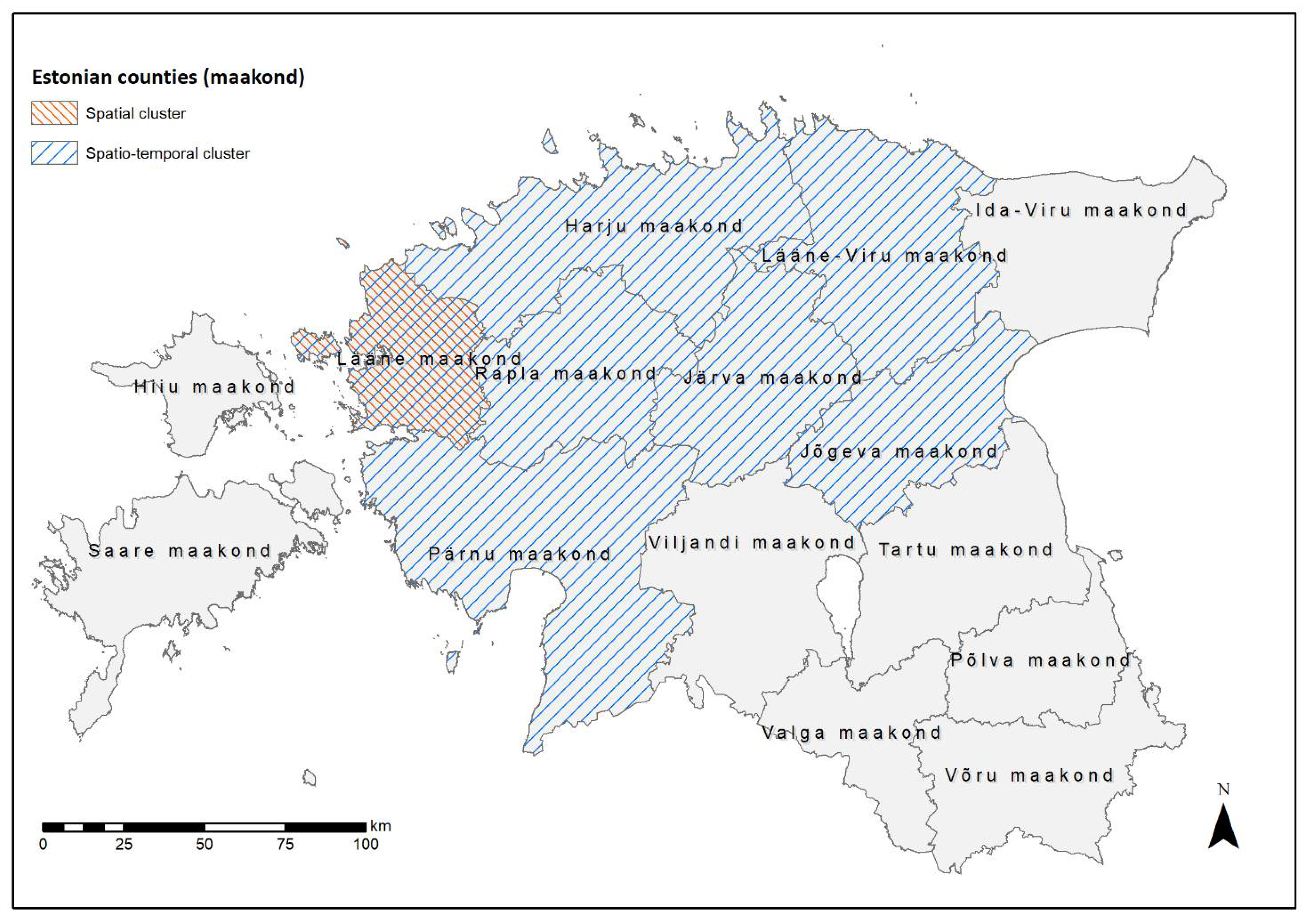
2.3.2. Cluster Analysis
2.4. Software
3. Results
3.1. Persistence
3.1.1. Sensitivity of the Surveillance System
3.1.2. Confidence in Freedom from ASF
3.2. Virus-Carrier-Hypothesis
3.2.1. Detection Probability
3.2.2. Cluster Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Montgomery, R.E. On a form of swine fever occurring in British East Africa (Kenya Colony). J. Comp. Pathol. 1921, 34, 159–191, 243–262. [Google Scholar] [CrossRef] [Green Version]
- Penrith, M.L.; Vosloo, W. Review of African swine fever: Transmission, spread and control. J. S. Afr. Vet. Assoc. 2009, 80, 58–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias, M.; Sánchez-Vizcaíno, J.; Morilla, A.; Yoon, K.-J.; Zimmerman, J. African Swine Fever Eradication: The Spanish Model. In Trends in Emerging Viral Infections of Swine; Iowa State Press: Ames, Iowa, 2002; pp. 133–139. [Google Scholar]
- Mur, L.; Atzeni, M.; Martinez-Lopez, B.; Feliziani, F.; Rolesu, S.; Sanchez-Vizcaino, J.M. Thirty-Five-Year Presence of African Swine Fever in Sardinia: History, Evolution and Risk Factors for Disease Maintenance. Transbound. Emerg. Dis. 2016, 63, E165–E177. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Franzke, K.; Beer, M. African swine fever—A review of current knowledge. Virus Res. 2020, 287, 198099. [Google Scholar] [CrossRef] [PubMed]
- Oļševskis, E.; Guberti, V.; Serzants, M.; Westergaard, J.; Gallardo, C.; Rodze, I.; Depner, K. African swine fever virus introduction into the EU in 2014: Experience of Latvia. Res. Vet. Sci. 2016, 105, 28–30. [Google Scholar] [CrossRef] [PubMed]
- Nurmoja, I.; Schulz, K.; Staubach, C.; Sauter-Louis, C.; Depner, K.; Conraths, F.J.; Viltrop, A. Development of African swine fever epidemic among wild boar in Estonia—Two different areas in the epidemiological focus. Sci. Rep. 2017, 7, 12562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautienius, A.; Grigas, J.; Pileviciene, S.; Zagrabskaite, R.; Buitkuviene, J.; Pridotkas, G.; Stankevicius, R.; Streimikyte, Z.; Salomskas, A.; Zienius, D.; et al. Prevalence and spatiotemporal distribution of African swine fever in Lithuania, 2014–2017. Virol J. 2018, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pejsak, Z.; Truszczynski, M.; Niemczuk, K.; Kozak, E.; Markowska-Daniel, I. Epidemiology of African Swine Fever in Poland since the detection of the first case. Pol. J. Vet. Sci. 2014, 17, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Chenais, E.; Ståhl, K.; Guberti, V.; Depner, K. Identification of Wild Boar–Habitat Epidemiologic Cycle in African Swine Fever Epizootic. Emerg. Infect. Dis. 2018, 24, 809–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, K.; Conraths, F.J.; Blome, S.; Staubach, C.; Sauter-Louis, C. African Swine Fever: Fast and Furious or Slow and Steady? Viruses-Basel 2019, 11, 866. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. Scientific opinion on African swine fever in wild boar. EFSA J. 2018, 16, 5344. [Google Scholar]
- Arias, M.; Jurado, C.; Gallardo, C.; Fernandez-Pinero, J.; Sanchez-Vizcaino, J.M. Gaps in African swine fever: Analysis and priorities. Transbound. Emerg. Dis. 2018, 65, 235–247. [Google Scholar] [CrossRef]
- Cukor, J.; Linda, R.; Václavek, P.; Šatrán, P.; Mahlerová, K.; Vacek, Z.; Kunca, T.; Havránek, F. Wild boar deathbed choice in relation to ASF: Are there any differences between positive and negative carcasses? Prev. Vet. Med. 2020, 177, 104943. [Google Scholar] [CrossRef]
- Morelle, K.; Bubnicki, J.; Churski, M.; Gryz, J.; Podgorski, T.; Kuijper, D.P.J. Disease-Induced Mortality Outweighs Hunting in Causing Wild Boar Population Crash After African Swine Fever Outbreak. Front. Vet. Sci. 2020, 7, 378. [Google Scholar] [CrossRef]
- Oļševskis, E.; Schulz, K.; Staubach, C.; Serzants, M.; Lamberga, K.; Pule, D.; Ozolins, J.; Conraths, F.J.; Sauter-Louis, C. African swine fever in Latvian wild boar-A step closer to elimination. Transbound. Emerg. Dis. 2020, 67, 2615–2629. [Google Scholar] [CrossRef]
- Schulz, K.; Staubach, C.; Blome, S.; Nurmoja, I.; Viltrop, A.; Conraths, F.J.; Kristian, M.; Sauter-Louis, C. How to Demonstrate Freedom from African Swine Fever in Wild Boar-Estonia as an Example. Vaccines 2020, 8, 336. [Google Scholar] [CrossRef]
- Nurmoja, I.; Mõtus, K.; Kristian, M.; Niine, T.; Schulz, K.; Depner, K.; Viltrop, A. Epidemiological analysis of the 2015–2017 African swine fever outbreaks in Estonia. Prev. Vet. Med. 2020, 181, 104556. [Google Scholar] [CrossRef]
- Putt, S.N.H.; Shaw, A.P.M.; Woods, A.J.; Tyler, L.; James, A.D. Veterinary Epidemiology and Economics in Africa; International Livestock Centre for Africa: Berkshire, UK, 1987.
- Petrov, A.; Forth, J.H.; Zani, L.; Beer, M.; Blome, S. No evidence for long-term carrier status of pigs after African swine fever virus infection. Transbound. Emerg. Dis. 2018, 65, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Eble, P.L.; Hagenaars, T.J.; Weesendorp, E.; Quak, S.; Moonen-Leusen, H.W.; Loeffen, W.L.A. Transmission of African Swine Fever Virus via carrier (survivor) pigs does occur. Vet. Microbiol. 2019, 237, 108345. [Google Scholar] [CrossRef] [PubMed]
- Schulz, K.; Staubach, C.; Blome, S.; Viltrop, A.; Nurmoja, I.; Conraths, F.J.; Sauter-Louis, C. Analysis of Estonian surveillance in wild boar suggests a decline in the incidence of African swine fever. Sci. Rep. 2019, 9, 8490. [Google Scholar] [CrossRef] [PubMed]
- Nurmoja, I.; Petrov, A.; Breidenstein, C.; Zani, L.; Forth, J.H.; Beer, M.; Kristian, M.; Viltrop, A.; Blome, S. Biological characterization of African swine fever virus genotype II strains from north-eastern Estonia in European wild boar. Transbound. Emerg. Dis. 2017, 64, 2034–2041. [Google Scholar] [CrossRef]
- Stahl, K.; Sternberg-Lewerin, S.; Blome, S.; Viltrop, A.; Penrith, M.L.; Chenais, E. Lack of evidence for long term carriers of African swine fever virus—A systematic review. Virus Res. 2019, 272, 197725. [Google Scholar] [CrossRef]
- Oh, T.; Nguyen, T.M.; Ngo, T.T.N.; Thinh, D.; Nguyen, T.T.P.; Do, L.D.; Do, D.T. Long-term follow-up of convalescent pigs and their offspring after an outbreak of acute African swine fever in Vietnam. Transbound. Emerg. Dis. 2021, 1–6. [Google Scholar] [CrossRef]
- Tignon, C.; Gallardo, C.; Iscaro, C.; Hutet, E.; Van der Stede, Y.; Kolbasov, D.; De Mia, G.M.; Le Potier, M.-F.; Bishop, R.P.; Arias, M.; et al. Development and inter-laboratory validation study of an improved new real-time PCR assay with internal control for detection and laboratory diagnosis of African swine fever virus. J. Virol. Methods 2011, 178, 161–170. [Google Scholar] [CrossRef]
- Fernandez-Pinero, J.; Gallardo, C.; Elizalde, M.; Robles, A.; Gomez, C.; Bishop, R.; Heath, L.; Couacy-Hymann, E.; Fasina, F.O.; Pelayo, V.; et al. Molecular Diagnosis of African Swine Fever by a New Real-Time PCR Using Universal Probe Library. Transbound. Emerg. Dis. 2013, 60, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Ebert, C.; Knauer, F.; Spielberger, B.; Thiele, B.; Hohmann, U. Estimating wild boar Sus scrofa population size using faecal DNA and capture-recapture modelling. Wildl. Biol. 2012, 18, 142–152. [Google Scholar] [CrossRef] [Green Version]
- Keuling, O.; Baubet, E.; Duscher, A.; Ebert, C.; Fischer, C.; Monaco, A.; Podgorski, T.; Prevot, C.; Ronnenberg, K.; Sodeikat, G.; et al. Mortality rates of wild boar Sus scrofa L. in central Europe. Eur. J. Wildl. Res. 2013, 59, 805–814. [Google Scholar] [CrossRef]
- Sergeant, E.S.G.; Important Formulae for Surveillance. AusVet Animal Health Services. Available online: http://epitools.ausvet.com.au/docs/Important_formulae_for_surveillance.pdf. (accessed on 27 July 2021).
- Cameron, A.R.; Conraths, F.J.; Fröhlich, A.; Schauer, B.; Schulz, K.; Sergeant, E.; Sonnenburg, J.; Staubach, C.R. Package of Functions for Risk-Based Surveillance. Available online: https://www.fp7-risksur.eu/sites/default/files/documents/Deliverables/RISKSUR_%28310806%29_D6.24.pdf (accessed on 27 July 2021).
- Watkins, R.E.; Martin, P.A.J.; Kelly, H.; Madin, B.; Watson, C. An evaluation of the sensitivity of acute flaccid paralysis surveillance for poliovirus infection in Australia. BMC Infect. Dis. 2009, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, A.R.; Baldock, F.C. A new probability formula for surveys to substantiate freedom from disease. Prev. Vet. Med. 1998, 34, 1–17. [Google Scholar] [CrossRef]
- Tango, T.; Takahashi, K. A flexibly shaped spatial scan statistic for detecting clusters. Int. J. Health Geogr. 2005, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Oden, N.; Jacquez, G.; Grimson, R. Adjusting Moran’s I for population density by Oden, N., Statistics in Medicine, 14, 17–26 (1995)—Reply. Stat. Med. 1998, 17, 1058–1062. [Google Scholar] [CrossRef]
- Kulldorff, M.; SaTScan. Software for the Spatial and Space-Time Scan Statistics. Version 9.6: Information Management Services. Available online: http://www.satscan.org (accessed on 27 July 2021).
- Grolemund, G.; Wickham, H. Dates and Times Made Easy with lubridate. J. Stat. Softw 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Sergeant, E. RSurveillance: Design and Analysis of Disease Surveillance Activitiess. R Package Version 0.2.1. Available online: https://CRAN.R-project.org/package=RSurveillance (accessed on 27 July 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Auguie, B. gridExtra: Miscellaneous Functions for ‘Grid’ Graphics. R Package Version 2.3. Available online: https://CRAN.R-project.org/package=gridExtra (accessed on 27 July 2021).
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots. R Package Version 0.4.0. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 27 July 2021).
- Bivand, R.; Pebesma, E.; Gomez-Rubio, V. Applied Spatial Data Analysis with R., 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Gómez-Rubio, V.; Ferrándiz-Ferragud, J.; López-Quílez, A. Detecting clusters of disease with R. J. Geogr. Syst. 2005, 7, 189–206. [Google Scholar] [CrossRef]
- Schulz, K.; Oļševskis, E.; Staubach, C.; Lamberga, K.; Seržants, M.; Cvetkova, S.; Conraths, F.J.; Sauter-Louis, C. Epidemiological evaluation of Latvian control measures for African swine fever in wild boar on the basis of surveillance data. Sci. Rep. 2019, 9, 4189. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Risk assessment of African swine fever in the south-eastern countries of Europe. EFSA J. 2019, 17, 5861. [Google Scholar] [CrossRef] [Green Version]
- Schulz, K.; Masiulis, M.; Staubach, C.; Malakauskas, A.; Pridotkas, G.; Conraths, F.J.; Sauter-Louis, C. African swine fever and its epidemiological course in Lithuanian wild boar. Viruses Submitt. 2021, 13, 1276. [Google Scholar] [CrossRef]
- Frant, M.; Lyjak, M.; Bocian, L.; Barszcz, A.; Niemczuk, K.; Wozniakowski, G. African swine fever virus (ASFV) in Poland: Prevalence in a wild boar population (2017–2018). Vet. Med. 2020, 65, 143–158. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority. ASF Exit Strategy: Providing cumulative evidence of the absence of African swine fever virus circulation in wild boar populations using standard surveillance measures. EFSA J. 2021, 19, 72. [Google Scholar]
- World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2018; pp. 1067–1068. [Google Scholar]
- Schoder, M.-E.; Tignon, M.; Linden, A.; Vervaeke, M.; Cay, A.B. Evaluation of seven commercial African swine fever virus detection kits and three Taq polymerases on 300 well-characterized field samples. J. Virol. Methods 2020, 280, 113874. [Google Scholar] [CrossRef]
- Gallardo, C.; Fernandez-Pinero, J.; Arias, M. African swine fever (ASF) diagnosis, an essential tool in the epidemiological investigation. Virus Res. 2019, 271, 197676. [Google Scholar] [CrossRef]
- Smietanka, K.; Wozniakowski, G.; Kozak, E.; Niemczuk, K.; Fraczyk, M.; Bocian, L.; Kowalczyk, A.; Pejsak, Z. African Swine Fever Epidemic, Poland, 2014-2015. Emerg. Infect. Dis. 2016, 22, 1201–1207. [Google Scholar] [CrossRef] [Green Version]
- Wozniakowski, G.; Kozak, E.; Kowalczyk, A.; Lyjak, M.; Pomorska-Mol, M.; Niemczuk, K.; Pejsak, Z. Current status of African swine fever virus in a population of wild boar in eastern Poland (2014–2015). Arch. Virol. 2016, 161, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keuling, O.; Lauterbach, K.; Stier, N.; Roth, M. Hunter feedback of individually marked wild boar Sus scrofa L.: Dispersal and efficiency of hunting in northeastern Germany. Eur. J. Wildl. Res. 2010, 56, 159–167. [Google Scholar] [CrossRef]
- Prevot, C.; Licoppe, A. Comparing red deer (Cervus elaphus, L.) and wild boar (Sus scrofa L.) dispersal patterns in southern Belgium. Eur. J. Wildl. Res. 2013, 59, 795–803. [Google Scholar] [CrossRef]
- Podgorski, T.; Lusseau, D.; Scandura, M.; Sonnichsen, L.; Jedrzejewska, B. Long-Lasting, Kin-Directed Female Interactions in a Spatially Structured Wild Boar Social Network. PLoS ONE 2014, 9, e99875. [Google Scholar]
- Gallardo, C.; Soler, A.; Nieto, R.; Sanchez, M.A.; Martins, C.; Pelayo, V.; Carrascosa, A.; Revilla, Y.; Simon, A.; Briones, V.; et al. Experimental Transmission of African Swine Fever (ASF) Low Virulent Isolate NH/P68 by Surviving Pigs. Transbound. Emerg. Dis. 2015, 62, 612–622. [Google Scholar] [CrossRef] [Green Version]
- World Organisation for Animal Health. Disease Situation. Available online: https://wahis.oie.int/#/dashboards/country-or-disease-dashboard (accessed on 15 June 2021).
- Vilem, A. (Veterinary and Food Laboratory, Tartu, Estonia). Personal communication, 2021.
- Forth, J.; Tignon, M.; Cay, A.B.; Forth, L.; Höper, D.; Blome, S.; Beer, M. Comparative Analysis of Whole-Genome Sequence of African Swine Fever Virus Belgium 2018/1. Emerg. Infect. Dis. J. 2019, 25, 1249. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Study Area | Time Period | Number of Wild Boar Investigated for ASFV | Surveillance Sensitivity (in %) | ||
|---|---|---|---|---|---|
| Hunted | Found Dead | dp = 1% | dp = 2% | ||
| Rapla County | March 2018–December 2018 | 81 | 1 | 41.5 | 68.4 |
| 2019 | 150 | 3 | 72.0 | 93.8 | |
| January 2020–July 2020 | 171 | 0 | 46.1 | 71.0 | |
| Lääne-Viru County | 2018 | 196 | 4 | 81.4 | 97.5 |
| 2019 | 163 | 1 | 56.5 | 82.6 | |
| January 2020–November 2020 | 313 | 4 | 87.8 | 98.9 | |
| Study Area | Individual Time Periods | Confidence in Freedom for Individual Time Periods (in %) | Consecutive Time Period | Cumulative Confidence in Freedom Over Consecutive Periods (in %) | ||
|---|---|---|---|---|---|---|
| dp = 1% | dp = 2% | dp = 1% | dp = 2% | |||
| Rapla County | March–December 2018 | 63.1 | 76.0 | March–December 2018 | 58.3 | 72.1 |
| 2019 | 78.1 | 94.2 | March 2018–December 2019 | 79.8 | 96.8 | |
| January–July 2020 | 65.0 | 77.5 | March 2018–July 2020 | 82.5 | 95.9 | |
| Lääne-Viru County | 2018 | 84.3 | 97.6 | January–December 2018 | 81.5 | 97.0 |
| 2019 | 69.7 | 85.2 | January 2018–December 2019 | 86.3 | 97.5 | |
| January–November 2020 | 89.2 | 98.9 | January 2018–November 2020 | 96.6 | 99.9 | |
| Time Period | Number of Index Wild Boar (Median (Minimum-Maximum)) | Days after Serological Investigation of Index Wild Boar | Proportion of Detection Probabilities above 95% | ||
|---|---|---|---|---|---|
| dp = 1% | dp = 2% | ||||
| Sero+ | 2018 | 10 (0–71) | 90 | 0 (0–21) | 0 (0–45) |
| 120 | 0 (0–27) | 0 (0–54) | |||
| 2019 | 4 (0–12) | 90 | 0 (0–0) | 0 (0–60) | |
| 120 | 0 (0–50) | 0 (0–60) | |||
| 2020 | 4 (0–17) | 90 | 0 (0–35) | 0 (0–88) | |
| 120 | 0 (0–88) | 0 (0–88) | |||
| Sero+/− | 2018 | 256 (101–1031) | 90 | 0 (0–230) | 0 (0–54) |
| 120 | 0 (0–27) | 0 (0–65) | |||
| 2019 | 244 (83–955) | 90 | 0 (0–0) | 0 (0–53) | |
| 120 | 0 (0–28) | 0 (0–64) | |||
| 2020 | 352 (137–1445) | 90 | 0 (0–39) | 0 (0–75) | |
| 120 | 0 (0–67) | 42 (0–83) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulz, K.; Schulz, J.; Staubach, C.; Blome, S.; Nurmoja, I.; Conraths, F.J.; Sauter-Louis, C.; Viltrop, A. African Swine Fever Re-Emerging in Estonia: The Role of Seropositive Wild Boar from an Epidemiological Perspective. Viruses 2021, 13, 2121. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112121
Schulz K, Schulz J, Staubach C, Blome S, Nurmoja I, Conraths FJ, Sauter-Louis C, Viltrop A. African Swine Fever Re-Emerging in Estonia: The Role of Seropositive Wild Boar from an Epidemiological Perspective. Viruses. 2021; 13(11):2121. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112121
Chicago/Turabian StyleSchulz, Katja, Jana Schulz, Christoph Staubach, Sandra Blome, Imbi Nurmoja, Franz J. Conraths, Carola Sauter-Louis, and Arvo Viltrop. 2021. "African Swine Fever Re-Emerging in Estonia: The Role of Seropositive Wild Boar from an Epidemiological Perspective" Viruses 13, no. 11: 2121. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112121