
Identification of Hepatitis E Virus Genotypes 3 and 7 in Israel: A Public Health Concern?


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Used for Identification of HEV
2.2. Screening for HEV among Human Patients
2.3. Characterization of HEV-3 and HEV-7 Genome by Population Sequencing
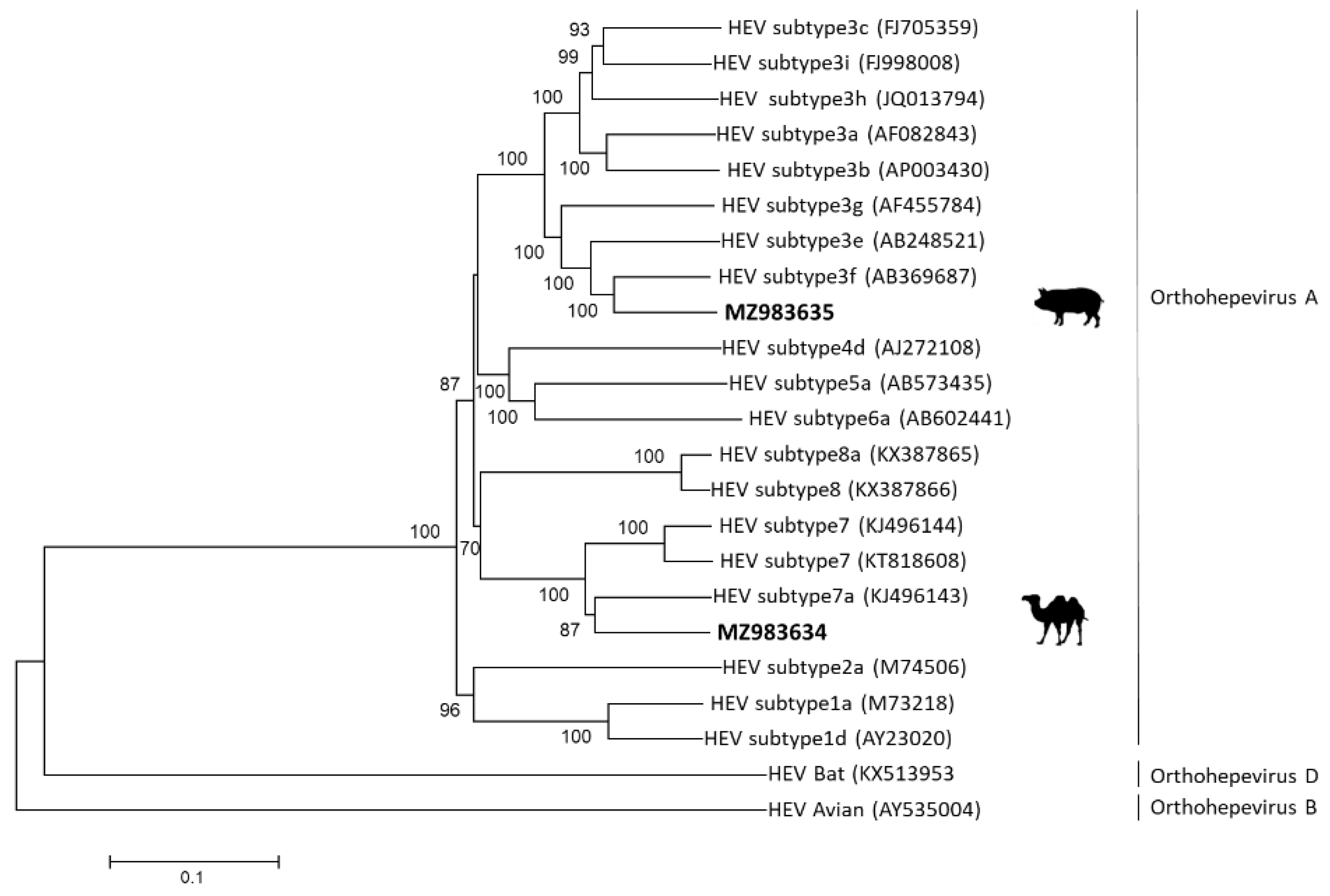
2.4. Phylogenetic Analysis of Camel and Swine-Derived HEV
2.5. Sequence Comparison
3. Results
3.1. Identification of HEV-7 in Camel Sera
3.2. Analysis of Camel- and Swine-Derived HEV Sequences
3.3. Analysis of Serum Samples from Patients with Acute Viral Hepatitis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Meng, X.J. Hepatitis E virus: Animal reservoirs and zoonotic risk. Vet. Microbiol. 2010, 140, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.H.; Tan, B.H.; Teo, E.C.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic Infection with Camelid Hepatitis E Virus in a Liver Transplant Recipient Who Regularly Consumes Camel Meat and Milk. Gastroenterology 2016, 150, 355–357.e3. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15, e04886. [Google Scholar] [CrossRef] [PubMed]
- Kamar, N.; Bendall, R.; Legrand-Abravanel, F.; Xia, N.S.; Ijaz, S.; Izopet, J.; Dalton, H.R. Hepatitis E. Lancet 2012, 379, 2477–2488. [Google Scholar] [CrossRef]
- Wang, B.; Meng, X.J. Structural and molecular biology of hepatitis E virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Lachish, T.; Tandlich, M.; Schwartz, E. Acute hepatitis in israeli travelers. J. Travel Med. 2013, 20, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Mor, O.; Bassal, R.; Michaeli, M.; Wax, M.; Ram, D.; Cohen-Ezra, O.; Cohen, D.; Mendelson, E.; Ben-Ari, Z.; Shohat, T. Prevalence of hepatitis E virus antibodies, Israel, 2009–2010. Emerg. Infect. Dis. 2015, 21, 692–694. [Google Scholar] [CrossRef] [Green Version]
- Ram, D.; Manor, Y.; Gozlan, Y.; Schwartz, E.; Ben-Ari, Z.; Mendelson, E.; Mor, O. Hepatitis E Virus Genotype 3 in Sewage and Genotype 1 in Acute Hepatitis Cases, Israel. Am. J. Trop. Med. Hyg. 2016, 95, 216–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, R.; Pozzi, P.; Wax, M.; Bar-Or, I.; Asulin, E.; Lustig, Y.; Mendelson, E.; Ben-Ari, Z.; Schwartz, E.; Mor, O. Hepatitis E in pigs in Israel: Seroprevalence, molecular characterisation and potential impact on humans. Eurosurveillance 2018, 23, 1800067. [Google Scholar] [CrossRef] [PubMed]
- A Fine of Half Million Shekels for Pigs-Farmers Who Caused Environmental Pollution. Available online: https://infospot.co.il/n/NIS_500000_for_pig_owners_who_caused_environmental_pollution (accessed on 5 May 2020).
- Bassal, R.; Wax, M.; Shirazi, R.; Shohat, T.; Cohen, D.; David, D.; Abu-Mouch, S.; Abu-Ghanem, Y.; Mendelson, E.; Ben-Ari, Z.; et al. Seroprevalence of hepatitis E virus in dromedary camels, Bedouins, Muslim Arabs and Jews in Israel, 2009–2017. Epidemiol. Infect. 2019, 147, e92. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.F.; Haqshenas, G.; Guenette, D.K.; Halbur, P.G.; Schommer, S.K.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Detection by reverse transcription-PCR and genetic characterization of field isolates of swine hepatitis E virus from pigs in different geographic regions of the United States. J. Clin. Microbiol. 2002, 40, 1326–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Yang, Y.; Nan, Y.; Ma, Z.; Yang, L.; Zhang, Y.J. The Capsid Protein of Hepatitis E Virus Inhibits Interferon Induction via Its N-terminal Arginine-Rich Motif. Viruses 2019, 11, 1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Simmonds, P.; Izopet, J.; Oliveira-Filho, E.F.; Ulrich, R.G.; Johne, R.; Koenig, M.; Jameel, S.; Harrison, T.J.; Meng, X.J.; et al. Proposed reference sequences for hepatitis E virus subtypes. J. Gen. Virol. 2016, 97, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Heller, B.; Capuccino, J.M.; Song, B.; Nimgaonkar, I.; Hrebikova, G.; Contreras, J.E.; Ploss, A. Hepatitis E virus ORF3 is a functional ion channel required for release of infectious particles. Proc. Natl. Acad. Sci. USA 2017, 114, 1147–1152. [Google Scholar] [CrossRef] [Green Version]
- Kenney, S.P.; Wentworth, J.L.; Heffron, C.L.; Meng, X.J. Replacement of the hepatitis E virus ORF3 protein PxxP motif with heterologous late domain motifs affects virus release via interaction with TSG101. Virology 2015, 486, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Tong, H.; Hoan, N.X.; Wang, B.; Wedemeyer, H.; Bock, C.T.; Velavan, T.P. Hepatitis E Virus Mutations: Functional and Clinical Relevance. EBioMedicine 2016, 11, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Wang, L.; Zhu, Y.; Geng, J.; Li, L.; Wang, X.; Bu, Q.; Zhuang, H. Analysing complete genome sequence of swine hepatitis E virus (HEV), strain CHN-XJ-SW13 isolated from Xinjiang, China: Putative host range, and disease severity determinants in HEV. Infect. Genet. Evol. 2011, 11, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Fu, H.; Wang, L.; Bu, Q.; Liu, P.; Wang, M.; Sui, Y.; Wang, X.; Zhu, Y.; Zhuang, H. Phylogenetic analysis of the full genome of rabbit hepatitis E virus (rbHEV) and molecular biologic study on the possibility of cross species transmission of rbHEV. Infect. Genet. Evol. 2011, 11, 2020–2025. [Google Scholar] [CrossRef]
- Vasickova, P.; Slany, M.; Chalupa, P.; Holub, M.; Svoboda, R.; Pavlik, I. Detection and phylogenetic characterization of human hepatitis E virus strains, Czech Republic. Emerg. Infect. Dis. 2011, 17, 917–919. [Google Scholar] [CrossRef]
- Micas, F.; Suin, V.; Péron, J.M.; Scholtes, C.; Tuaillon, E.; Vanwolleghem, T.; Bocket, L.; Lhomme, S.; Dimeglio, C.; Izopet, J.; et al. Analyses of Clinical and Biological Data for French and Belgian Immunocompetent Patients Infected with Hepatitis E Virus Genotypes 4 and 3. Front. Microbiol. 2021, 12, 822. [Google Scholar] [CrossRef] [PubMed]
- Grigas, J.; Montoya, M.; Simkute, E.; Buitkus, M.; Zagrabskaite, R.; Pautienius, A.; Razukevicius, D.; Jonaitis, L.V.; Kiudelis, G.; Skieceviciene, J.; et al. Molecular Characterization and Seroprevalence of Hepatitis E Virus in Inflammatory Bowel Disease Patients and Solid Organ Transplant Recipients. Viruses 2021, 13, 670. [Google Scholar] [CrossRef] [PubMed]
- Treagus, S.; Wright, C.; Baker-Austin, C.; Longdon, B.; Lowther, J. The Foodborne Transmission of Hepatitis E Virus to Humans. Food Environ. Virol. 2021, 13, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Hogema, B.M.; Hakze-van der Honing, R.W.; Molier, M.; Zaaijer, H.L.; van der Poel, W.H.M. Comparison of Hepatitis E Virus Sequences from Humans and Swine, the Netherlands, 1998–2015. Viruses 2021, 13, 1265. [Google Scholar] [CrossRef] [PubMed]
- Garbuglia, A.R.; Bruni, R.; Villano, U.; Vairo, F.; Lapa, D.; Madonna, E.; Picchi, G.; Binda, B.; Mariani, R.; De Paulis, F.; et al. Hepatitis E Outbreak in the Central Part of Italy Sustained by Multiple HEV Genotype 3 Strains, June–December 2019. Viruses 2021, 13, 1159. [Google Scholar] [CrossRef] [PubMed]
- Minosse, C.; Biliotti, E.; Lapa, D.; Rianda, A.; Marchili, M.; Luzzitelli, I.; Capobianchi, M.R.; McPhee, F.; Garbuglia, A.R.; D’Offizi, G. Clinical Characteristics of Acute Hepatitis E and Their Correlation with HEV Genotype 3 Subtypes in Italy. Pathogens 2020, 9, 832. [Google Scholar] [CrossRef]
- Mulder, A.C.; Kroneman, A.; Franz, E.; Vennema, H.; Tulen, A.D.; Takkinen, J.; Hofhuis, A.; Adlhoch, C. HEVnet: A One Health, collaborative, interdisciplinary network and sequence data repository for enhanced hepatitis E virus molecular typing, characterisation and epidemiological investigations. Eurosurveillance 2019, 24, 1560–7917. [Google Scholar] [CrossRef]
- Domanovic, D.; Tedder, R.; Blumel, J.; Zaaijer, H.; Gallian, P.; Niederhauser, C.; Sauleda Oliveras, S.; O’Riordan, J.; Boland, F.; Harritshoj, L.; et al. Hepatitis E and blood donation safety in selected European countries: A shift to screening? Eurosurveillance 2017, 22, 30514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Pischke, S.; Steinmann, E.; Blümel, J.; Glebe, D. Why all blood donations should be tested for hepatitis E virus (HEV). BMC Infect. Dis. 2019, 19, 541. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| (a) | |||||||
| HEV Swine | |||||||
| Genotypes/Strains (GenBank Accession) | Nucleotide Identity (%) | Amino Acid Identity (%) | |||||
| Whole Genome | ORF1 (5112 nt) | ORF2 (1980 nt) | ORF3 (366 nt) | ORF1 (1704 aa) | ORF2 (660 aa) | ORF3 (122 aa) | |
| HEV subtype 3a (AF082843) | 81.4 | 79.9 | 84.8 | 92.1 | 81.7 | 98.8 | 96.7 |
| HEV subtype 3b (APOO3430) | 80.9 | 79.4 | 84.4 | 92.7 | 81.8 | 98.8 | 97.6 |
| HEV subtype 3c (FJ705359) | 81.0 | 79.2 | 85.2 | 94.3 | 81.9 | 99.2 | 98.1 |
| HEV subtype 3e (AB248521) | 84.7 | 83.3 | 88.2 | 95.7 | 82.4 | 99.4 | 98.4 |
| HEV subtype 3f (AB369687) | 88.2 | 87.3 | 90.4 | 96.7 | 99.2 | 99.5 | 98.9 |
| HEV subtype 3f (MH504151) | 87.5 | 87.1 | 88.5 | 96.2 | 82.3 | 99.6 | 98.6 |
| HEV subtype 3g (AF455784) | 77.8 | 74.6 | 85.7 | 93.2 | 81.4 | 99.2 | 98.4 |
| HEV subtype 3 (KY232312) | 87.3 | 86.5 | 89.0 | 94.3 | 99.2 | 99.2 | 98.9 |
| HEV subtype 3 (MH450029) | 87.4 | 86.5 | 89.4 | 96.2 | 99.1 | 99.4 | 98.9 |
| (b) | |||||||
| HEV Camel | |||||||
| Genotypes/Strains (GenBank Accession) | Nucleotide Identity (%) | Amino Acid Identity (%) | |||||
| Whole Genome | ORF1 (5046 nt) | ORF2 (1980 nt) | ORF3 (339 nt) | ORF1 (1682 aa) | ORF2 (660 aa) | ORF3 (113 aa) | |
| HEV subtype 7 (KJ496144) | 85.0 | 83.9 | 87.7 | 94.7 | 81.3 | 99.3 | 98.2 |
| HEV subtype 7a (KJ496143) | 86.4 | 85.4 | 88.8 | 95.6 | 81.4 | 99.4 | 99.4 |
| a. Differences in HEV-3 Subtypes at ORF2 and ORF3 | ||||||||
| HEV-3 Subtypes | ORF2, Amino Acid | HEV-3 Subtypes | ORF3, Amino Acid | |||||
| 23 | 25 | 72 | 606 | 99 | 102 | 110 | ||
| HEV_Swine_ISRAEL | S | S | T | V | HEV_Swine_ISRAEL | H | I | S |
| HEV_AAC97210_3a | G | P | A | A | HEV_AAC97209_3a | L | T | P |
| HEV_BAB63941_3b | G | P | A | A | HEV_BAB63940_3b | L | T | P |
| HEV_ACR56298_3c | G | P | A | A | HEV_ACR56299_3c | L | I | S |
| HEV_BAE98089_3e | G | P | A | A | HEV_BAE98088_3e | L | T | P |
| HEV_ACU12605_3f | G | P | A | A | HEV_QBG59341_3f | L | T | P |
| HEV_AOG18222_3f | G | P | A | A | HEV_AOG18224_3f | H | T | P |
| HEV_AFH35001_3f | G | P | A | A | HEV_AFH35000_3f | L | T | P |
| HEV_BBB44460_3f | G | P | A | A | HEV_AFH35003_3f | L | T | P |
| HEV_BAT32887_3f | G | P | V | A | HEV_BBB44459_3f | L | T | P |
| HEV_AOG18225_3f | G | P | A | V | HEV_BAT32886_3f | L | T | P |
| HEV_QBG59342_3f | G | P | A | A | HEV_AOG18221_3f | L | T | P |
| HEV_BAG32126_3f | G | P | A | A | HEV_ACU12606_3f | L | T | P |
| HEV_AFH35004_3f | G | P | A | A | HEV_AAO67360_3g | L | T | P |
| HEV_AAO67359_3g | G | P | A | A | HEV_AEX65898_3h | L | T | P |
| HEV_AEX65899_3h | G | P | A | A | HEV_ACV66471_3i | L | T | P |
| HEV_ACV66470_3i | G | P | A | A | ||||
| b. Differences between HEV-7 Subtypes at ORF2 and ORF3 | ||||||||
| HEV-7 Subtypes | ORF2, Amino Acid | ORF3, Amino Acid | ||||||
| 35 | 68 | 70 | 113 | 149 | 92 | 108 | ||
| HEV_Camel_ISRAEL | S | S | P | I | A | A | L | |
| HEV_AHY61299_7 | G | A | S | V | S | V | L | |
| HEV_AHY61296_7a | G | A | S | V | S | V | H | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shirazi, R.; Pozzi, P.; Gozlan, Y.; Wax, M.; Lustig, Y.; Linial, M.; Mendelson, E.; Bardenstein, S.; Mor, O. Identification of Hepatitis E Virus Genotypes 3 and 7 in Israel: A Public Health Concern? Viruses 2021, 13, 2326. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112326
Shirazi R, Pozzi P, Gozlan Y, Wax M, Lustig Y, Linial M, Mendelson E, Bardenstein S, Mor O. Identification of Hepatitis E Virus Genotypes 3 and 7 in Israel: A Public Health Concern? Viruses. 2021; 13(11):2326. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112326
Chicago/Turabian StyleShirazi, Rachel, Paolo Pozzi, Yael Gozlan, Marina Wax, Yaniv Lustig, Michal Linial, Ella Mendelson, Svetlana Bardenstein, and Orna Mor. 2021. "Identification of Hepatitis E Virus Genotypes 3 and 7 in Israel: A Public Health Concern?" Viruses 13, no. 11: 2326. https://0-doi-org.brum.beds.ac.uk/10.3390/v13112326