
Emergence of a Novel Pathogenic Poxvirus Infection in the Endangered Green Sea Turtle (Chelonia mydas) Highlights a Key Threatening Process



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Capture and Tissue Sampling
2.2. Histology
2.3. Transmission Electron Microscopy (TEM)
2.4. Extraction of DNA
2.5. Library Construction and Sequencing
2.6. Genome Assembly
2.7. Genome Annotation and Bioinformatics
2.8. Comparative Genomics
2.9. Phylogenetic Analyses
3. Results
3.1. Gross Lesions
3.2. Histology and TEM
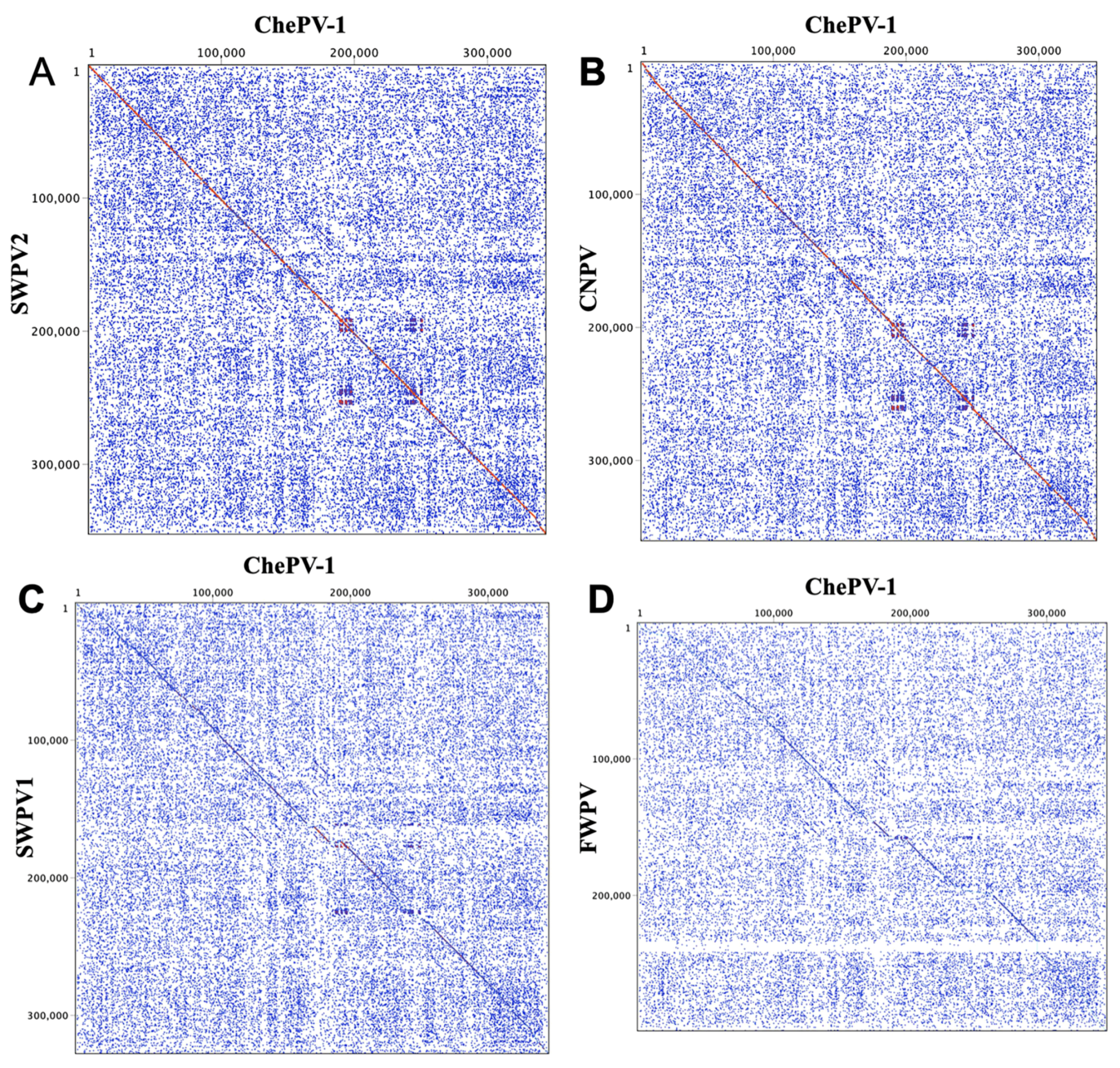
3.3. Genome Structure and Analysis of ChePV-1
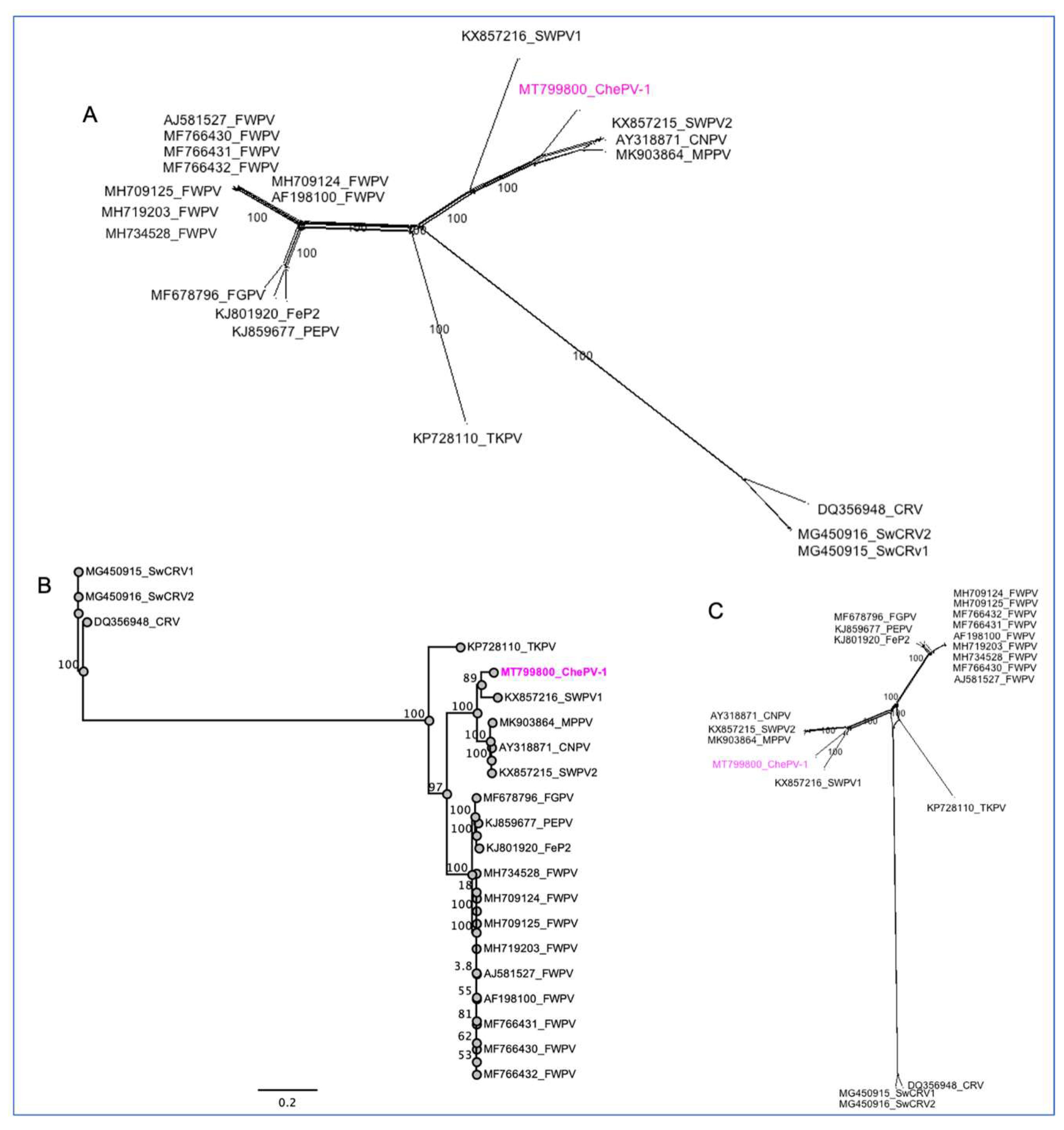
3.4. Evolutionary Relationships of ChePV-1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Australian Government Department of Agriculture, Water and the Environment: Green turtle (Chelonia mydas). Available online: https://www.environment.gov.au/marine/marine-species/marine-turtles/green (accessed on 20 August 2020).
- IUC Red List of Threatened Species: Green Turtle Chelonia mydas. Available online: https://www.iucnredlist.org/species/4615/11037468 (accessed on 30 September 2020).
- Aguirre, A.A.; Lutz, P.L. Marine turtles as sentinels of ecosystem health: Is fibropapillomatosis an indicator? EcoHealth 2004, 1, 275–283. [Google Scholar] [CrossRef]
- Limpus, J.C.; Read, A.M. The green turtle, Chelonia mydas, Queensland: A preliminary description of the population structure in a coral reef feeding ground. In Biology of Australasian Frogs and Reptiles; Grigg, G., Shine, R., Ehmann, R., Eds.; Royal Zoological Society of New South Wales: Sydney, Australia, 1985; pp. 47–52. [Google Scholar]
- Limpus, C.J.; Limpus, D.J.; Arthur, K.E.; Parmenter, C.J. Monitoring Green Turtle Population Dynamics in Shoalwater Bay 2000–2004; Research Publication No. 83; Great Barrier Reef Marine Park Authority Research Publication Series: Townsville, Australia, 2005. [Google Scholar]
- Bjorndal, K.A. Nutritional ecology of sea turtles. Copeia 1985, 1985, 736–751. [Google Scholar] [CrossRef]
- Prior, B.; Booth, D.T.; Limpus, C.J. Investigating diet and diet switching in green turtles (Chelonia mydas). Aust. J. Zool. 2016, 63, 365–375. [Google Scholar] [CrossRef]
- Bolten, B.A. Variation in sea turtle life history patterns: Neritic vs. oceanic development stages. In The Biology of Sea Turtles; Lutz, P.L., Musick, J.A., Wyneken, J., Eds.; CRC Press: Washington, DC, USA, 2003; Volume 2, pp. 243–257. [Google Scholar]
- Babcock, R.C.; Baird, M.E.; Pillans, R.; Patterson, T.; Clementson, L.A.; Haywood, M.E.; Rochester, W.; Morello, E.; Kelly, N.; Oubelkheir, K.; et al. An Integrated Study of the Gladstone Marine System; CSIRO Oceans and Atmosphere Flagship: Brisbane, Australia, 2015; p. 184. ISBN 978-1-4863-0539-1. [Google Scholar]
- Connolly, R.M.; Currie, D.R.; Danaher, K.F.; Dunning, M.; Melzer, A.; Platten, J.R.; Shearer, D.; Stratford, P.J.; Teasdale, P.R.; Vandergragt, M. Intertidal Wetlands of Port Curtis: Ecological Patterns and Processes, and Their Implications; Technical Report No. 43; CRC for Coastal Zone, Estuary and Waterway Management: Brisbane, Australia, 2006. [Google Scholar]
- Jones, K.; Burgess, G.; Budd, A.M.; Huerlimann, R.; Mashkour, N.; Ariel, E. Molecular evidence for horizontal transmission of chelonid alphaherpesvirus 5 at green turtle (Chelonia mydas) foraging grounds in Queensland, Australia. PLoS ONE 2020, 15, e0227268. [Google Scholar] [CrossRef]
- Yetsko, K.; Farrell, J.; Stammnitz, M.R.; Whitmore, L.; Whilde, J.; Eastman, C.B.; Ramia, D.R.; Thomas, R.; Krstic, A.; Linser, P.; et al. Mutational, transcriptional and viral shedding dynamics of the marine turtle fibropapillomatosis tumor epizootic. bioRxiv 2020. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Gottdenker, N.L.; Whitfield, J.; Herbst, L.; Norton, T.M.; Ritchie, B. Potential noncutaneous sites of chelonid herpesvirus 5 persistence and shedding in green sea turtles Chelonia mydas. J. Aquat. Anim. Health 2017, 29, 136–142. [Google Scholar] [CrossRef]
- Manire, C.A.; Stacy, B.A.; Kinsel, M.J.; Daniel, H.T.; Anderson, E.T.; Wellehan, J.F.X. Proliferative dermatitis in a loggerhead turtle, Caretta caretta, and a green turtle, Chelonia mydas, associated with novel papillomaviruses. Vet. Microbiol. 2008, 130, 227–237. [Google Scholar] [CrossRef]
- Mashkour, N.; Maclaine, A.; Burgess, G.W.; Ariel, E. Discovery of an Australian Chelonia mydas papillomavirus via green turtle primary cell culture and qPCR. J. Virol. Methods 2018, 258, 13–23. [Google Scholar] [CrossRef]
- Ng, T.F.F.; Manire, C.; Borrowman, K.; Langer, T.; Ehrhart, L.; Breitbart, M. Discovery of a novel single-stranded DNA virus from a sea turtle fibropapilloma by using viral metagenomics. J. Virol. 2009, 83, 2500–2509. [Google Scholar] [CrossRef] [Green Version]
- Skinner, M.A.; Buller, R.M.; Damon, I.K.; Lefkowitz, E.J.; McFadden, G.; McInnes, C.J.; Mercer, A.A.; Moyer, R.W.; Upton, C. Family Poxviridae. In Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 291–309. [Google Scholar]
- Oliveira, G.P.; Rodrigues, R.A.L.; Lima, M.T.; Drumond, B.P.; Abrahão, J.S. Poxvirus host range genes and virus-host spectrum: A critical review. Viruses 2017, 9, 331. [Google Scholar] [CrossRef] [Green Version]
- Afonso, C.L.; Tulman, E.R.; Delhon, G.; Lu, Z.; Viljoen, G.J.; Wallace, D.B.; Kutish, G.F.; Rock, D.L. Genome of crocodilepox virus. J. Virol. 2006, 80, 4978–4991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.; Isberg, R.S.; Moran, L.J.; Araujo, D.R.; Elliott, N.; Melville, L.; Beddoe, T.; Helbig, J.K. Crocodilepox virus evolutionary genomics supports observed poxvirus infection dynamics on saltwater crocodile (Crocodylus porosus). Viruses 2019, 11, 1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.; Isberg, S.R.; Milic, N.L.; Lock, P.; Helbig, K.J. Molecular characterization of the first saltwater crocodilepox virus genome sequences from the world’s largest living member of the Crocodylia. Sci. Rep. 2018, 8, 5623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orós, J.; Rodríguez, J.L.; Déniz, S.; Fernández, L.; Fernández, A. Cutaneous poxvirus-like infection in a captive Hermann’s tortoise (Testudo hermanni). Vet. Rec. 1998, 143, 508–509. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.R.; Telford, S.R. Chlamydial and poxvirus infections of circulating monocytes of a flap-necked chameleon (Chamaeleo dilepis). J. Wildl. Dis. 1990, 26, 572–577. [Google Scholar] [CrossRef]
- Frye, F.L. Infectious diseases. In Biomedical and Surgical Aspects of Captive Reptile Husbandry, 2nd ed.; Frye, F.L., Ed.; Krieger Publishing Co.: Malabar, FL, USA, 1991; p. 137. [Google Scholar]
- Sarker, S.; Roberts, H.K.; Tidd, N.; Ault, S.; Ladmore, G.; Peters, A.; Forwood, J.K.; Helbig, K.; Raidal, S.R. Molecular and microscopic characterization of a novel Eastern grey kangaroopox virus genome directly from a clinical sample. Sci. Rep. 2017, 7, 16472. [Google Scholar] [CrossRef] [Green Version]
- Sarker, S.; Das, S.; Helbig, K.; Peters, A.; Raidal, S.R. Genome sequence of an Australian strain of Canid alphaherpesvirus 1. Aust. Vet. J. 2018, 96, 24–27. [Google Scholar] [CrossRef]
- Jensen, R.H.; Mollerup, S.; Mourier, T.; Hansen, T.A.; Fridholm, H.; Nielsen, L.P.; Willerslev, E.; Hansen, A.J.; Vinner, L. Target-dependent enrichment of virions determines the reduction of high-throughput sequencing in virus discovery. PLoS ONE 2015, 10, e0122636. [Google Scholar]
- Sarker, S.; Batinovic, S.; Talukder, S.; Das, S.; Park, F.; Petrovski, S.; Forwood, J.K.; Helbig, K.J.; Raidal, S.R. Molecular characterisation of a novel pathogenic avipoxvirus from the Australian magpie (Gymnorhina tibicen). Virology 2020, 540, 1–16. [Google Scholar] [CrossRef]
- Sarker, S.; Das, S.; Lavers, J.L.; Hutton, I.; Helbig, K.; Imbery, J.; Upton, C.; Raidal, S.R. Genomic characterization of two novel pathogenic avipoxviruses isolated from pacific shearwaters (Ardenna spp.). BMC Genom. 2017, 18, 298. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tcherepanov, V.; Ehlers, A.; Upton, C. Genome Annotation Transfer Utility (GATU): Rapid annotation of viral genomes using a closely related reference genome. BMC Genom. 2006, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2013, 41, D36–D42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tusnády, G.E.; Simon, I. The HMMTOP transmembrane topology prediction server. Bioinformatics 2001, 17, 849–850. [Google Scholar] [CrossRef]
- Hofmann, K.; Stoffel, W. TMBASE—A database of membrane spanning protein segments. Biol. Chem. Hoppe-Seyler 1993, 374, 166. [Google Scholar]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Hillary, W.; Lin, S.-H.; Upton, C. Base-by-base version 2: Single nucleotide-level analysis of whole viral genome alignments. Microb. Inform. Exp. 2011, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maizel, J.V., Jr.; Lenk, R.P. Enhanced graphic matrix analysis of nucleic acid and protein sequences. PNAS 1981, 78, 7665–7669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Meth. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Huson, D.H.; Bryant, D. Application of phylogenetic networks in evolutionary studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The genome of canarypox virus. J. Virol. 2004, 78, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Offerman, K.; Carulei, O.; van der Walt, A.P.; Douglass, N.; Williamson, A.-L. The complete genome sequences of poxviruses isolated from a penguin and a pigeon in South Africa and comparison to other sequenced avipoxviruses. BMC Genom. 2014, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. The genome of fowlpox virus. J. Virol. 2000, 74, 3815–3831. [Google Scholar] [CrossRef] [Green Version]
- Joshi, L.R.; Bauermann, F.V.; Hain, K.S.; Kutish, G.F.; Armién, A.G.; Lehman, C.P.; Neiger, R.; Afonso, C.L.; Tripathy, D.N.; Diel, D.G. Detection of fowlpox virus carrying distinct genome segments of reticuloendotheliosis virus. Virus Res. 2019, 260, 53–59. [Google Scholar] [CrossRef]
- Croville, G.; Le Loc’h, G.; Zanchetta, C.; Manno, M.; Camus-Bouclainville, C.; Klopp, C.; Delverdier, M.; Lucas, M.-N.; Donnadieu, C.; Delpont, M.; et al. Rapid whole-genome based typing and surveillance of avipoxviruses using nanopore sequencing. J. Virol. Methods 2018, 261, 34–39. [Google Scholar] [CrossRef]
- Banyai, K.; Palya, V.; Denes, B.; Glavits, R.; Ivanics, E.; Horvath, B.; Farkas, S.L.; Marton, S.; Balint, A.; Gyuranecz, M.; et al. Unique genomic organization of a novel avipoxvirus detected in turkey (Meleagris gallopavo). Infect. Genet. Evol. 2015, 35, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Carulei, O.; Douglass, N.; Williamson, A.-L. Comparative analysis of avian poxvirus genomes, including a novel poxvirus from lesser flamingos (Phoenicopterus minor), highlights the lack of conservation of the central region. BMC Genom. 2017, 18, 947. [Google Scholar] [CrossRef] [PubMed]
- Weli, S.C.; Tryland, M. Avipoxviruses: Infection biology and their use as vaccine vectors. Virol. J. 2011, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, S.; Athukorala, A.; Raidal, S.R. Molecular characterisation of a novel pathogenic avipoxvirus from an Australian passerine bird, mudlark (Grallina cyanoleuca). Virology 2021, 554, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Wang, C.; Upton, C. Poxviruses: Past, present and future. Virus Res. 2006, 117, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Gyuranecz, M.; Foster, J.T.; Dán, Á.; Ip, H.S.; Egstad, K.F.; Parker, P.G.; Higashiguchi, J.M.; Skinner, M.A.; Höfle, U.; Kreizinger, Z.; et al. Worldwide phylogenetic relationship of avian poxviruses. J. Virol. 2013, 87, 4938–4951. [Google Scholar] [CrossRef] [Green Version]
- Mayr, A.; Mahnel, H. Characterization of a fow lpox virus isolated from a rhinoceros. Archiv fur die Gesamte Virusforschung 1970, 31, 51–60. [Google Scholar] [CrossRef]
- Abrahão, J.S.; Guedes, M.I.; Trindade, G.S.; Fonseca, F.G.; Campos, R.K.; Mota, B.F.; Lobato, Z.I.; Silva-Fernandes, A.T.; Rodrigues, G.O.; Lima, L.S.; et al. One more piece in the VACV ecological puzzle: Could peridomestic rodents be the link between wildlife and bovine vaccinia outbreaks in Brazil? PLoS ONE 2009, 4, e7428. [Google Scholar] [CrossRef] [Green Version]
- Hutson, C.L.; Nakazawa, Y.J.; Self, J.; Olson, V.A.; Regnery, R.L.; Braden, Z.; Weiss, S.; Malekani, J.; Jackson, E.; Tate, M.; et al. Laboratory investigations of African pouched rats (Cricetomys gambianus) as a potential reservoir host species for monkeypox virus. PLoS Negl. Trop. Dis. 2015, 9, e0004013. [Google Scholar] [CrossRef] [Green Version]
- Mauldin, M.R.; Antwerpen, M.; Emerson, G.L.; Li, Y.; Zoeller, G.; Carroll, D.S.; Meyer, H. Cowpox virus: What’s in a name? Viruses 2017, 9, 101. [Google Scholar] [CrossRef]
- Birds of Port Curtis DIWA Nationally Important Wetland. Available online: https://wetlandinfo.des.qld.gov.au/wetlands/facts-maps/wildlife/?AreaID=diwa-wetland-port-curtis&Kingdom=animals&Class=birds (accessed on 23 December 2020).
- McKenzie, L.J.; Yoshida, R.L. Seagrass-watch. In Proceedings of the Workshop for Monitoring Seagrass Habitats in the Port Curtis Coral Coast Region, Gladstone, QLD, Australia, 28–30 June 2019. [Google Scholar]
- Wildlife Unlimited. Gladstone Ports Corporation Report for Migratory Shorebird Monitoring, Port Curtis and the Curtis Coast, Annual Report—2019; Gladstone Ports Corporation: Gladstone, Australia, 2020. [Google Scholar]
- Moore, R.L.; Isberg, S.R.; Shilton, C.M.; Milic, N.L. Impact of poxvirus lesions on saltwater crocodile (Crocodylus porosus) skins. Vet. Microbiol. 2017, 211, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Yeo, G.; Wang, Y.; Chong, S.M.; Humaidi, M.; Lim, X.F.; Mailepessov, D.; Chan, S.; How, C.B.; Lin, Y.N.; Huangfu, T.; et al. Characterization of Fowlpox virus in chickens and bird-biting mosquitoes: A molecular approach to investigating Avipoxvirus transmission. J. Gen. Virol. 2019, 100, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Feare, C.J. Role of Wild birds in the spread of highly pathogenic avian influenza virus H5N1 and implications for global surveillance. Avian Dis. 2010, 54, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, T.; Yamane, M.; Kinoshita, C.; Narazaki, T.; Marshall, G.J.; Abernathy, K.J.; Miyazaki, N.; Sato, K. The feeding habit of sea turtles influences their reaction to artificial marine debris. Sci. Rep. 2016, 6, 28015. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Acronym | GenBank Accession Number | Reference |
|---|---|---|---|
| canarypox virus | CNPV | AY318871 | [45] |
| shearwaterpox virus-1 | SWPV1 | KX857216 | [29] |
| shearwaterpox virus-2 | SWPV2 | KX857215 | [29] |
| pigeonpox virus | FeP2 | KJ801920 | [46] |
| fowlpox virus | FWPV | AF198100 MF766430-32 MH709124-25 MH719203 MH734528 AJ581527 | [47,48,49] |
| turkeypox virus | TKPV | NC_028238 | [50] |
| penguinpox virus | PEPV | KJ859677 | [46] |
| flamingopox virus | FGPV | MF678796 | [51] |
| magpiepox virus | MPPV | MK903864 | [28] |
| nile crocodile poxvirus | CRV | DQ356948 | [19] |
| saltwater crocodile poxvirus-1 | SwCRV1 | MG450915 | [20,21] |
| saltwater crocodile poxvirus-2 | SwCRV2 | MG450916 | [20,21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarker, S.; Hannon, C.; Athukorala, A.; Bielefeldt-Ohmann, H. Emergence of a Novel Pathogenic Poxvirus Infection in the Endangered Green Sea Turtle (Chelonia mydas) Highlights a Key Threatening Process. Viruses 2021, 13, 219. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020219
Sarker S, Hannon C, Athukorala A, Bielefeldt-Ohmann H. Emergence of a Novel Pathogenic Poxvirus Infection in the Endangered Green Sea Turtle (Chelonia mydas) Highlights a Key Threatening Process. Viruses. 2021; 13(2):219. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020219
Chicago/Turabian StyleSarker, Subir, Christabel Hannon, Ajani Athukorala, and Helle Bielefeldt-Ohmann. 2021. "Emergence of a Novel Pathogenic Poxvirus Infection in the Endangered Green Sea Turtle (Chelonia mydas) Highlights a Key Threatening Process" Viruses 13, no. 2: 219. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020219