
The 28 Ser Amino Acid of Cucumber Mosaic Virus Movement Protein Has a Role in Symptom Formation and Plasmodesmata Localization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructions
2.2. Plant Inoculation and Symptom Detection
2.3. Analysis of Plants
2.4. Protein Analysis, SDS-PAGE and Immunoblotting
2.5. Agroinfiltration
2.6. Confocal Microscopy
3. Results
3.1. Mutations in the CMV MP
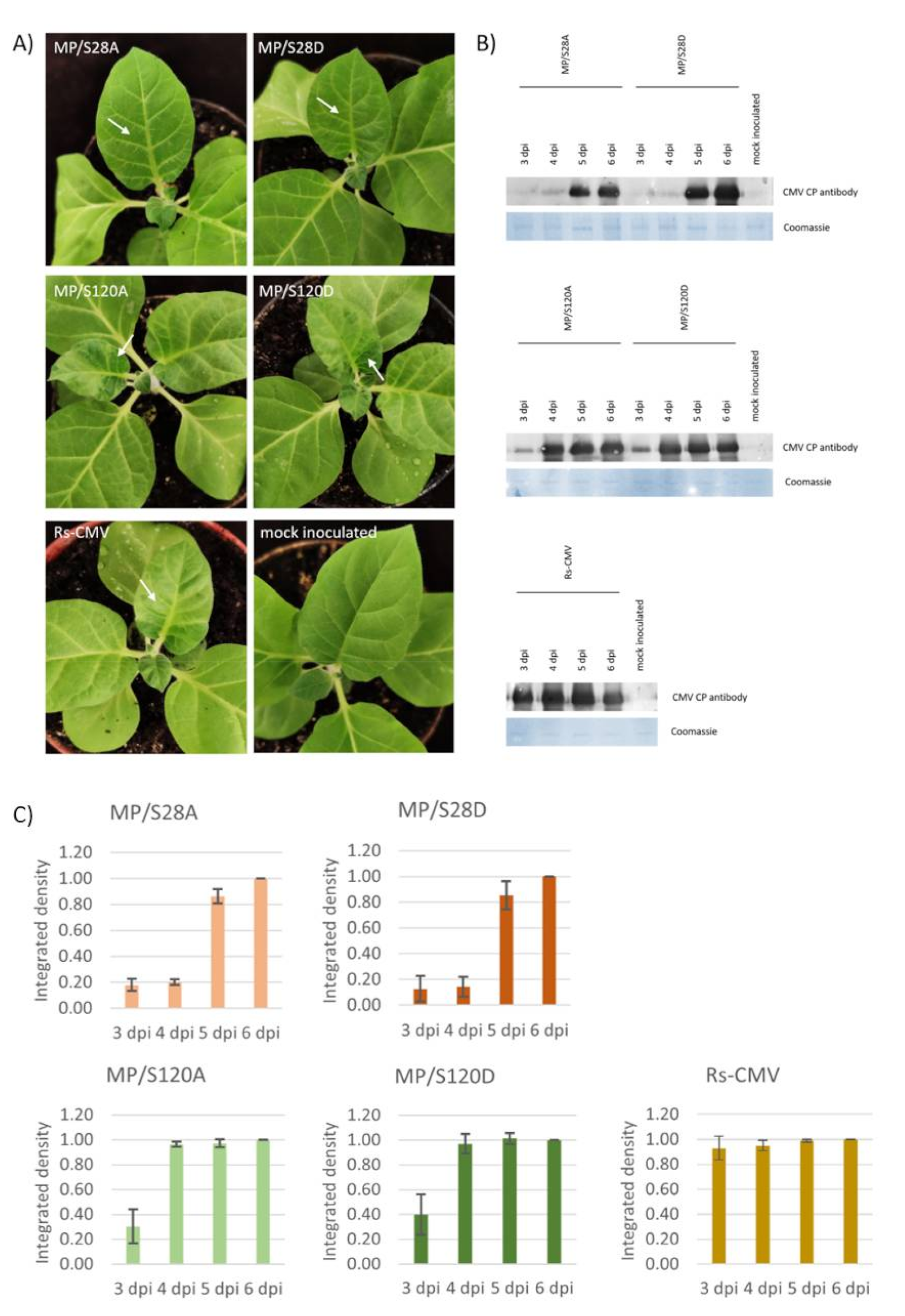
3.2. Systemic Symptom Development of MP/S28A, MP/S28D, MP/S120A and MP/S120D Mutant Viruses on Nicotiana tabacum cv. Xanthi
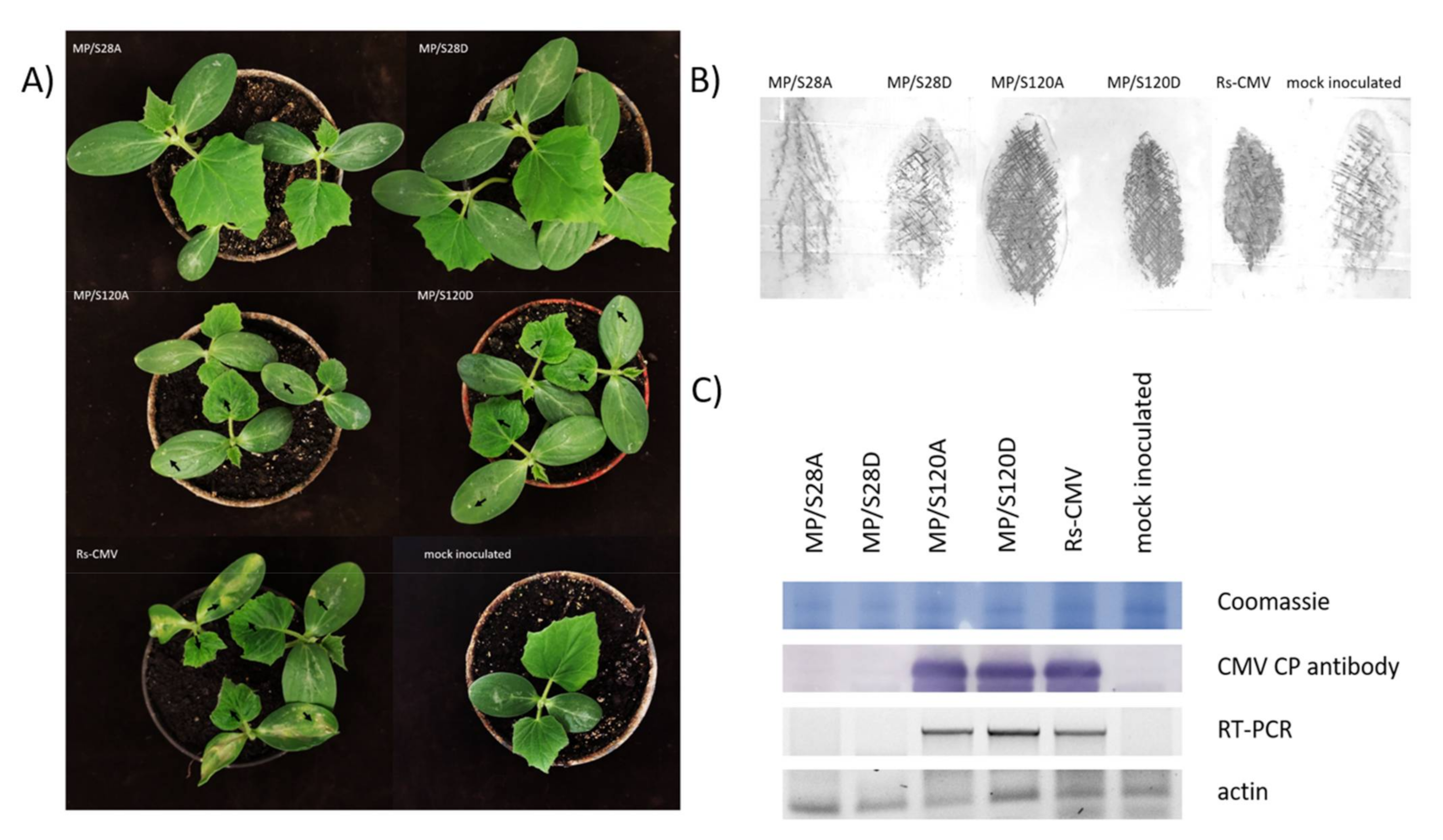
3.3. Symptom Development on Cucumis sativus
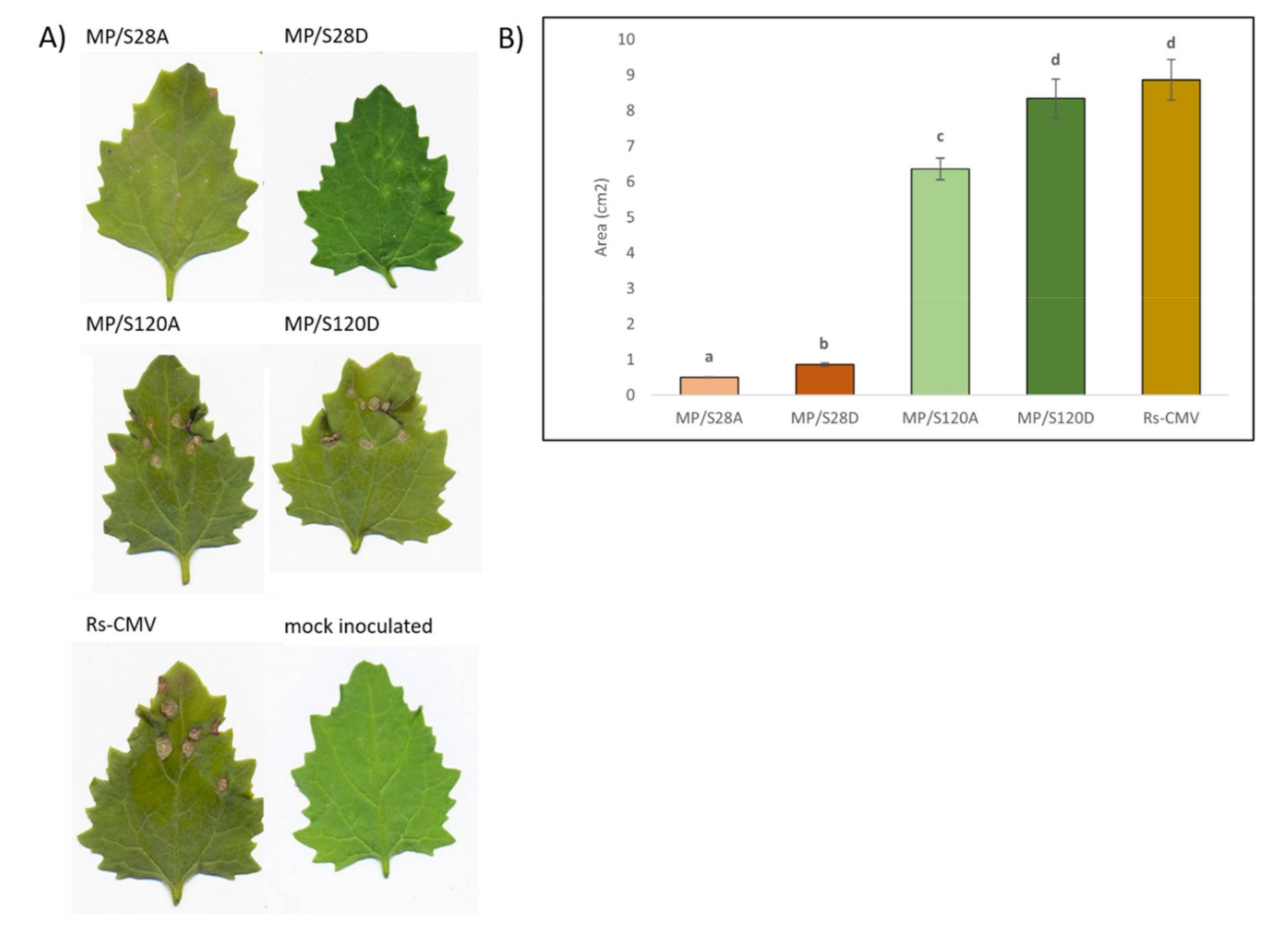
3.4. Local Lesion Induction on Chenopodium murale
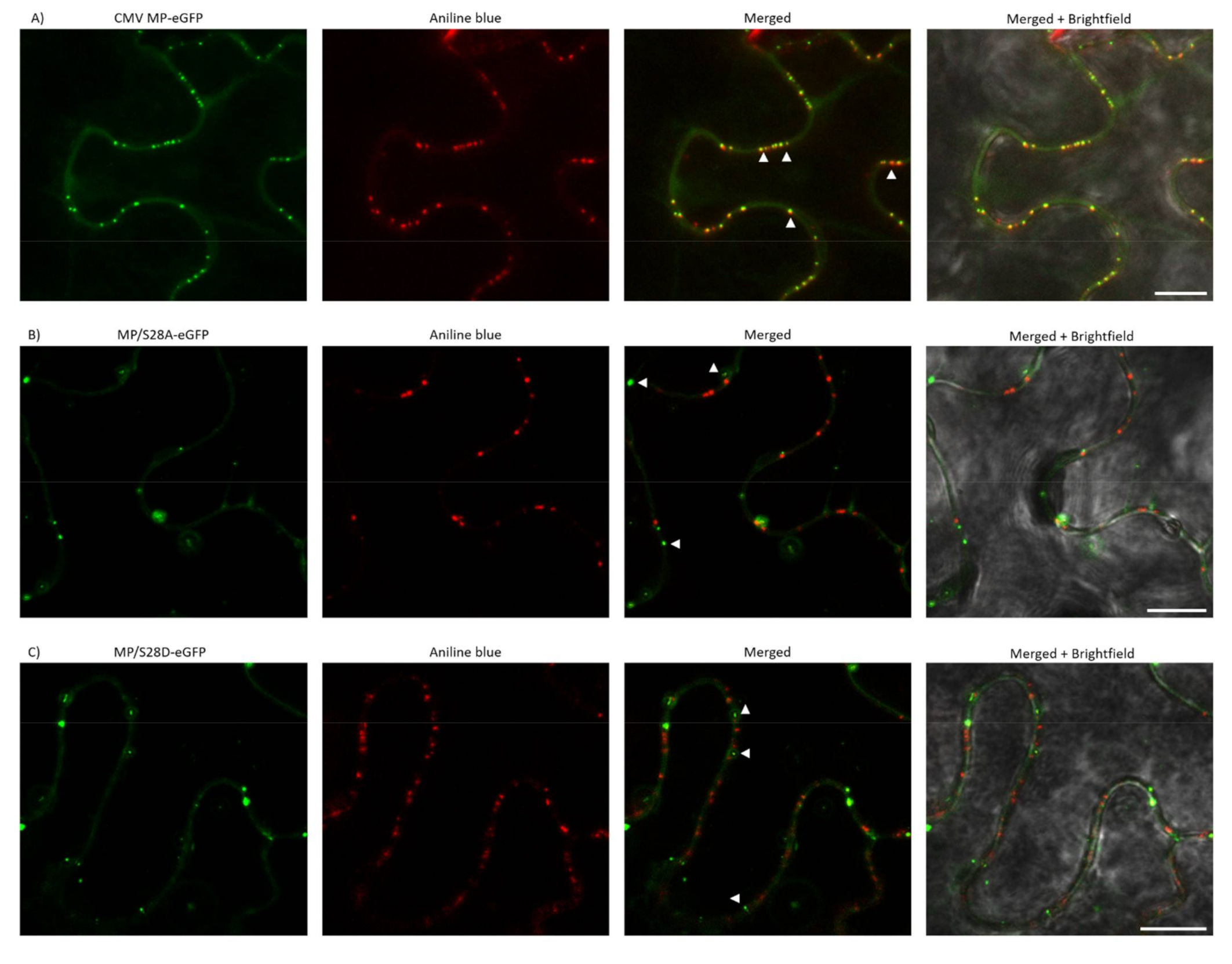
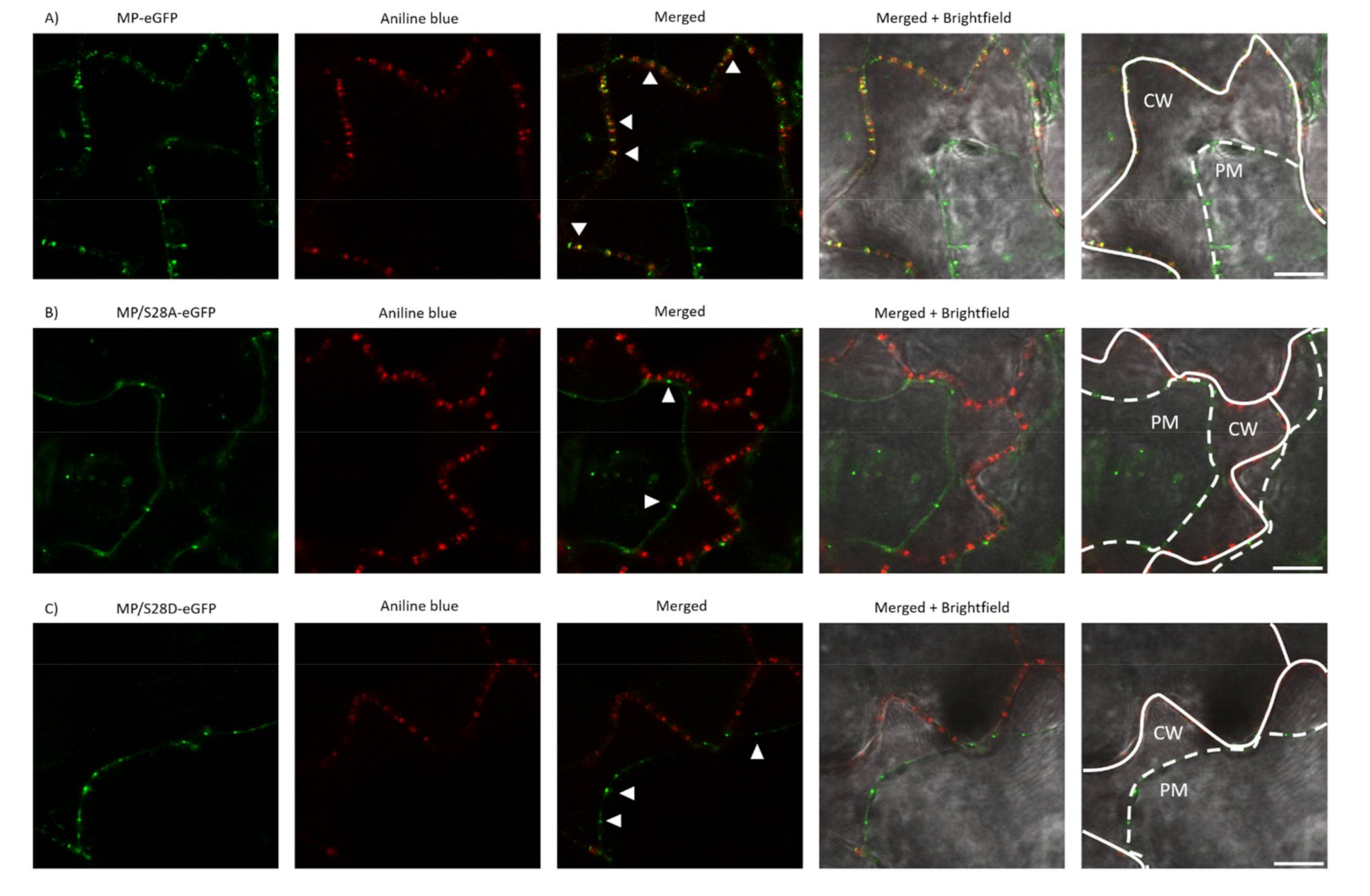
3.5. PD Localization is Affected by Mutations of Ser28 of CMV MP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Navarro, J.A.; Navarro, J.A.S.; Pallas, V. Key checkpoints in the movement of plant viruses through the host. Adv. Virus Res. 2019, 104, 1–64. [Google Scholar] [CrossRef] [PubMed]
- Melcher, U. The ‘30K’ superfamily of viral movement proteins. J. Gen. Virol. 2000, 81, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Nagano, H.; Mise, K.; Furusawa, I.; Okuno, T. Conversion in the requirement of coat protein in cell-to-cell movement mediated by the cucumber mosaic virus movement protein. J. Virol. 2001, 75, 8045–8053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aparicio, F.; Pallas, V.; Navarro, J.S. Implication of the C terminus of the Prunus necrotic ringspot virus movement protein in cell-to-cell transport and in its interaction with the coat protein. J. Gen. Virol. 2010, 91, 1865–1870. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Han, Y.; Howell, S.H. Formation of surface tubules and fluorescent foci in Arabidopsis thaliana protoplasts expressing a fusion between the green fluorescent protein and the cauliflower mosaic virus movement protein. Virology 2000, 271, 58–64. [Google Scholar] [CrossRef]
- Laporte, C.; Vetter, G.; Loudes, A.M.; Robinson, D.G.; Hillmer, S.; Garaud, C.S.; Ritzenthaler, C. Involvement of the secretory pathway and the cytoskeleton in intracellular targeting and tubule assembly of Grapevine fanleaf virus movement protein in tobacco BY-2 cells. Plant Cell 2003, 15, 2058–2075. [Google Scholar] [CrossRef] [Green Version]
- Andreev, I.A.; Kim, S.H.; Kalinina, N.O.; Rakitina, D.V.; Fitzgerald, A.G.; Palukaitis, P.; Taliansky, M.E. Molecular interactions between a plant virus movement protein and RNA: Force spectroscopy investigation. J. Mol. Biol. 2004, 339, 1041–1047. [Google Scholar] [CrossRef]
- Lecoq, H.; Desbiez, C. Viruses of Cucurbit Crops in the Mediterranean Region: An Ever-Changing picture. In Viruses and Virus Diseases of Vegetables in the Mediterranean Basin; US Vegetable Laboratory: Charleston, SC, USA, 2012; pp. 68–114. [Google Scholar]
- Porika, J.; Kodaru, A.; Prasanthi, Y.; Bharathi, B. Effect of cucumber mosaic virus (CMV) on yield and yield attributes of tomato. Res. J. Agric. Sci. 2015, 6, 443–446. [Google Scholar]
- Jones, R.A.C.; Coutts, B.A.; Latham, L.J.; McKirdy, S.J. Cucumber mosaic virus infection of chickpea stands: Temporal and spatial patterns of spread and yield limiting potential. Plant Pathol. 2008, 57, 842–853. [Google Scholar] [CrossRef]
- Chen, B.; Francki, R.I.B. Cucumovirus transmission by the aphid Myzus persicae is determined solely by the viral coat protein. J. Gen. Virol. 1990, 71, 939–944. [Google Scholar] [CrossRef]
- Kaplan, I.B.; On, A.G.; Palukaitis, P. Characterization of cucumber mosaic virus. III. Localization of sequences in the movement protein controlling systemic infection in cucurbits. Virology 1997, 230, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, K.H.; Kim, C.H.; Palukaitis, P. The coat protein of cucumber mosaic virus is a host-range determinant for infection of maize. Mol. Plant Microbe Interact. 1998, 11, 351–357. [Google Scholar] [CrossRef]
- Palukaitis, P.; Arenal, F.G. Cucumoviruses. Adv. Virus Res. 2003, 62, 241–323. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, Y.; Wang, C.; Lei, R.; Wu, Y.; Li, X.; Zhu, S. Cucumber mosaic virus coat protein induces the development of chlorotic symptoms through interacting with the chloroplast ferredoxin I protein. Sci. Rep. 2018, 8, 1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Palukaitis, P.; Park, Y.I. Phosphorylation of cucumber mosaic virus RNA polymerase 2a protein inhibits formation of replicase complex. EMBO J. 2002, 21, 2292–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucy, A.P.; Guo, H.S.; Li, W.X.; Ding, S.W. Suppression of post-transcriptional gene silencing by a plant viral protein localized in the nucleus. EMBO J. 2000, 19, 1672–1680. [Google Scholar] [CrossRef] [Green Version]
- Nemes, K.; Gellért, Á.; Almási, A.; Vági, P.; Sáray, R.; Kádár, K.; Salánki, K. Phosphorylation regulates the subcellular localization of Cucumber Mosaic Virus 2b protein. Sci. Rep. 2017, 7, 13444. [Google Scholar] [CrossRef] [Green Version]
- Nemes, K.; Gellért, Á.; Bóka, K.; Vági, P.; Salánki, K. Symptom recovery is affected by Cucumber mosaic virus coat protein phosphorylation. Virology 2019, 536, 68–77. [Google Scholar] [CrossRef]
- Li, Q.; Ryu, K.H.; Palukaitis, P. Cucumber mosaic virus-plant interactions: Identification of 3a protein sequences affecting infectivity, cell-to-cell movement, and long-distance movement. Mol. Plant Microbe Interact. 2001, 14, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Vaquero, C.; Liao, Y.C.; Nähring, J.; Fischer, R. Mapping of the RNA-binding domain of the cucumber mosaic virus movement protein. J. Gen. Virol. 1997, 78, 2095–2099. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, N.; Park, J.; Maule, A.J.; Nelson, R.S. The cysteine-histidine-rich region of the movement protein of Cucumber mosaic virus contributes to plasmodesmal targeting, zinc binding and pathogenesis. Virology 2006, 349, 396–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackman, L.M.; Boevink, P.; Cruz, S.S.; Palukaitis, P.; Oparka, K.J. The movement protein of cucumber mosaic virus traffics into sieve elements in minor veins of nicotiana clevelandii. Plant Cell 1998, 10, 525–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, H.; Okuno, T.; Mise, K.; Furusawa, I. Deletion of the C-terminal 33 amino acids of cucumber mosaic virus movement protein enables a chimeric brome mosaic virus to move from cell to cell. J. Virol. 1997, 71, 2270–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kalinina, N.O.; Andreev, I.; Ryabov, E.V.; Fitzgerald, A.G.; Taliansky, M.E.; Palukaitis, P. The C-terminal 33 amino acids of the cucumber mosaic virus 3a protein affect virus movement, RNA binding and inhibition of infection and translation. J. Gen. Virol. 2004, 85, 221–230. [Google Scholar] [CrossRef]
- Salánki, K.; Gellért, Á.; Huppert, E.; Szabó, G.N.; Balázs, E. Compatibility of the movement protein and the coat protein of cucumoviruses is required for cell-tocell movement. J. Gen. Virol. 2004, 85, 1039–1048. [Google Scholar] [CrossRef]
- Takeshita, M.; Suzuki, M.; Takanami, Y. Combination of amino acids in the 3a protein and the coat protein of cucumber mosaic virus determines symptom expression and viral spread in bottle gourd. Arch. Virol. 2001, 146, 697–711. [Google Scholar] [CrossRef]
- On, A.G.; Kaplan, I.B.; Palukaitis, P. Characterization of cucumber mosaic virus. II. Identification of movement protein sequences that influence its accumulation and systemic infection in tobacco. Virology 1996, 226, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.K.; Palukaitis, P.; Min, B.E.; Lee, M.Y.; Choi, J.K.; Ryu, K.H. Cucumber mosaic virus 2a polymerase and 3a movement proteins independently affect both virus movement and the timing of symptom development in zucchini squash. J. Gen. Virol. 2005, 86, 1213–1222. [Google Scholar] [CrossRef]
- Hwang, M.S.; Kim, K.N.; Lee, J.H.; Park, Y.I. Identification of amino acid sequences determining interaction between the cucumber mosaic virus-encoded 2a polymerase and 3a movement proteins. J. Gen. Virol. 2007, 88, 3445–3451. [Google Scholar] [CrossRef]
- Matsushita, Y.; Yoshioka, K.; Shigyo, T.; Takahashi, H.; Nyunoya, H. Phosphorylation of the movement protein of cucumber mosaic virus in transgenic tobacco plants. Virus Genes 2002, 24, 231–234. [Google Scholar] [CrossRef]
- Diveki, Z.; Salanki, K.; Balazs, E. The necrotic pathotype of the cucumber mosaic virus (CMV) ns strain is solely determined by amino acid 461 of the 1a protein. Mol. Plant Microbe Interact. 2004, 17, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Lot, H.; Marrou, J.; Qulor, J.B.; Esvan, C.H. Contribution á l’étude de virus de la mosaique de concombre (CMV)—II. Methode de purification rapide du virus. Ant. Phytopathol. 1972, 4, 25–38. [Google Scholar]
- Johansen, K.L.; Carrington, C.J. Silencing on the spot. Induction and suppression of RNA silencing in the Agrobacterium-mediated transient expression system. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Wu, S.W.; Iswanto, A.B.B.; Kumar, D.; Han, X.; Kim, J.Y. A Strategy to Validate the Role of Callose-mediated Plasmodesmal Gating in the Tropic Response. J. Vis. Exp. 2016, 110, 53513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinkel, H.; Michael, S.; Weatheritt, R.J.; Davey, N.E.; van Roey, K.; Altenberg, B.; Toedt, G.; Uyar, B.; Seiler, M.; Budd, A.; et al. ELM—The database of eukaryotic linear motifs. Nucleic Acids Res. 2012, 40, D242–D251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef]
- Canto, T.; Prior, D.A.; Hellwald, K.H.; Oparka, K.J.; Palukaitis, P. Characterization of cucumber mosaic virus. IV. Movement protein and coat protein are both essential for cell-to-cell movement of cucumber mosaic virus. Virology 1997, 237, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Bond, A.E.; Row, P.E.; Dudley, E. Post-translation modification of proteins; methodologies and applications in plant sciences. Phytochemistry 2011, 72, 975–996. [Google Scholar] [CrossRef]
- Khoury, G.A.; Baliban, R.C.; Floudas, C.A. Proteome-wide post-translational modification statistics: Frequency analysis and curation of the swiss-prot database. Sci. Rep. 2011, 1, 90. [Google Scholar] [CrossRef]
- Mochizuki, T.; Ohki, S.T. Cucumber mosaic virus: Viral genes as virulence determinants. Mol. Plant Pathol. 2012, 13, 217–225. [Google Scholar] [CrossRef]
- Berna, A.; Gafny, R.; Wolf, S.; Lucas, W.J.; Holt, C.A.; Beachy, R.N. The TMV movement protein: Role of the C-terminal 73 amino acids in subcellular localization and function. Virology 1991, 182, 682–689. [Google Scholar] [CrossRef]
- Boyko, V.; Laak, J.; Ferralli, J.; Suslova, E.; Kwon, M.-O.; Heinlein, M. Cellular Targets of Functional and Dysfunctional Mutants of Tobacco Mosaic Virus Movement Protein Fused to Green Fluorescent Protein. J. Virol. 2000, 74, 11339–11346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gafny, R.; Lapidot, M.; Berna, A.; Holt, C.A.; Deom, C.M.; Beachy, R.N. Effects of terminal deletion mutations on function of the movement protein of tobacco mosaic virus. Virology 1992, 187, 499–507. [Google Scholar] [CrossRef]
- Waigmann, E.; Chen, M.H.; Bachmaier, R.; Ghoshroy, S.; Citovsky, V. Regulation of plasmodesmal transport by phosphorylation of tobacco mosaic virus cell-to-cell movement protein. EMBO J. 2000, 19, 4875–4884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karger, E.M.; Frolova, O.Y.; Fedorova, N.V.; Baratova, L.A.; Ovchinnikova, T.V.; Susi, P.; Makinen, K.; Ronnstrand, L.; Dorokhov, Y.L.; Atabekov, J.G. Dysfunctionality of a tobacco mosaic virus movement protein mutant mimicking threonine 104 phosphorylation. J. Gen. Virol. 2003, 84, 727–732. [Google Scholar] [CrossRef]
- Kawakami, S.; Padgett, H.S.; Hosokawa, D.; Okada, Y.; Beachy, R.N.; Watanabe, Y. Phosphorylation and/or presence of serine 37 in the movement protein of tomato mosaic tobamovirus is essential for intracellular localization and stability in vivo. J. Virol. 1999, 73, 6831–6840. [Google Scholar] [CrossRef] [Green Version]
- Samuilova, O.; Santala, J.; Valkonen, J.P. Tyrosine phosphorylation of the triple gene block protein 3 regulates cell-to-cell movement and protein interactions of Potato mop-top virus. J. Virol. 2013, 87, 4313–4321. [Google Scholar] [CrossRef] [Green Version]
- Kleinow, T.; Nischang, M.; Beck, A.; Kratzer, U.; Tanwir, F.; Preiss, W.; Kepp, G.; Jeske, H. Three C-terminal phosphorylation sites in the Abutilon mosaic virus movement protein affect symptom development and viral DNA accumulation. Virology 2009, 390, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, Z.; Yuan, C.; Jin, X.; Yan, L.; Zhao, X.; Zhang, Y.; Jackson, A.O.; Wang, X.; Han, C. Phosphorylation of TGB1 by protein kinase CK2 promotes barley stripe mosaic virus movement in monocots and dicots. J. Exp. Bot. 2015, 66, 4733–4747. [Google Scholar] [CrossRef]
- Trutnyeva, K.; Bachmaier, R.; Waigmann, E. Mimicking carboxyterminal phosphorylation differentially effects subcellular distribution and cell-to-cell movement of Tobacco mosaic virus movement protein. Virology 2005, 332, 563–577. [Google Scholar] [CrossRef] [Green Version]
- Link, K.; Vogel, F.; Sonnewald, U. PD Trafficking of Potato Leaf Roll Virus Movement Protein in Arabidopsis Depends on Site-specific Protein Phosphorylation. Front. Plant Sci. 2011, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Taoka, K.I.; Yoo, B.C.; Nissan, G.B.; Kim, D.J.; Lucas, W.J. Plasmodesmal-associated protein kinase in tobacco and Arabidopsis recognizes a subset of non-cell-autonomous proteins. Plant Cell 2005, 17, 2817–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Lazarowitz, S.G.; Citovsky, V. Identification of a Functional Plasmodesmal Localization Signal in a Plant Viral Cell-To-Cell-Movement Protein. mBio 2016, 7, e02052-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, C.; Lazarowitz, S.G.; Citovsky, V. The Plasmodesmal Localization Signal of TMV MP Is Recognized by Plant Synaptotagmin SYTA. mBio 2018, 9, e01314-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáray, R.; Fábián, A.; Palkovics, L.; Salánki, K. The 28 Ser Amino Acid of Cucumber Mosaic Virus Movement Protein Has a Role in Symptom Formation and Plasmodesmata Localization. Viruses 2021, 13, 222. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020222
Sáray R, Fábián A, Palkovics L, Salánki K. The 28 Ser Amino Acid of Cucumber Mosaic Virus Movement Protein Has a Role in Symptom Formation and Plasmodesmata Localization. Viruses. 2021; 13(2):222. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020222
Chicago/Turabian StyleSáray, Réka, Attila Fábián, László Palkovics, and Katalin Salánki. 2021. "The 28 Ser Amino Acid of Cucumber Mosaic Virus Movement Protein Has a Role in Symptom Formation and Plasmodesmata Localization" Viruses 13, no. 2: 222. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020222