
Antiviral Cytokine Response in Neuroinvasive and Non-Neuroinvasive West Nile Virus Infection


 , , , ,
, , , , 

Abstract
:1. Introduction
2. Patients and Methods
2.1. Patients and Sample Collection
2.2. WNV Detection
2.3. Antiviral Cytokine Response
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, M.B.; Schaefer, T.J. West Nile Virus; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar] [PubMed]
- Sejvar, J.J. Clinical Manifestations and Outcomes of West Nile Virus Infection. Viruses 2014, 6, 606–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, N.; Varman, M. West Nile Virus Cerebellitis in a Healthy 10-Year-Old Child. Pediatr. Infect. Dis. J. 2007, 26, 767. [Google Scholar] [CrossRef] [PubMed]
- Josekutty, J.; Yeh, R.; Mathew, S.; Ene, A.; Ramessar, N.; Trinidad, J. Atypical Presentation of West Nile Virus in a Newly Diagnosed Human Immunodeficiency Virus Patient in New York City. J. Clin. Microbiol. 2013, 51, 1307–1309. [Google Scholar] [CrossRef] [Green Version]
- Castaldo, N.; Graziano, E.; Peghin, M.; Gallo, T.; D’Agaro, P.; Sartor, A.; Bove, T.; Cocconi, R.; Merlino, G.; Bassetti, M. Neuroinvasive West Nile Infection with an Unusual Clinical Presentation: A Single-Center Case Series. Trop. Med. Infect. Dis. 2020, 5, 138. [Google Scholar] [CrossRef]
- Sejvar, J.J. West Nile Virus Infection. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Bai, F.; Thompson, E.A.; Vig, P.J.S.; Leis, A.A. Current Understanding of West Nile Virus Clinical Manifestations, Immune Responses, Neuroinvasion, and Immunotherapeutic Implications. Pathogens 2019, 8, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkelmann, E.R.; Luo, H.; Wang, T. West Nile Virus Infection in the Central Nervous System. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Wang, T. Recent Advances in Understanding West Nile Virus Host Immunity and Viral Pathogenesis. F1000Research 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Peng, B.-H.; Wang, T. West Nile Virus Induced Cell Death in the Central Nervous System. Pathogens 2019, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Stonedahl, S.; Clarke, P.; Tyler, K.L. The Role of Microglia during West Nile Virus Infection of the Central Nervous System. Vaccines 2020, 8, 485. [Google Scholar] [CrossRef]
- Tobler, L.H.; Cameron, M.J.; Lanteri, M.C.; Prince, H.E.; Danesh, A.; Persad, D.; Lanciotti, R.S.; Norris, P.J.; Kelvin, D.J.; Busch, M.P. Interferon and Interferon-Induced Chemokine Expression Is Associated with Control of Acute Viremia in West Nile Virus-Infected Blood Donors. J. Infect. Dis. 2008, 198, 979–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares-Gusmao, R.; Rocha, B.C.; Sippert, E.; Lanteri, M.C.; Áñez, G.; Rios, M. Differential Pattern of Soluble Immune Markers in Asymptomatic Dengue, West Nile and Zika Virus Infections. Sci. Rep. 2019, 9, 17172. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, K.W.; Sachs, D.; Bardina, S.V.; Michlmayr, D.; Rodriguez, C.A.; Sum, J.; Foster, G.A.; Krysztof, D.; Stramer, S.L.; Lim, J.K. Differences in Early Cytokine Production Are Associated With Development of a Greater Number of Symptoms Following West Nile Virus Infection. J. Infect. Dis. 2016, 214, 634–643. [Google Scholar] [CrossRef]
- Hoffman, K.W.; Lee, J.J.; Foster, G.A.; Krysztof, D.; Stramer, S.L.; Lim, J.K. Sex differences in cytokine production following West Nile virus infection: Implications for symptom manifestation. Pathog. Dis. 2019, 77, ftz016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.N.; Hause, A.M.; Walker, C.M.; Orange, J.S.; Hasbun, R.; Murray, K.O. Evaluation of Prolonged Fatigue Post–West Nile Virus Infection and Association of Fatigue with Elevated Antiviral and Proinflammatory Cytokines. Viral Immunol. 2014, 27, 327–333. [Google Scholar] [CrossRef] [Green Version]
- Leis, A.A.; Grill, M.F.; Goodman, B.P.; Sadiq, S.B.; Sinclair, D.J.; Vig, P.J.S.; Bai, F. Tumor Necrosis Factor-Alpha Signaling May Contribute to Chronic West Nile Virus Post-Infectious Proinflammatory State. Front. Med. 2020, 7, 164. [Google Scholar] [CrossRef]
- Qian, F.; Thakar, J.; Yuan, X.; Nolan, M.; Murray, K.O.; Lee, W.T.; Wong, S.J.; Meng, H.; Fikrig, E.; Kleinstein, S.H.; et al. Immune Markers Associated with Host Susceptibility to Infection with West Nile Virus. Viral Immunol. 2014, 27, 39–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pem-Novosel, I.; Vilibic-Cavlek, T.; Gjenero-Margan, I.; Pandak, N.; Peric, L.; Barbic, L.; Listes, E.; Cvitkovic, A.; Stevanovic, V.; Savini, G. First Outbreak of West Nile Virus Neuroinvasive Disease in Humans, Croatia, 2012. Vector Borne Zoonotic Dis. 2014, 14, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Vilibic-Cavlek, T.; Kaic, B.; Barbic, L.; Pem-Novosel, I.; Slavic-Vrzic, V.; Lesnikar, V.; Kurecic-Filipovic, S.; Babic-Erceg, A.; Listes, E.; Stevanovic, V.; et al. First Evidence of Simultaneous Occurrence of West Nile Virus and Usutu Virus Neuroinvasive Disease in Humans in Croatia during the 2013 Outbreak. Infection 2014, 42, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and Molecular Epidemiology of West Nile and Usutu Virus Infections in Croatia in the “One Health” Context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Kurolt, I.C.; Krajinović, V.; Topić, A.; Kuzman, I.; Baršić, B.; Markotić, A. First molecular analysis of West Nile virus during the 2013 outbreak in Croatia. Virus Res. 2014, 189, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Barbic, L.; Mrzljak, A.; Brnic, D.; Klobucar, A.; Ilic, M.; Janev-Holcer, N.; Bogdanic, M.; Jemersic, L.; Stevanovic, V.; et al. Emerging and Neglected Viruses of Zoonotic Importance in Croatia. Pathogens 2021, 10, 73. [Google Scholar] [CrossRef]
- Konjevoda, S.; Dzelalija, B.; Canovic, S.; Pastar, Z.; Savic, V.; Tabain, I.; Barbic, L.; Peric, L.; Sabadi, D.; Stevanovic, V.; et al. West Nile Virus Retinitis in a Patient with Neuroinvasive Disease. Rev. Soc. Bras. Med. Trop. 2019, 52, e20190065. [Google Scholar] [CrossRef] [Green Version]
- Sabadi, D.; Peric, L.; Savic, V.; Rubil, I.; Baraban, V.; Tabain, I.; Barbic, L.; Duvnjak, M.; Bogdanic, M.; Stevanovic, V.; et al. Fatal Case of West Nile Encephalitis Associated with Acute Anteroseptal ST Elevation Myocardial Infarction (STEMI): A Case Report. New Microbiol. 2020, 43, 51–53. [Google Scholar]
- Santini, M.; Zupetic, I.; Viskovic, K.; Krznaric, J.; Kutlesa, M.; Krajinovic, V.; Polak, V.L.; Savic, V.; Tabain, I.; Barbic, L.; et al. Cauda Equina Arachnoiditis—A Rare Manifestation of West Nile Virus Neuroinvasive Disease: A Case Report. World J. Clin. Cases 2020, 8, 3797–3803. [Google Scholar] [CrossRef]
- Zember, S.; Vukelic, D. Opsoclonus-myoclonus syndrome caused by West Nile virus: A case report. In Proceedings of the Symposium with International Participation—Diagnosis and Surveillance of West Nile Virus Infections in the “One Health” Context, Faculty of Veterinary Medicine University of Zagreb, Zagreb, Croatia, 7 June 2019; pp. 16–17. [Google Scholar]
- European Centre for Disease Control and Prevention (ECDC). Meeting Report. Expert Consultation on West Nile Virus Infection. Stockholm. 21–22 April 2009. Available online: http://ecdc.europa.eu/en/publications/Publications/0909_MER_Expert_consultation_on_WNV.pdf (accessed on 4 January 2021).
- Tang, Y.; Anne Hapip, C.; Liu, B.; Fang, C.T. Highly Sensitive TaqMan RT-PCR Assay for Detection and Quantification of Both Lineages of West Nile Virus RNA. J. Clin. Virol. 2006, 36, 177–182. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Url, A.; Lussy, H.; Rebel-Bauder, B.; Nowotny, N. Emergence of Usutu Virus, an African Mosquito-Borne Flavivirus of the Japanese Encephalitis Virus Group, Central Europe. Emerg. Infect. Dis. 2002, 8, 652–656. [Google Scholar] [CrossRef]
- Di Gennaro, A.; Lorusso, A.; Casaccia, C.; Conte, A.; Monaco, F.; Savini, G. Serum Neutralization Assay Can Efficiently Replace Plaque Reduction Neutralization Test for Detection and Quantitation of West Nile Virus Antibodies in Human and Animal Serum Samples. Clin. Vaccine Immunol. 2014, 21, 1460–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.S.; Rughwani, P.; Kolenovic, M.; Ji, S.; Sun, B. LEGENDplex: Bead-assisted multiplex cytokine profiling by flow cytometry. Methods. Enzymol. 2019, 629, 151–176. [Google Scholar] [CrossRef] [PubMed]
- Gorenec, L.; Zidovec Lepej, S.; Grgic, I.; Planinic, A.; Iscic Bes, J.; Vince, A.; Begovac, J. The comparison of Th1, Th2, Th9, Th17 and Th22 cytokine profiles in acute and chronic HIV-1 infection. Microb. Pathog. 2016, 97, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LEGENDPlex Multi-analyte Flow Assay Kit, Human Th Cytokine Panel. Available online: https://www.biolegend.com/en-us/products/legendplex-hu-th-cytokine-panel-12-plex-wfp-v02-19471 (accessed on 2 February 2021).
- Rothaug, M.; Becker-Pauly, C.; Rose-John, S. The Role of Interleukin-6 Signaling in Nervous Tissue. Biochim. Biophys. Acta. 2016, 1863, 1218–1227. [Google Scholar] [CrossRef]
- Liba, Z.; Nohejlova, H.; Capek, V.; Krsek, P.; Sediva, A.; Kayserova, J. Utility of Chemokines CCL2, CXCL8, 10 and 13 and Interleukin 6 in the Pediatric Cohort for the Recognition of Neuroinflammation and in the Context of Traditional Cerebrospinal Fluid Neuroinflammatory Biomarkers. PLoS ONE 2019, 14, e0219987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshio, T.; Okamoto, H.; Kurasawa, K.; Dei, Y.; Hirohata, S.; Minota, S. IL-6, IL-8, IP-10, MCP-1 and G-CSF Are Significantly Increased in Cerebrospinal Fluid but Not in Sera of Patients with Central Neuropsychiatric Lupus Erythematosus. Lupus 2016, 25, 997–1003. [Google Scholar] [CrossRef]
- Al-Tamimi, Y.Z.; Bhargava, D.; Orsi, N.M.; Teraifi, A.; Cummings, M.; Ekbote, U.V.; Quinn, A.C.; Homer-Vanniasinkam, S.; Ross, S. Compartmentalisation of the Inflammatory Response Following Aneurysmal Subarachnoid Haemorrhage. Cytokine 2019, 123, 154778. [Google Scholar] [CrossRef]
- Sitati, E.M.; Diamond, M.S. CD4+ T-Cell Responses Are Required for Clearance of West Nile Virus from the Central Nervous System. J. Virol. 2006, 80, 12060–12069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brien, J.D.; Uhrlaub, J.L.; Nikolich-Zugich, J. West Nile Virus-Specific CD4 T Cells Exhibit Direct Anti-Viral Cytokine Secretion and Cytotoxicity and Are Sufficient for Antiviral Protection. J. Immunol. 2008, 181, 8568–8575. [Google Scholar] [CrossRef] [Green Version]
- Acharya, D.; Wang, P.; Paul, A.M.; Dai, J.; Gate, D.; Lowery, J.E.; Stokic, D.S.; Leis, A.A.; Flavell, R.A.; Town, T.; et al. Interleukin-17A Promotes CD8+ T Cell Cytotoxicity To Facilitate West Nile Virus Clearance. J. Virol. 2016, 91, e01529-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Bai, F.; Zenewicz, L.A.; Dai, J.; Gate, D.; Cheng, G.; Yang, L.; Qian, F.; Yuan, X.; Montgomery, R.R.; et al. IL-22 signaling contributes to West Nile encephalitis pathogenesis. PLoS ONE 2012, 7, e44153. [Google Scholar] [CrossRef] [PubMed]
- Vilibic-Cavlek, T.; Zidovec-Lepej, S.; Ledina, D.; Knezevic, S.; Savic, V.; Tabain, I.; Ivic, I.; Slavuljica, I.; Bogdanic, M.; Grgic, I.; et al. Clinical, Virological, and Immunological Findings in Patients with Toscana Neuroinvasive Disease in Croatia: Report of Three Cases. Trop. Med. Infect. Dis. 2020, 5, 144. [Google Scholar] [CrossRef]
- Günther, G.; Haglund, M.; Lindquist, L.; Forsgren, M.; Andersson, J.; Andersson, B.; Sköldenberg, B. Tick-Borne Encephalitis Is Associated with Low Levels of Interleukin-10 in Cerebrospinal Fluid. Infect. Ecol. Epidemiol. 2011, 1, 6029. [Google Scholar] [CrossRef] [Green Version]
- Bogovič, P.; Lusa, L.; Korva, M.; Pavletič, M.; Rus, K.R.; Lotrič-Furlan, S.; Avšič-Županc, T.; Strle, K.; Strle, F. Inflammatory Immune Responses in the Pathogenesis of Tick-Borne Encephalitis. J. Clin. Med. 2019, 8, 731. [Google Scholar] [CrossRef] [Green Version]
- Grygorczuk, S.; Świerzbińska, R.; Kondrusik, M.; Dunaj, J.; Czupryna, P.; Moniuszko, A.; Siemieniako, A.; Pancewicz, S. The Intrathecal Expression and Pathogenetic Role of Th17 Cytokines and CXCR2-Binding Chemokines in Tick-Borne Encephalitis. J. Neuroinflamm. 2018, 15, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, A.; Addo, M.M.; Dahlke, C. Sex Differences in Immunity: Implications for the Development of Novel Vaccines Against Emerging Pathogens. Front. Immunol. 2021, 11, 601170. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.P.; Teixeira, V.R.; Alencar-Silva, T.; Simonassi-Paiva, B.; Pereira, R.W.; Pogue, R.; Carvalho, J.L. Hallmarks of aging and immunosenescence: Connecting the dots. Cytokine Growth Factor Rev. 2021. [Google Scholar] [CrossRef]
- Yao, Y.; Montgomery, R.R. Role of Immune Aging in Susceptibility to West Nile Virus. Methods Mol. Biol. 2016, 1435, 235. [Google Scholar] [CrossRef] [Green Version]
- Kong, K.F.; Delroux, K.; Wang, X.; Qian, F.; Arjona, A.; Malawista, S.E.; Fikrig, E.; Montgomery, R.R. Dysregulation of TLR3 impairs the innate immune response to West Nile virus in the elderly. J. Virol. 2008, 82, 7613. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Characteristic | N Tested (%) | 95%CI | Median, Years (IQR) |
|---|---|---|---|
| Male | 27 (40.9) | 29.57–53.30 | 61 (53–76) |
| Female | 39 (59.1) | 46.69–70.43 | 66 (61–76) |
| Total | 66 (100) |
| Clinical Diagnosis | N Tested (%) | 95%CI | Median, Years (IQR) |
|---|---|---|---|
| WNV fever | 6 (9.1) | 4.08–19.04 | 52 (40–66) |
| Meningitis | 36 (54.5) | 42.28–66.28 | 64.5 (56.5–76) |
| Meningoencephalitis | 24 (36.4) | 25.53–48.79 | 71 (63.5–76) |
| Total | 66 (100) |
| Parameter | Median | IQR | Reference Values |
|---|---|---|---|
| Blood leukocyte count (×1012) | 9.4 | 7.2–11.2 | 3.4–9.7 |
| Cerebrospinal fluid (CSF) leukocyte count | 484 | 95–736 | <5 |
| CSF proteins (g/L) | 0.945 | 0.691–1.300 | 0.17–0.37 |
| CSF Neutrophils (%) | 47 | 20–58 | |
| CSF Lymphocytes (%) | 53 | 42–76 |
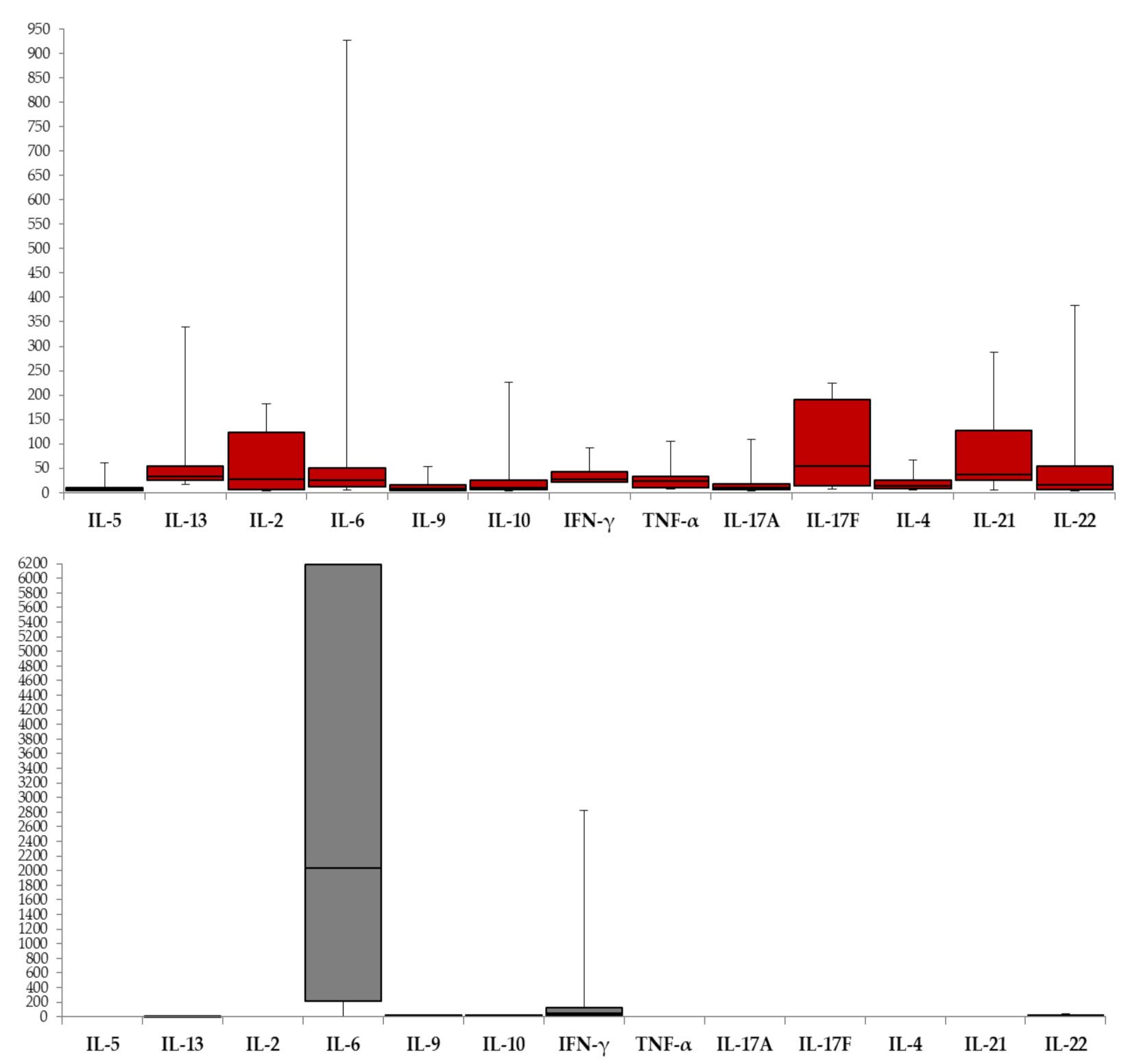
| Cytokine | WNV Serum (n = 66) | WNV CSF (n = 37) | p | ||||
|---|---|---|---|---|---|---|---|
| N Positive (%) | Median (pg/mL) | IQR | N Positive (%) | Median (pg/mL) | IQR | ||
| IL-5 | 32 (48.5) | 6.70 | 4.91–10.42 | 1 (2.6) | NA | NA | NA |
| IL-13 | 11 (16.7) | 33.26 | 24.65–54.6 | 3 (7.9) | 9.34 | 2.41–9.34 | NA |
| IL-2 | 8 (12.1) | 27.48 | 5.70–123.71 | 0 | NA | NA | NA |
| IL-6 | 61 (92.4) | 24.48 | 11.93–49.81 | 35 (92.1) | 2036.71 | 213.82–6190.5 | <0.001 |
| IL-9 | 24 (36.4) | 7.70 | 4.38–15.51 | 25 (65.8) | 5.34 | 3.57–8.35 | 0.25 |
| IL-10 | 18 (27.3) | 10.74 | 5.75–25.18 | 17 (44.7) | 5.94 | 4.06–10.54 | 0.62 |
| IFN-γ | 15 (22.7) | 27.22 | 20.95–41.82 | 27 (71.1) | 49.59 | 23.07–127.37 | 1.00 |
| TNF-α | 17 (25.8) | 22.69 | 9.99–32.35 | 0 | NA | NA | NA |
| IL-17A | 38 (57.6) | 9.44 | 6.11–17.42 | 0 | NA | NA | NA |
| IL-17F | 12 (18.2) | 53.84 | 13.68–190.17 | 1 (2.6) | NA | NA | NA |
| IL-4 | 16 (24.2) | 13.68 | 8.17–26.11 | 1 (2.6) | NA | NA | NA |
| IL-21 | 17 (25.8) | 36.56 | 24.89–127.32 | 0 | NA | NA | NA |
| IL-22 | 18 (27.3) | 15.75 | 6.37–53.44 | 20 (52.6) | 5.58 | 3.62–11.86 | 0.07 |
| Sample | Cytokine (pg/mL) | Age (Years) | Days after Disease Onset | Clinical Diagnosis | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| rho | p | p adj. | rho | p | p adj. | rho | p | p adj. | ||
| Serum | IL-5 | 0.195 | 0.411 | 1 | −0.454 | 0.044 | 0.266 | 0.511 | 0.021 | 0.128 |
| IL-13 | 0.464 | 0.354 | 1 | −0.058 | 0.913 | 1 | 0.488 | 0.326 | 1 | |
| IL-2 | −0.342 | 0.452 | 1 | −0.273 | 0.554 | 1 | 0.144 | 0.757 | 1 | |
| IL-6 | 0.407 | 0.006 | 0.036 | 0.111 | 0.471 | 1 | 0.279 | 0.066 | 0.396 | |
| IL-9 | 0.000 | 1 | 1 | −0.387 | 0.195 | 1 | 0.290 | 0.336 | 1 | |
| IL-10 | 0.063 | 0.845 | 1 | 0.239 | 0.456 | 1 | 0.190 | 0.553 | 1 | |
| IFN-γ | 0.207 | 0.593 | 1 | −0.219 | 0.571 | 1 | 0.367 | 0.332 | 1 | |
| TNF-α | −0.551 | 0.063 | 0.379 | −0.228 | 0.476 | 1 | 0.235 | 0.461 | 1 | |
| IL-17A | 0.079 | 0.692 | 1 | 0.116 | 0.563 | 1 | 0.067 | 0.739 | 1 | |
| IL-17F | 0.308 | 0.420 | 1 | −0.366 | 0.333 | 1 | 0.733 | 0.024 | 0.147 | |
| IL-4 | 0.384 | 0.218 | 1 | −0.357 | 0.255 | 1 | 0.291 | 0.358 | 1 | |
| IL-21 | 0.398 | 0.225 | 1 | −0.213 | 0.528 | 1 | 0.290 | 0.386 | 1 | |
| IL-22 | 0.182 | 0.552 | 1 | −0.360 | 0.309 | 1 | 0.475 | 0.101 | 0.606 | |
| CSF | IL-6 | 0.281 | 0.125 | 0.752 | 0.235 | 0.203 | 1 | 0.054 | 0.772 | 1 |
| IL-9 | 0.209 | 0.348 | 1 | 0.141 | 0.532 | 1 | −0.194 | 0.386 | 1 | |
| IL-10 | 0.044 | 0.877 | 1 | 0.221 | 0.429 | 1 | 0.159 | 0.569 | 1 | |
| IFN-γ | 0.016 | 0.940 | 1 | −0.054 | 0.800 | 1 | 0.197 | 0.356 | 1 | |
| IL-22 | 0.153 | 0.544 | 1 | −0.209 | 0.403 | 1 | −0.187 | 0.458 | 1 | |
| Cytokine | Blood Leukocyte Count | CSF Leukocyte Count | CSF Protein Level | CSF % Neutrophils | CSF % Lymphocytes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| rho | p | p adj. | rho | p | p adj. | rho | p | p adj. | rho | p | p adj. | rho | p | p adj. | |
| IL-6 | 0.143 | 0.503 | 1 | 0.552 | 0.005 | 0.077 | 0.322 | 0.125 | 1 | 0.423 | 0.039 | 0.585 | −0.437 | 0.033 | 0.490 |
| IL-9 | 0.263 | 0.307 | 1 | 0.178 | 0.495 | 1 | 0.185 | 0.477 | 1 | 0.321 | 0.209 | 1 | −0.327 | 0.199 | 1 |
| IL-10 | 0.218 | 0.519 | 1 | 0.654 | 0.029 | 0.433 | 0.245 | 0.467 | 1 | 0.409 | 0.211 | 1 | −0.427 | 0.189 | 1 |
| IFN-γ | 0.664 | 0.001 | 0.028 | 0.706 | 0.001 | 0.011 | 0.101 | 0.681 | 1 | 0.041 | 0.868 | 1 | −0.340 | 0.889 | 1 |
| IL-22 | −0.218 | 0.455 | 1 | −0.226 | 0.436 | 1 | −0.072 | 0.805 | 1 | −0.271 | 0.349 | 1 | 0.283 | 0.326 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zidovec-Lepej, S.; Vilibic-Cavlek, T.; Barbic, L.; Ilic, M.; Savic, V.; Tabain, I.; Ferenc, T.; Grgic, I.; Gorenec, L.; Bogdanic, M.; et al. Antiviral Cytokine Response in Neuroinvasive and Non-Neuroinvasive West Nile Virus Infection. Viruses 2021, 13, 342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020342
Zidovec-Lepej S, Vilibic-Cavlek T, Barbic L, Ilic M, Savic V, Tabain I, Ferenc T, Grgic I, Gorenec L, Bogdanic M, et al. Antiviral Cytokine Response in Neuroinvasive and Non-Neuroinvasive West Nile Virus Infection. Viruses. 2021; 13(2):342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020342
Chicago/Turabian StyleZidovec-Lepej, Snjezana, Tatjana Vilibic-Cavlek, Ljubo Barbic, Maja Ilic, Vladimir Savic, Irena Tabain, Thomas Ferenc, Ivana Grgic, Lana Gorenec, Maja Bogdanic, and et al. 2021. "Antiviral Cytokine Response in Neuroinvasive and Non-Neuroinvasive West Nile Virus Infection" Viruses 13, no. 2: 342. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020342