
Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Denv2 Strain
2.2. Virus Infection
2.3. cDNA Library Construction and Transcriptome Sequencing
2.4. Sequencing Analyses
2.5. qRT-PCR Validation of RNA-Seq Data
3. Results
3.1. Infection of C6/36 Cells with DENV2
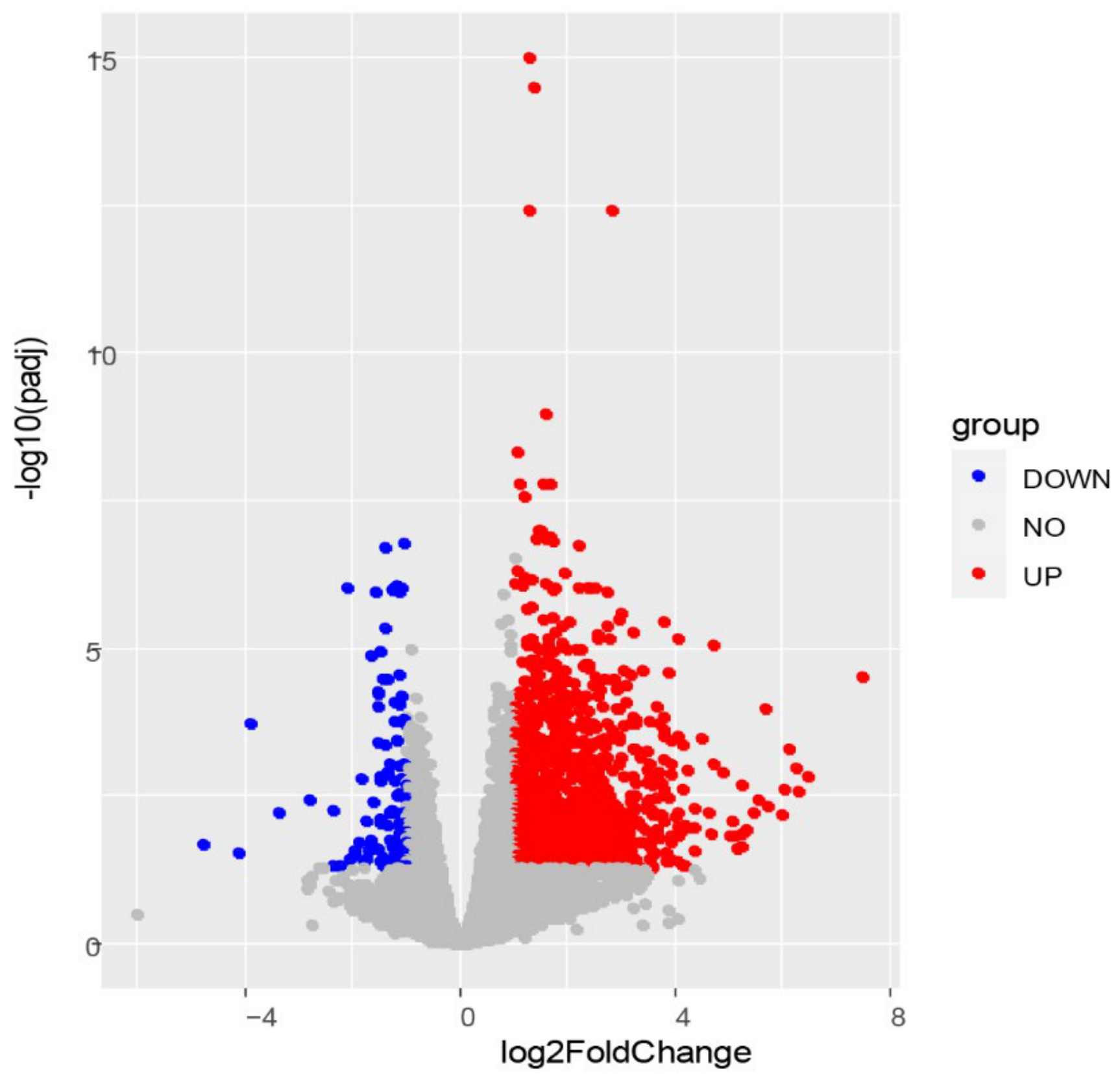
3.2. RNA-Seq Data and DEG Analysis
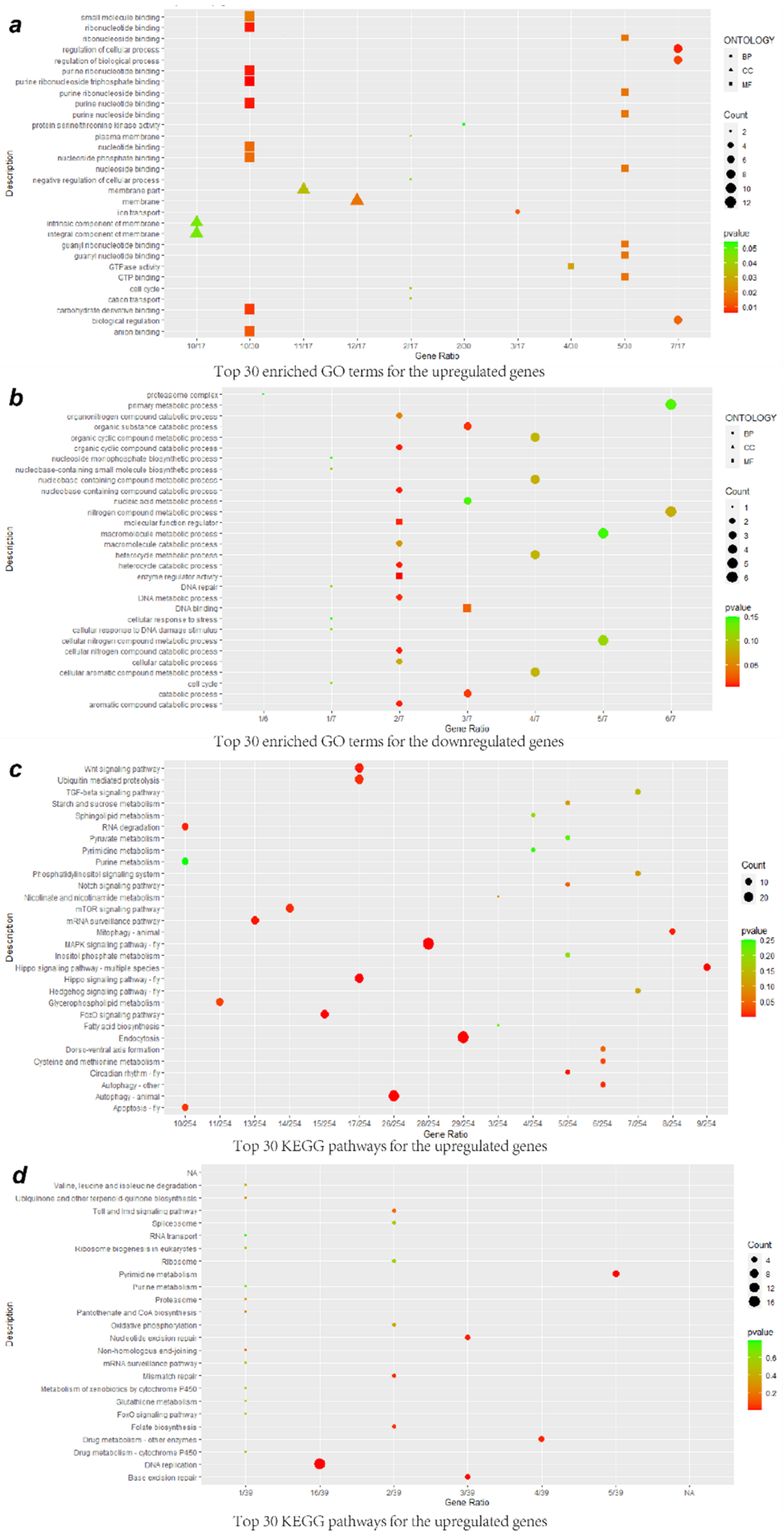
3.3. GO Enrichment Analysis of DEGs
3.4. KEGG Enrichment Analysis of DEGs
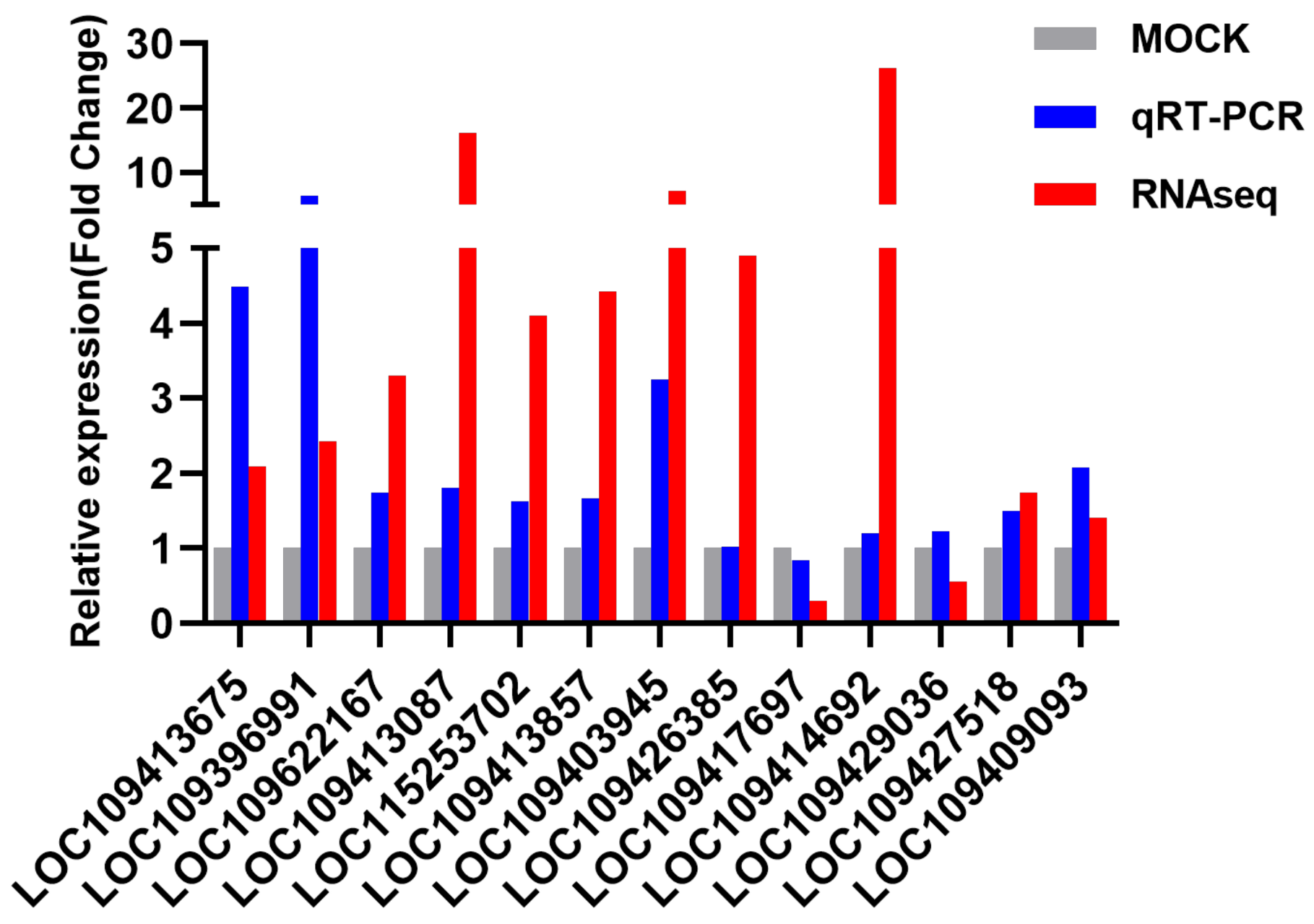
3.5. RNA-Seq Data Validation by qRT-PCR
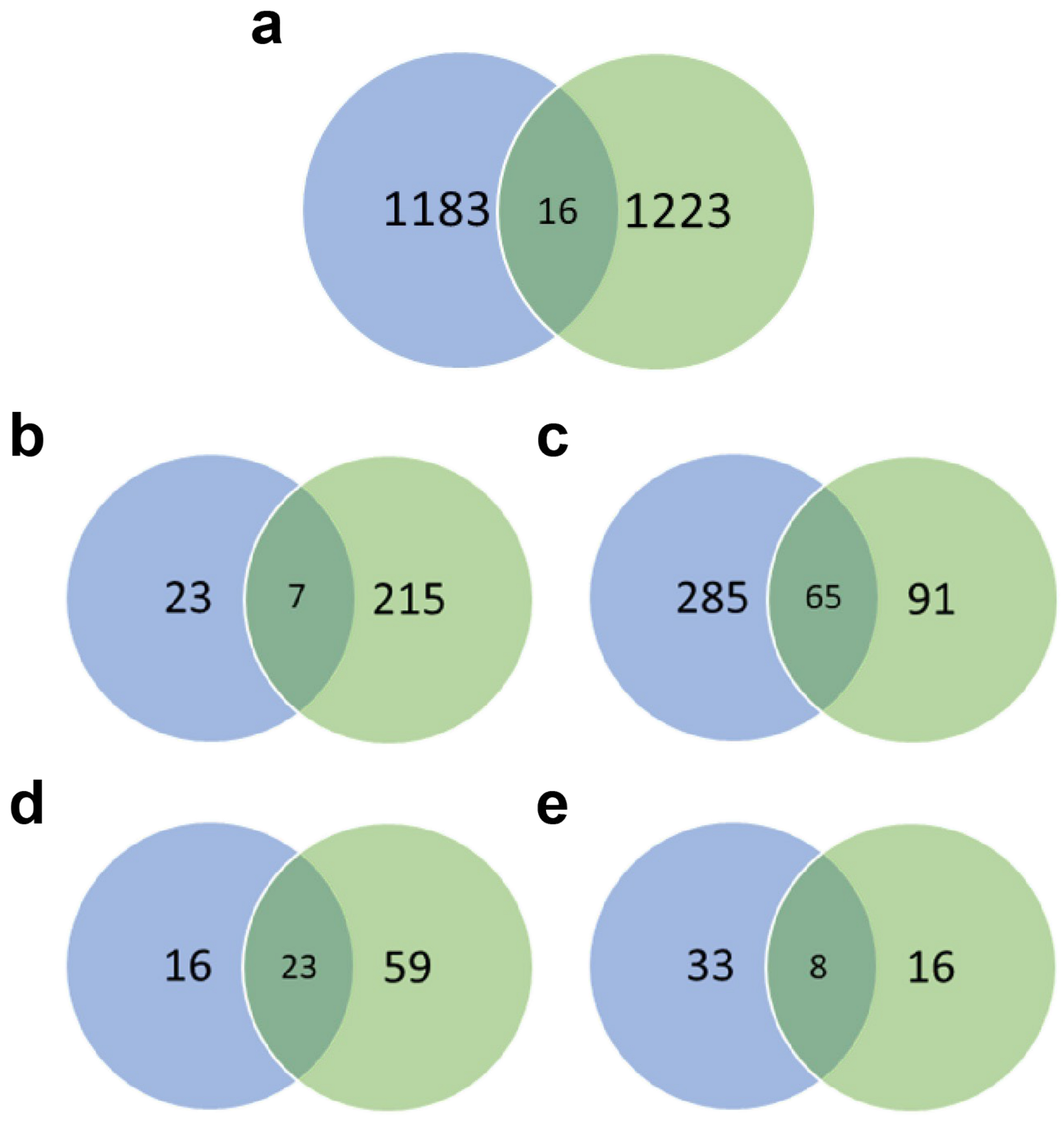
3.6. Comparison between C6/36 Cell and Aag2 Cell Transcriptomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castrom, C.; Wilsonm, E.; Bloomd, E. Disease and economic burdens of dengue. Lancet Infect. Dis. 2017, 17, e70–e78. [Google Scholar] [CrossRef]
- Messinaj, P.; Bradyo, J.; Scottt, W.; Zou, C.; Pigott, D.M.; Duda, K.A.; Bhatt, S.; Katzelnick, L.; Howes, R.E.; Battle, K.E.; et al. Global spread of dengue virus types: Mapping the 70 year history. Trends Microbiol. 2014, 22, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Dimopoulos, G. Insect immunity and its implication in mosquito-malaria interactions. Cell. Microbiol. 2003, 5, 3–14. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [Green Version]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Sanchez-Vargas, I.; Adelman, Z.N.; Blair, C.D.; Beaty, B.J.; James, A.A.; Olson, K.E. Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc. Natl. Acad. Sci. USA 2006, 103, 4198–4203. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.E.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito’s RNA interference pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Palomares, L.A.; Moreno-García, M.; Lanz-Mendoza, H.; Salazar, M.I. Molecular basis for arbovirus transmission by aedes aegypti mosquitoes. Intervirology 2018, 61, 255–264. [Google Scholar] [CrossRef]
- Walker, T.; Jeffries, C.L.; Mansfield, K.L.; Johnson, N. Mosquito cell lines: History, isolation, availability and application to assess the threat of arboviral transmission in the United Kingdom. Parasites Vectors 2014, 7, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.R.P. Cell cultures derived from larvae of Aedes albopictus (Skuse) and Aedes Aegypti (L). Curr. Sci. 1967, 36, 506–508. [Google Scholar]
- Miller, J.R.; Koren, S.; Dilley, K.A.; Puri, V.; Brown, D.M.; Harkins, D.M.; Thibaud-Nissen, F.; Rosen, B.; Chen, X.-G.; Tu, Z.; et al. Analysis of the Aedes albopictus C6/36 genome provides insight into cell line utility for viral propagation. GigaScience 2018, 7, gix135. [Google Scholar] [CrossRef]
- Brackney, D.E.; Scott, J.C.; Sagawa, F.; Woodward, J.E.; Miller, N.A.; Schilkey, F.D.; Mudge, J.; Wiolusz, J.; Olson, K.E.; Blair, C.; et al. C6/36 Aedes albopictus cells have a dysfunctional antiviral RNA interference response. PLoS Negl. Trop. Dis. 2010, 4, e856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi, A. Isolation of a Singh’s Aedes albopictus cell clone sensitive to Dengue and Chikungunya viruses. J. Gen. Virol. 1978, 40, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Martínez, A.B.; Vega-Almeida, T.O.; Salas-Benito, M.; García-Espitia, M.; De Nova-Ocampo, M.; Del Ángel, R.M.; Salas-Benito, J.S. Detection and sequencing of defective viral genomes in C6/36 cells persistently infected with dengue virus 2. Arch. Virol. 2013, 158, 583–599. [Google Scholar] [CrossRef]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Infectious dengue-1 virus entry into mosquito C6/36 cells. Virus Res. 2011, 160, 173–179. [Google Scholar] [CrossRef]
- Sultan, M.; Schulz, M.H.; Richard, H.; Magen, A.; Klingenhoff, A.; Scherf, M.; Seifert, M.; Borodina, T.; Soldatov, A.; Parkhomchuk, D.; et al. A global view of gene activity and alternative splicing by deep sequencing of the human transcriptome. Science 2008, 321, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Hrdlickova, R.; Toloue, M.; Tian, B. RNA-Seq methods for transcriptome analysis. Wiley Interdiscip. Rev. RNA 2017, 8, e1364. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-G.; Jiang, X.; Gu, J.; Xu, M.; Wu, Y.; Deng, Y.; Zhang, C.; Bonizzoni, M.; Dermauw, W.; Vontas, J.; et al. Genome sequence of the Asian Tiger mosquito, Aedes albopictus, reveals insights into its biology, genetics, and evolution. Proc. Natl. Acad. Sci. USA 2015, 112, E5907–E5915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarre, D.D.; Lowy, R.J. Improvements in methods for calculating virus titer estimates from TCID50 and plaque assays. J. Virol. Methods 2001, 96, 107–126. [Google Scholar] [CrossRef]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. ClusterProfiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Du, J.; Gao, S.; Tian, Z.; Guo, Y.; Kang, D.; Xing, S.; Zhang, G.; Liu, G.; Luo, J.; Chang, H.; et al. Transcriptome analysis of responses to bluetongue virus infection in Aedes albopictus cells. BMC Microbiol. 2019, 19, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bubner, B.; Gase, K.; Baldwin, I.T. Two-Fold differences are the detection limit for determining transgene copy numbers in plants by real-time PCR. BMC Biotechnol. 2004, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Li, M.-j.; Lan, C.-j.; Gao, H.-t.; Xing, D.; Gu, Z.-y.; Su, D.; Zhao, T.-y.; Yang, H.-y.; Li, C.-x. Transcriptome analysis of Aedes aegypti Aag2 cells in response to dengue virus-2 infection. Parasites Vectors 2020, 13, 421. [Google Scholar] [CrossRef]
- Khetarpal, N.; Khanna, I. Dengue fever: Causes, complications, and vaccine strategies. J. Immunol. Res. 2016, 2016, 6803098. [Google Scholar] [CrossRef] [Green Version]
- Green, A.M.; Beatty, P.R.; Hadjilaou, A.; Harris, E. Innate immunity to dengue virus infection and subversion of antiviral responses. J. Mol. Biol. 2014, 426, 1148–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedururu, R.K.; Neave, M.J.; Tachedjian, M.; Klein, M.J.; Gorry, P.R.; Duchemin, J.-B.; Paradkar, P.N. RNASeq analysis of Aedes albopictus mosquito midguts after chikungunya virus infection. Viruses 2019, 11, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedururu, R.K.; Neave, M.J.; Sundaramoorthy, V.; Green, D.; Harper, J.A.; Gorry, P.R.; Duchemin, J.-B.; Paradkar, P.N. Whole transcriptome analysis of Aedes albopictus mosquito head and thorax post-chikungunya virus infection. Pathogens 2019, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Shrinet, J.; Srivastava, P.; Sunil, S. Transcriptome analysis of Aedes aegypti in response to mono-infections and co-infections of dengue virus-2 and chikungunya virus. Biochem. Biophys. Res. Commun. 2017, 492, 617–623. [Google Scholar] [CrossRef]
- Kang, D.S.; Barron, M.S.; Lovin, D.D.; Cunningham, J.M.; Eng, M.W.; Chadee, D.D.; Li, J.; Severson, D.W. A transcriptomic survey of the impact of environmental stress on response to dengue virus in the mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 2018, 12, e0006568. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Dunn, W.A.; Campbell, C.L.; Olson, K.E.; Marinotti, O.; James, A.A. Complex modulation of the Aedes aegypti transcriptome in response to dengue virus infection. PLoS ONE 2012, 7, e50512. [Google Scholar] [CrossRef]
- Garavito, M.F.; Narváez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine metabolism: Dynamic and versatile pathways in pathogens and cellular development. J. Genet. Genom. 2015, 42, 195–205. [Google Scholar] [CrossRef]
- Vastag, L.; Koyuncu, E.; Grady, S.L.; Shenk, T.E.; Rabinowitz, J.D. Divergent effects of human cytomegalovirus and herpes simplex virus-1 on cellular metabolism. PLoS Pathog. 2011, 7, e1002124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, K.L.; Morrisey, J.M.; Wilinski, D.; Burns, J.M.; Vaidya, A.B.; Rabinowitz, J.D.; Llinás, M. Host-Parasite interactions revealed by Plasmodium falciparum metabolomics. Cell Host Microbe 2009, 5, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Chotiwan, N.; Andre, B.G.; Sanchez-Vargas, I.; Islam, M.N.; Grabowski, J.M.; Hopf-Jannasch, A.; Gough, E.; Nakayasu, E.; Blair, C.D.; Belisle, J.T.; et al. Dynamic remodeling of lipids coincides with dengue virus replication in the midgut of Aedes aegypti mosquitoes. PLoS Pathog. 2018, 14, e1006853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoeman, J.C.; Hou, J.; Harms, A.C.; Vreeken, R.J.; Berger, R.; Hankemeier, T.; Boonstra, A. Metabolic characterization of the natural progression of chronic hepatitis B. Genome Med. 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, J.W.; Chan, F.K. Staying alive: Cell death in antiviral immunity. Mol. Cell 2014, 54, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Marsh, M.; Helenius, A. Virus entry: Open sesame. Cell 2006, 124, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Pelkmans, L.; Helenius, A. Insider information: What viruses tell us about endocytosis. Curr. Opin. Cell Biol. 2003, 15, 414–422. [Google Scholar] [CrossRef]
- Sieczkarski, S.B.; Whittaker, G.R. Dissecting virus entry via endocytosis. J. Gen. Virol. 2002, 83, 1535–1545. [Google Scholar] [CrossRef]
- Acosta, E.G.; Castilla, V.; Damonte, E.B. Functional entry of dengue virus into Aedes albopictus mosquito cells is dependent on clathrin-mediated endocytosis. J. Gen. Virol. 2008, 89, 474–484. [Google Scholar] [CrossRef]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Richards, A.L.; Jackson, W.T. How positive-strand RNA viruses benefit from autophagosome maturation. J. Virol. 2013, 87, 9966–9972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.-R.; Lei, H.-Y.; Liu, M.-T.; Wang, J.-R.; Chen, S.-H.; Jiang-Shieh, Y.-F.; Lin, Y.-S.; Yeh, T.-M.; Liu, C.-C.; Liu, H.-S. Autophagic machinery activated by dengue virus enhances virus replication. Virology 2008, 374, 240–248. [Google Scholar] [CrossRef] [Green Version]
- Mateo, R.; Nagamine, C.M.; Spagnolo, J.; Méndez, E.; Rahe, M.; Gale, M.; Yuan, J.; Kirkegaard, K. Inhibition of cellular autophagy deranges dengue virion maturation. J. Virol. 2013, 87, 1312–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamar, N.H.; Kritsiligkou, P.; Grant, C.M. Loss of mRNA surveillance pathways results in widespread protein aggregation. Sci. Rep. 2018, 8, 3894. [Google Scholar] [CrossRef] [Green Version]
- Wada, M.; Lokugamage, K.G.; Nakagawa, K.; Narayanan, K.; Makino, S. Interplay between coronavirus, a cytoplasmic RNA virus, and nonsense-mediated mRNA decay pathway. Proc. Natl. Acad. Sci. USA 2018, 115, E10157–E10166. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, K.A.; Leon, K.E.; Khalid, M.M.; Tomar, S.; Jimenez-Morales, D.; Dunlap, M.; Kaye, J.A.; Shah, P.S.; Finkbeiner, S.; Krogan, N.J.; et al. The cellular NMD pathway restricts zika virus infection and is targeted by the viral capsid protein. mBio 2018, 9, e02126-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciechanover, A.; Schwartz, A.L. The ubiquitin-mediated proteolytic pathway: Mechanisms of recognition of the proteolytic substrate and involvement in the degradation of native cellular proteins. FASEB J. 1994, 8, 182–191. [Google Scholar] [CrossRef]
- Vu, P.K.; Sakamoto, K.M. Ubiquitin-Mediated proteolysis and human disease. Mol. Genet. Metab. 2000, 71, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, M.D.; Fradet-Turcotte, A. Virus DNA replication and the host DNA damage response. Annu. Rev. Virol. 2018, 5, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.T.; Park, E.-M.; Lim, Y.-S.; Hwang, S.B. Nonstructural protein 5A impairs DNA damage repair: Implications for hepatitis C virus-mediated hepatocarcinogenesis. J. Virol. 2018, 92, e00178-18. [Google Scholar] [CrossRef] [Green Version]
- Chambers, B.S.; Heaton, B.E.; Rausch, K.; Dumm, R.E.; Hamilton, J.R.; Cherry, S.; Heaton, N.S. DNA mismatch repair is required for the host innate response and controls cellular fate after influenza virus infection. Nat. Microbiol. 2019, 4, 1964–1977. [Google Scholar] [CrossRef] [PubMed]
- Haas, I.G.; Wabl, M. Immunoglobulin heavy chain binding protein. Nature 1983, 306, 387–389. [Google Scholar] [CrossRef]
- Pobre, K.F.R.; Poet, G.J.; Hendershot, L.M. The endoplasmic reticulum (ER) chaperone BiP is a master regulator of ER functions: Getting by with a little help from ERdj friends. J. Biol. Chem. 2019, 294, 2098–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panayi, G.S.; Corrigall, V.M. Immunoglobulin heavy-chain-binding protein (BiP): A stress protein that has the potential to be a novel therapy for rheumatoid arthritis. Biochem. Soc. Trans. 2014, 42, 1752–1755. [Google Scholar] [CrossRef]
- Lewy, T.G.; Grabowski, J.M.; Bloom, M.E. BiP: Master regulator of the unfolded protein response and crucial factor in flavivirus biology. Yale J. Biol. Med. 2017, 90, 291–300. [Google Scholar] [PubMed]
- Turpin, J.; Frumence, E.; Harrabi, W.; Haddad, J.G.; El Kalamouni, C.; Desprès, P.; Krejbich-Trotot, P.; Viranaïcken, W. Zika virus subversion of chaperone GRP78/BiP expression in A549 cells during UPR activation. Biochimie 2020, 175, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.-H.; Chiang, Y.-H.; Hou, J.-N.; Cheng, C.-C.; Sofiyatun, E.; Chiu, C.-H.; Chen, W.-J. XBP1-Mediated BiP/GRP78 upregulation copes with oxidative stress in mosquito cells during dengue 2 virus infection. BioMed Res. Int. 2017, 2017, 3519158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.Y.; Kaufman, R.J. The unfolded protein response. J. Cell Sci. 2003, 116, 1861–1862. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Zhong, Y.; Sanborn, M.A.; Teoh, T.C.; Ruan, J.; Yusof, R.; Hang, J.; Henderson, M.J.; Fang, S. Small molecule grp94 inhibitors block dengue and Zika virus replication. Antivir. Res. 2019, 171, 104590. [Google Scholar] [CrossRef]
- Zhu, H.; Blake, S.; Chan, K.T.; Pearson, R.B.; Kang, J. Cystathionine β-Synthase in physiology and cancer. BioMed Res. Int. 2018, 2018, 3205125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikawada, T.; Saito, A.; Kanamori, Y.; Nakahara, Y.; Iwata, K.-I.; Tanaka, D.; Watanabe, M.; Okuda, T. Trehalose transporter 1, a facilitated and high-capacity trehalose transporter, allows exogenous trehalose uptake into cells. Proc. Natl. Acad. Sci. USA 2007, 104, 11585–11590. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Description | Primer | Length (bp) |
|---|---|---|---|
| LOC109405344 | β-actin | F:GGAGAAGATCTGGCATCACA | 95 |
| R:TGTCATCTTCTCGCGGTTAG | |||
| LOC109413675 | endoplasmic reticulum chaperone BiP | F:TCGAGAGCTACGCCTACAGT | 97 |
| R:GCTTCCTCCATCTTGGCCTT | |||
| LOC109396991 | endoplasmin-like | F:GTACTCCATCTCCGCTCTGC | 87 |
| R:TCCTCGGACTCGTTCAGGAT | |||
| LOC109622167 | protein lifeguard 1 | F:GTGGAGAGATGCGACGGAAA | 129 |
| R:TACCAACCGCCTTGAGAACC | |||
| LOC109413087 | RNA polymerase II transcriptional coactivator | F:GACTCCGCTAGCACAACCAA | 140 |
| R:GGCAGGGATTGACCGTCTTT | |||
| LOC115253702 | mitochondrial carrier protein Rim2-like | F:AGTAACATGCCCGCTGGAAG | 92 |
| R:TCGATTGGGCAATACGAGGC | |||
| LOC109413857 | cold shock domain-containing protein E1-like | F:ACGTTTTTGGTGGAGGTGACA | 105 |
| R:CTCCACCGGACATGCTCATC | |||
| LOC109403945 | endoribonuclease Dicer | F:GGTGGATCCGAAGACTAGCG | 114 |
| R:GTCTTGTCCCCCATGGTCTG | |||
| LOC109426385 | insulin-like growth factor 2 mRNA-binding protein 1 | F:GTTCGAGAGCCACGAACAAG | 108 |
| R:CGGTTCGCTTTGTTACTGTCG | |||
| LOC109417697 | glutathione S-transferase 1-like | F:CTTCTCGAGCCGTGGATCTC | 127 |
| R:GGGATTGTATGCCGAGGGTT | |||
| LOC109414692 | protein kibra | F:TCACATCGTCAGCCGATTCC | 107 |
| R:TCGTAAACAGCGACTGCGAT | |||
| LOC109429036 | UDP-glucuronosyl-transferase 2B1-like | F:ACGAGTCGGTCCAAGGTCTA | 129 |
| R:CTCCCTCCTGAGTTCCCAGA | |||
| LOC109427518 | cystathionine beta-synthase | F:GCCACGTCCCAAGGTTATGA | 101 |
| R:CATTGCACTTCAACCCTGCC | |||
| LOC109409093 | facilitated trehalose transporter Tret1 | F:TCTTCGGGGGATGCTTTGTG | 124 |
| R:CACCGGAAGGCATGCAGATA |
| Gene | RNA-Seq | 1 dpi | 2 dpi | 4 dpi | ||||
|---|---|---|---|---|---|---|---|---|
| FC | Padj | FC | p | FC | p | FC | p | |
| LOC109413675 | 2.08 | 0.000 | 1.19 | 0.275 | 1.02 | 0.711 | 4.49 | 0.000 |
| LOC109396991 | 2.43 | 0.000 | 1.89 | 0.002 | 1.31 | 0.001 | 6.41 | 0.000 |
| LOC109622167 | 3.30 | 0.000 | 1.04 | 0.801 | 0.96 | 0.667 | 1.74 | 0.002 |
| LOC109413087 | 16.12 | 0.007 | 1.06 | 0.788 | 1.07 | 0.514 | 1.81 | 0.008 |
| LOC115253702 | 4.10 | 0.009 | 0.69 | 0.360 | 0.72 | 0.057 | 1.62 | 0.097 |
| LOC109413857 | 4.43 | 0.005 | 0.79 | 0.568 | 1.09 | 0.549 | 1.66 | 0.036 |
| LOC109403945 | 7.04 | 0.000 | 0.86 | 0.701 | 0.96 | 0.648 | 3.26 | 0.004 |
| LOC109426385 | 4.90 | 0.030 | 0.75 | 0.077 | 0.55 | 0.042 | 1.02 | 0.939 |
| LOC109417697 | 0.29 | 0.041 | 0.84 | 0.554 | 0.74 | 0.035 | 0.84 | 0.313 |
| LOC109414692 | 26.23 | 0.001 | 0.03 | 0.239 | 0.85 | 0.677 | 1.20 | 0.729 |
| LOC109429036 | 0.55 | 0.015 | 2.65 | 0.008 | 1.85 | 0.003 | 1.23 | 0.487 |
| LOC109427518 | 1.74 | 0.028 | 0.83 | 0.634 | 0.62 | 0.019 | 1.50 | 0.010 |
| LOC109409093 | 1.40 | 0.050 | 0.95 | 0.887 | 1.07 | 0.638 | 2.07 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Xing, D.; Su, D.; Wang, D.; Gao, H.; Lan, C.; Gu, Z.; Zhao, T.; Li, C. Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells. Viruses 2021, 13, 343. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020343
Li M, Xing D, Su D, Wang D, Gao H, Lan C, Gu Z, Zhao T, Li C. Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells. Viruses. 2021; 13(2):343. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020343
Chicago/Turabian StyleLi, Manjin, Dan Xing, Duo Su, Di Wang, Heting Gao, Cejie Lan, Zhenyu Gu, Tongyan Zhao, and Chunxiao Li. 2021. "Transcriptome Analysis of Responses to Dengue Virus 2 Infection in Aedes albopictus (Skuse) C6/36 Cells" Viruses 13, no. 2: 343. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020343