
Predominance of HBV Genotype B and HDV Genotype 1 in Vietnamese Patients with Chronic Hepatitis

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
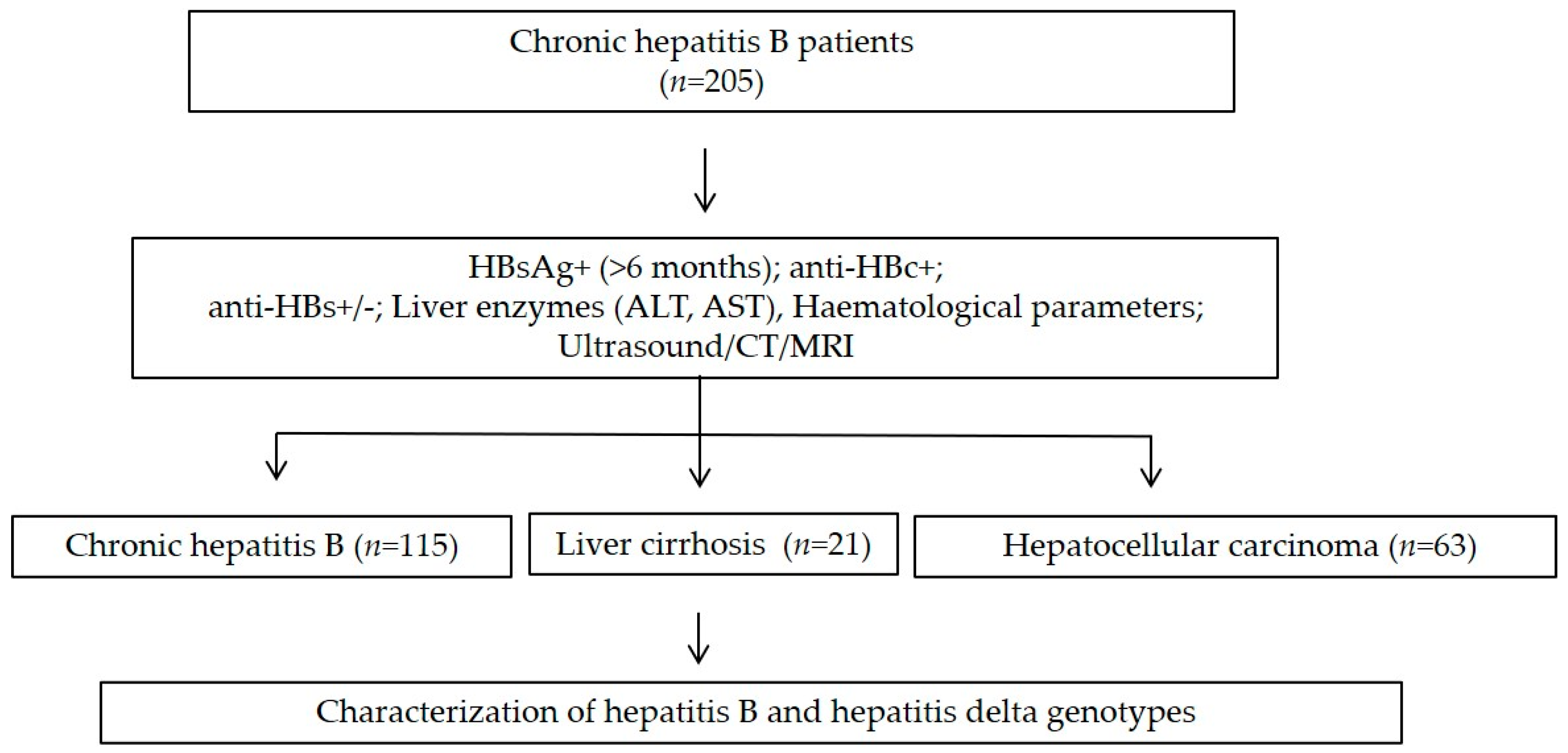
2.2. Study Cohort
2.3. Serological Assays
2.4. Nucleic Acids Isolation
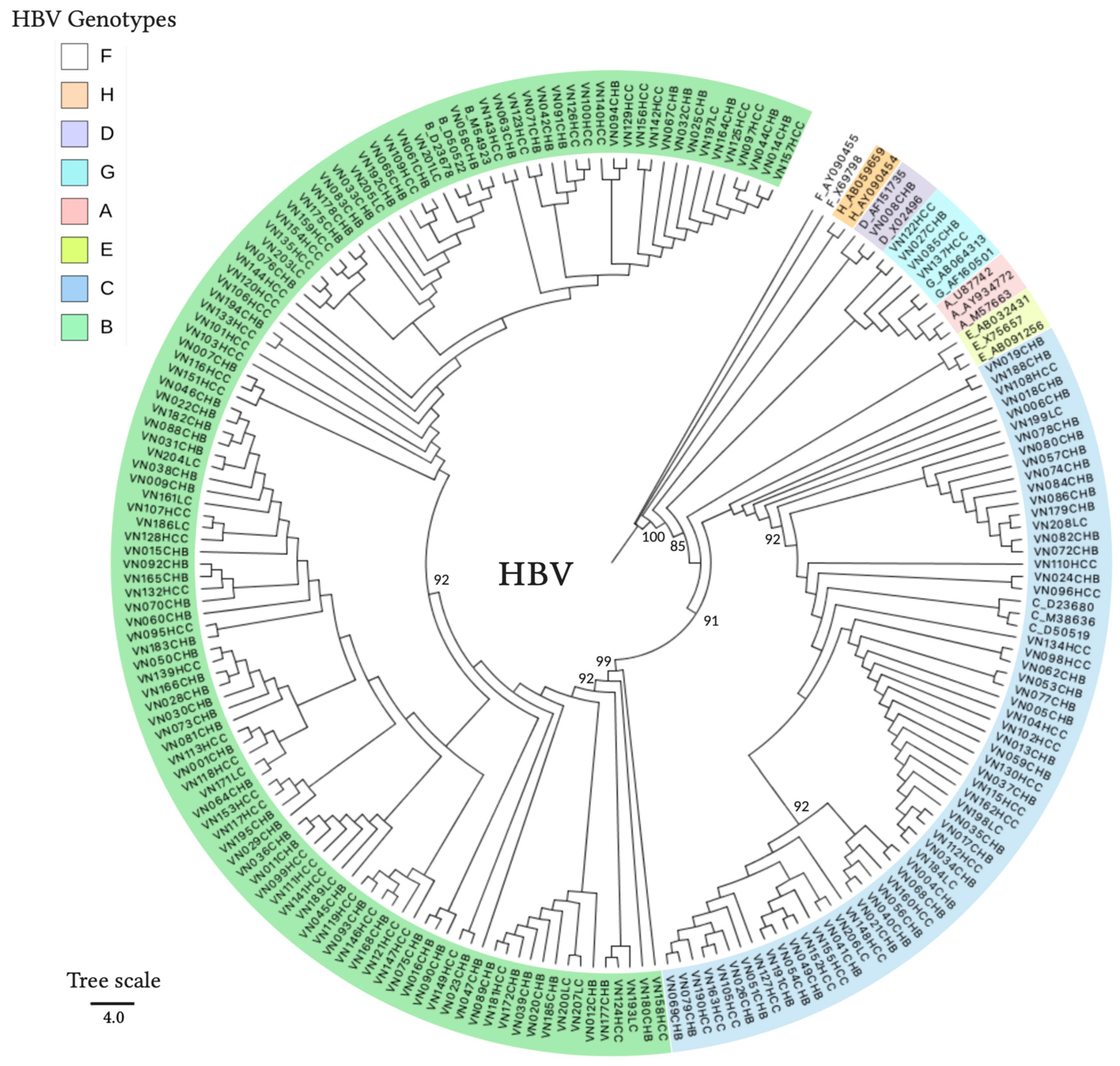
2.5. HBV and HDV Genotyping
2.6. Sequence Data Analysis
2.7. Statistical Analysis
3. Results
3.1. Baseline Characteristics of HBV Patients
3.2. HBV and HDV Genotypes and Their Associations with the Outcomes of the Patients
3.3. Hepatitis B Surface Antigen Escape Mutations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Hepatitis B Fact Sheet. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 1 January 2021).
- WHO. Hepatitis D Fact Sheet. Available online: https://www.who.int/en/news-room/fact-sheets/detail/hepatitis-d (accessed on 1 January 2021).
- Shen, T.; Yan, X.M. Hepatitis B virus genetic mutations and evolution in liver diseases. World J. Gastroenterol. 2014, 20, 5435–5441. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.S.; Choo, Q.L.; Weiner, A.J.; Ou, J.H.; Najarian, R.C.; Thayer, R.M.; Mullenbach, G.T.; Denniston, K.J.; Gerin, J.L.; Houghton, M. Structure, sequence and expression of the hepatitis delta (delta) viral genome. Nature 1986, 323, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Dandri, M. Hepatitis B and Delta Virus: Advances on Studies about Interactions between the Two Viruses and the Infected Hepatocyte. J. Clin. Transl. Hepatol. 2015, 3, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Rizzetto, M.; Hoyer, B.; Canese, M.G.; Shih, J.W.; Purcell, R.H.; Gerin, J.L. delta Agent: Association of delta antigen with hepatitis B surface antigen and RNA in serum of delta-infected chimpanzees. Proc. Natl. Acad. Sci. USA 1980, 77, 6124–6128. [Google Scholar] [CrossRef] [Green Version]
- Ringelhan, M.; McKeating, J.A.; Protzer, U. Viral hepatitis and liver cancer. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [Green Version]
- Fattovich, G.; Bortolotti, F.; Donato, F. Natural history of chronic hepatitis B: Special emphasis on disease progression and prognostic factors. J. Hepatol. 2008, 48, 335–352. [Google Scholar] [CrossRef]
- Fattovich, G.; Giustina, G.; Christensen, E.; Pantalena, M.; Zagni, I.; Realdi, G.; Schalm, S.W. Influence of hepatitis delta virus infection on morbidity and mortality in compensated cirrhosis type B. The European Concerted Action on Viral Hepatitis (Eurohep). Gut 2000, 46, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Tatematsu, K.; Tanaka, Y.; Kurbanov, F.; Sugauchi, F.; Mano, S.; Maeshiro, T.; Nakayoshi, T.; Wakuta, M.; Miyakawa, Y.; Mizokami, M. A genetic variant of hepatitis B virus divergent from known human and ape genotypes isolated from a Japanese patient and provisionally assigned to new genotype J. J. Virol. 2009, 83, 10538–10547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.; Trinh, T.N.; Abe, K. New complex recombinant genotype of hepatitis B virus identified in Vietnam. J. Virol. 2008, 82, 5657–5663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gal, F.; Gault, E.; Ripault, M.P.; Serpaggi, J.; Trinchet, J.C.; Gordien, E.; Deny, P. Eighth major clade for hepatitis delta virus. Emerg. Infect. Dis. 2006, 12, 1447–1450. [Google Scholar] [CrossRef]
- Le Gal, F.; Brichler, S.; Drugan, T.; Alloui, C.; Roulot, D.; Pawlotsky, J.M.; Deny, P.; Gordien, E. Genetic diversity and worldwide distribution of the deltavirus genus: A study of 2152 clinical strains. Hepatology 2017, 66, 1826–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Kao, J.H. Natural history of acute and chronic hepatitis B: The role of HBV genotypes and mutants. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.H.; Chen, P.J.; Lai, M.Y.; Chen, D.S. Hepatitis B genotypes correlate with clinical outcomes in patients with chronic hepatitis B. Gastroenterology 2000, 118, 554–559. [Google Scholar] [CrossRef]
- Kao, J.H. Hepatitis B viral genotypes: Clinical relevance and molecular characteristics. J. Gastroenterol. Hepatol. 2002, 17, 643–650. [Google Scholar] [CrossRef]
- Yan, H.; Yang, Y.; Zhang, L.; Tang, G.; Wang, Y.; Xue, G.; Zhou, W.; Sun, S. Characterization of the genotype and integration patterns of hepatitis B virus in early- and late-onset hepatocellular carcinoma. Hepatology 2015, 61, 1821–1831. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.I.; Yeh, S.H.; Chen, P.J.; Iloeje, U.H.; Jen, C.L.; Su, J.; Wang, L.Y.; Lu, S.N.; You, S.L.; Chen, D.S.; et al. Associations between hepatitis B virus genotype and mutants and the risk of hepatocellular carcinoma. J. Natl. Cancer Inst. 2008, 100, 1134–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sy, B.T.; Ratsch, B.A.; Toan, N.L.; Song, L.H.; Wollboldt, C.; Bryniok, A.; Nguyen, H.M.; Luong, H.V.; Velavan, T.P.; Wedemeyer, H.; et al. High prevalence and significance of hepatitis D virus infection among treatment-naive HBsAg-positive patients in Northern Vietnam. PLoS ONE 2013, 8, e78094. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, Y.; Tang, Y.; Yao, T.; Lv, M.; Tang, Z.; Zang, G.; Yu, Y.; Chen, X. Molecular epidemiology and clinical characteristics of hepatitis delta virus (HDV) infected patients with elevated transaminases in Shanghai, China. BMC Infect. Dis. 2020, 20, 565. [Google Scholar] [CrossRef]
- Tong, H.V.; Song, L.H.; Hoan, N.X.; Cuong, B.K.; Sy, B.T.; Son, H.A.; Quyet, D.; Binh, V.Q.; Kremsner, P.G.; Bock, C.T.; et al. Soluble MICB protein levels and platelet counts during hepatitis B virus infection and response to hepatocellular carcinoma treatment. BMC Infect. Dis. 2015, 15, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, W.; Li, F.; Hua, F.; Zhan, Y.; Li, Y.; Ji, L.; Zou, S.; Min, Z.; Song, D.; et al. Abnormal platelet kinetics are detected before the occurrence of thrombocytopaenia in HBV-related liver disease. Liver Int. 2014, 34, 535–543. [Google Scholar] [CrossRef]
- Mohamed, R.; Desmond, P.; Suh, D.J.; Amarapurkar, D.; Gane, E.; Guangbi, Y.; Hou, J.L.; Jafri, W.; Lai, C.L.; Lee, C.H.; et al. Practical difficulties in the management of hepatitis B in the Asia-Pacific region. J. Gastroenterol. Hepatol. 2004, 19, 958–969. [Google Scholar] [CrossRef]
- Pham, T.T.; Le, H.M.; Nguyen, D.T.; Maertens, K.; Leuridan, E.; Theeten, H.; Hendrickx, G.; Van Damme, P. Assessment of the timely administration of the hepatitis B and BCG birth dose and the primary infant vaccination schedule in 2015–2016 in the Mekong Delta, Viet Nam. Vaccine 2018, 36, 5760–5765. [Google Scholar] [CrossRef] [PubMed]
- WHO. Hepatitis B (HepB3) Immunization Coverage among 1-Year-Olds. Available online: https://www.who.int/data/gho/data/indicators/indicator-details/GHO/hepatitis-b-(hepb3)-immunization-coverage-among-1-year-olds-(-) (accessed on 1 January 2021).
- Nguyen, V.T.; Law, M.G.; Dore, G.J. An enormous hepatitis B virus-related liver disease burden projected in Vietnam by 2025. Liver Int. 2008, 28, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A. Hepatitis B virus S gene escape mutants. Asian J. Transfus. Sci. 2007, 1, 62–70. [Google Scholar] [CrossRef]
- Romano, L.; Paladini, S.; Galli, C.; Raimondo, G.; Pollicino, T.; Zanetti, A.R. Hepatitis B vaccination. Hum. Vaccin Immunother 2015, 11, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Bui, T.T.T.; Tran, T.T.; Nghiem, M.N.; Rahman, P.; Tran, T.T.T.; Dinh, M.N.H.; Le, M.H.; Nguyen, V.V.C.; Thwaites, G.; Rahman, M. Molecular characterization of hepatitis B virus in Vietnam. BMC Infect. Dis. 2017, 17, 601. [Google Scholar] [CrossRef] [Green Version]
- Do, S.H. Epidemiology of Hepatitis B and C Virus Infections and Liver Cancer in Vietnam. Euroasian J. Hepatogastroenterol. 2015, 5, 49–51. [Google Scholar] [CrossRef]
- Nguyen, V.T.; McLaws, M.L.; Dore, G.J. Highly endemic hepatitis B infection in rural Vietnam. J. Gastroenterol. Hepatol. 2007, 22, 2093–2100. [Google Scholar] [CrossRef]
- Tran, H.T.; Ushijima, H.; Quang, V.X.; Phuong, N.; Li, T.C.; Hayashi, S.; Xuan Lien, T.; Sata, T.; Abe, K. Prevalence of hepatitis virus types B through E and genotypic distribution of HBV and HCV in Ho Chi Minh City, Vietnam. Hepatol. Res. 2003, 26, 275–280. [Google Scholar] [CrossRef]
- Binh, M.T.; Hoan, N.X.; Van Tong, H.; Giang, D.P.; Sy, B.T.; Toan, N.L.; Song, L.H.; Bang, M.H.; Wedemeyer, H.; Meyer, C.G.; et al. HDV infection rates in northern Vietnam. Sci. Rep. 2018, 8, 8047. [Google Scholar] [CrossRef]
- Dunford, L.; Carr, M.J.; Dean, J.; Nguyen, L.T.; Thi, T.H.T.; Nguyen, B.T.; Connell, J.; Coughlan, S.; Nguyen, H.T.; Hall, W.W.; et al. A multicentre molecular analysis of hepatitis B and blood-borne virus coinfections in Viet Nam. PLoS ONE 2012, 7, e39027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.M.; Sy, B.T.; Trung, N.T.; Hoan, N.X.; Wedemeyer, H.; Velavan, T.P.; Bock, C.T. Prevalence and genotype distribution of hepatitis delta virus among chronic hepatitis B carriers in Central Vietnam. PLoS ONE 2017, 12, e0175304. [Google Scholar] [CrossRef] [Green Version]
- Baqlani, S.A.A.; Sy, B.T.; Ratsch, B.A.; Naamani, K.A.; Awaidy, S.A.; Busaidy, S.A.; Pauli, G.; Bock, C.T. Molecular epidemiology and genotyping of hepatitis B virus of HBsAg-positive patients in Oman. PLoS ONE 2014, 9, e97759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayer, J.; Jadeau, F.; Deleage, G.; Kay, A.; Zoulim, F.; Combet, C. HBVdb: A knowledge database for Hepatitis B Virus. Nucleic Acids Res. 2013, 41, D566–D570. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree, version 1.3.1 ed; Computer Program; 2009; Available online: http://tree.bio.ed.ac.uk/software/ (accessed on 1 January 2021).
- Lazarevic, I.; Banko, A.; Miljanovic, D.; Cupic, M. Immune-Escape Hepatitis B Virus Mutations Associated with Viral Reactivation upon Immunosuppression. Viruses 2019, 11, 778. [Google Scholar] [CrossRef] [Green Version]
- Mendy, M.; D’Mello, F.; Kanellos, T.; Oliver, S.; Whittle, H.; Howard, C.R. Envelope protein variability among HBV-Infected asymptomatic carriers and immunized children with breakthrough infections. J. Med. Virol. 2008, 80, 1537–1546. [Google Scholar] [CrossRef]
- Nguyen, V.T. Hepatitis B infection in Vietnam: Current issues and future challenges. Asia Pac. J. Public Health 2012, 24, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.Z.; Jin, F.F.; Liu, X.; Liu, N.; Wen, F.; Lou, J.L. Coexistence of low levels of HBsAg and high levels of anti-HBs may increase risk of hepatocellular carcinoma in chronic hepatitis B patients with high HBV load. Braz. J. Infect. Dis. 2019, 23, 343–351. [Google Scholar] [CrossRef]
- Lada, O.; Benhamou, Y.; Poynard, T.; Thibault, V. Coexistence of hepatitis B surface antigen (HBs Ag) and anti-HBs antibodies in chronic hepatitis B virus carriers: Influence of “a” determinant variants. J. Virol. 2006, 80, 2968–2975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Katano, Y.; Chuong, T.X.; Takeda, Y.; Ishigami, M.; Itoh, A.; Hirooka, Y.; Nakano, I.; Huy, T.V.; Minh, N.N.; et al. Prevalence of hepatitis B virus subgenotypes and basal core promoter, precore variants in patients with acute hepatitis B in central Vietnam. Intervirology 2009, 52, 22–28. [Google Scholar] [CrossRef]
- Alvarado-Mora, M.V.; Locarnini, S.; Rizzetto, M.; Pinho, J.R. An update on HDV: Virology, pathogenesis and treatment. Antivir. Ther. 2013, 18, 541–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.A.; Wedemeyer, H.; Harrison, P.M. Hepatitis delta virus. Lancet 2011, 378, 73–85. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics/Parameters | Total (n = 205) | CHB (n = 115) | LC (n = 21) | HCC (n = 69) | p-Value |
|---|---|---|---|---|---|
| Age (years) | 49 [36–61] | 40 [31–53] | 45 [39–59] | 61 [53–67] | <0.0001 * |
| Male/female | 158/47 | 78/37 | 17/4 | 63/6 | 0.0011 # |
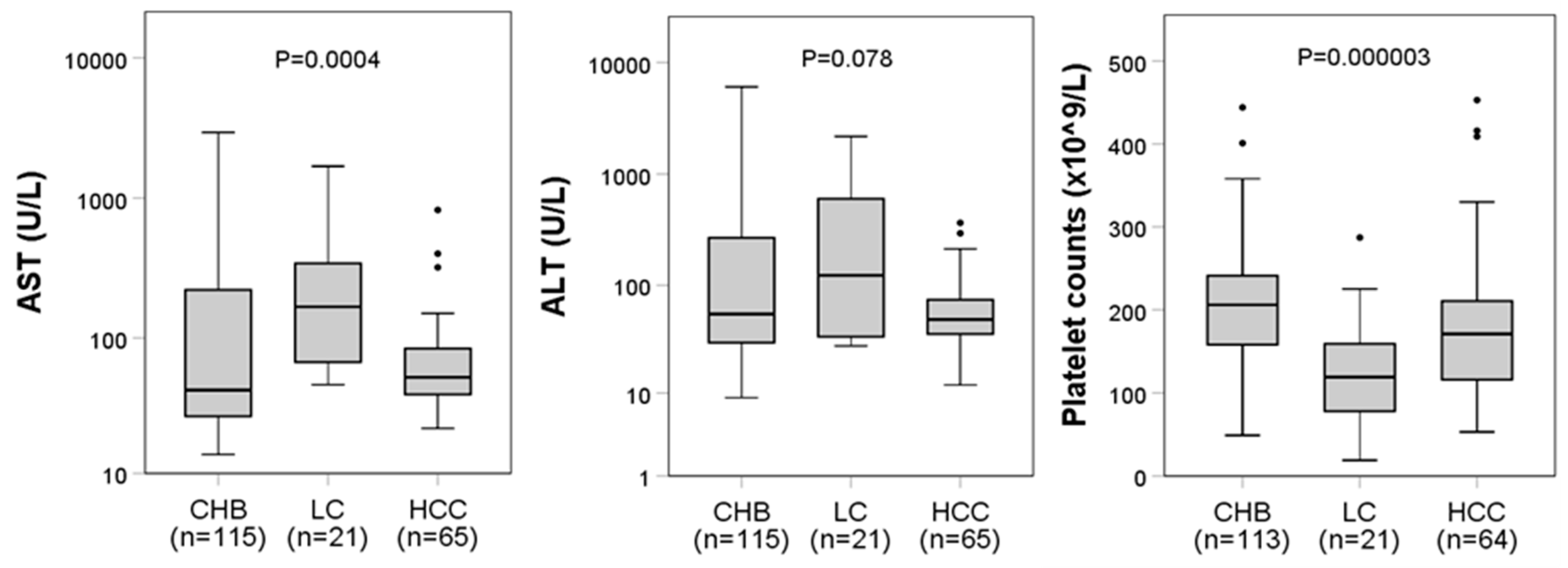
| AST (U/L) | 54 [32–157] | 42 [27–221] | 167 [67–342] | 52 [39–84] | 0.0004 * |
| ALT (U/L) | 55 [34–157] | 55 [30–269] | 123 [34–601] | 49 [36–74] | 0.0778 * |
| PLT (×109/L) | 187 [142–229] | 206 [158–241] | 119 [78–159] | 171 [118–210] | <0.0001 * |
| Anti-HBc (+/−) | 204/1 | 115/0 | 20/1 | 69/0 | N/A |
| Anti-HBs (+/−) | 15/190 | 5/110 | 1/20 | 9/60 | 0.0743 ~ |
| HBV genotypes (B/C/D/G) | 134/64/2/5 | 71/39/2/3 | 16/5/0/0 | 47/20/0/2 | N/A |
| HDV coinfections | 33 | 16 | 1 | 16 | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoan, N.X.; Hoechel, M.; Tomazatos, A.; Anh, C.X.; Pallerla, S.R.; Linh, L.T.K.; Binh, M.T.; Sy, B.T.; Toan, N.L.; Wedemeyer, H.; et al. Predominance of HBV Genotype B and HDV Genotype 1 in Vietnamese Patients with Chronic Hepatitis. Viruses 2021, 13, 346. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020346
Hoan NX, Hoechel M, Tomazatos A, Anh CX, Pallerla SR, Linh LTK, Binh MT, Sy BT, Toan NL, Wedemeyer H, et al. Predominance of HBV Genotype B and HDV Genotype 1 in Vietnamese Patients with Chronic Hepatitis. Viruses. 2021; 13(2):346. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020346
Chicago/Turabian StyleHoan, Nghiem Xuan, Mirjam Hoechel, Alexandru Tomazatos, Chu Xuan Anh, Srinivas Reddy Pallerla, Le Thi Kieu Linh, Mai Thanh Binh, Bui Tien Sy, Nguyen Linh Toan, Heiner Wedemeyer, and et al. 2021. "Predominance of HBV Genotype B and HDV Genotype 1 in Vietnamese Patients with Chronic Hepatitis" Viruses 13, no. 2: 346. https://0-doi-org.brum.beds.ac.uk/10.3390/v13020346