
In Vitro and In Vivo Characterization of a Pigeon Paramyxovirus Type 1 Isolated from Domestic Pigeons in Victoria, Australia 2011

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus Isolation and Identification
2.3. Hemagglutination (HA) and Hemagglutination Inhibition (HI) Tests
2.4. Elution Rate and Hemagglutinin Thermostability
2.5. Plaque Formation
2.6. Pathogenicity Tests
2.7. RT-PCR and Sequencing
3. Results
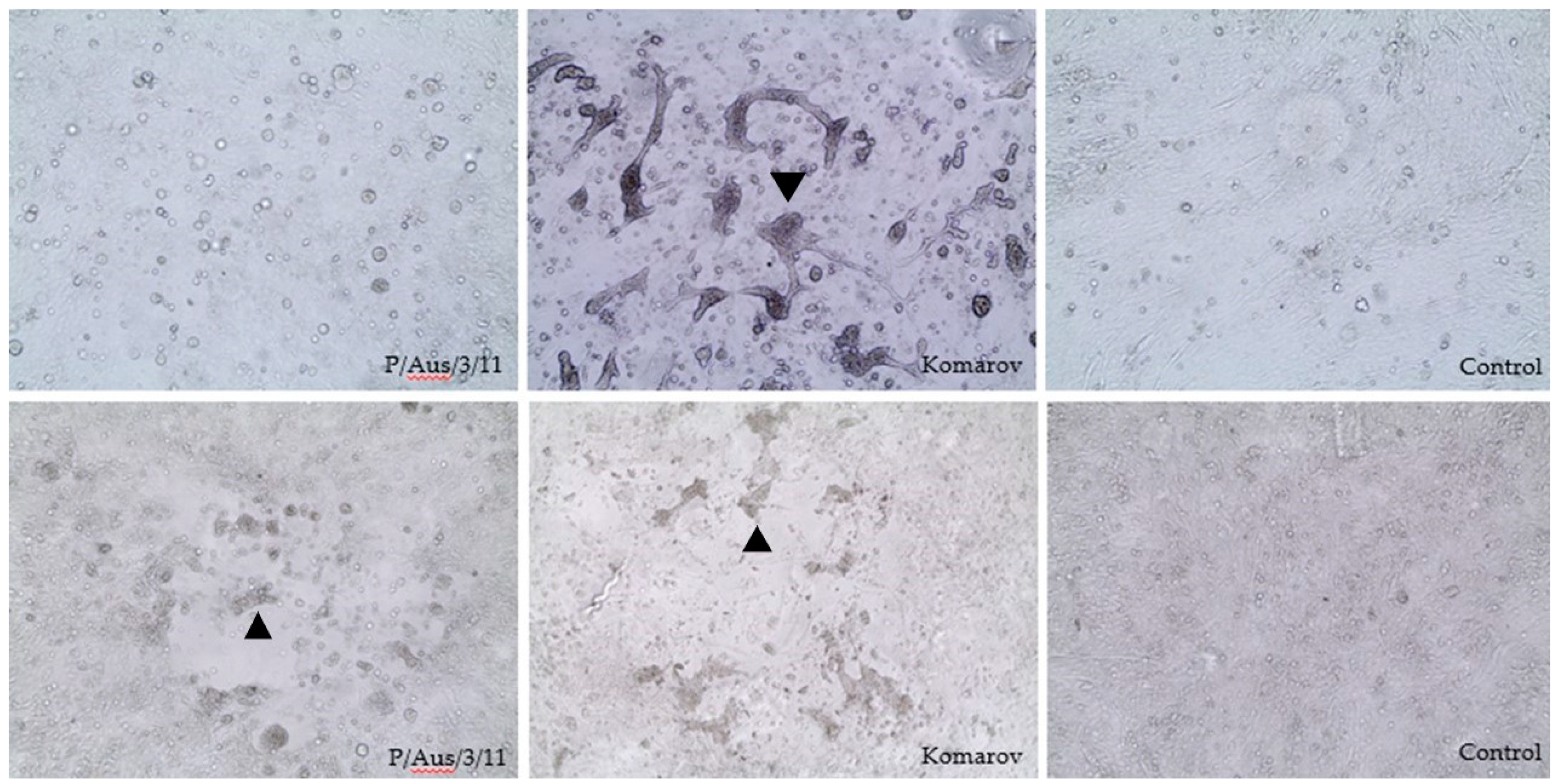
3.1. Virus Isolation and Identification
3.2. Virus Characterization
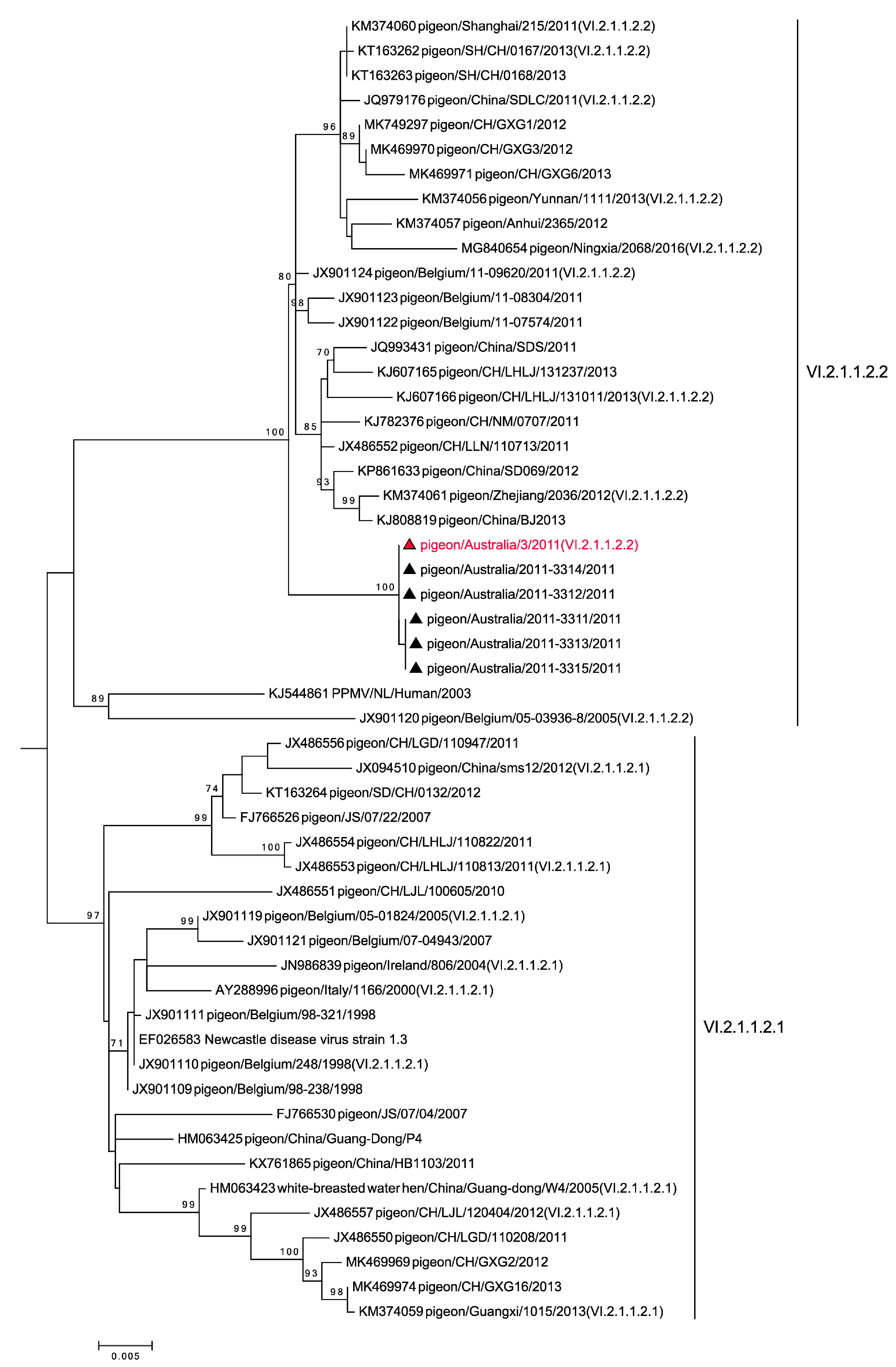
3.3. Genetic Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [Green Version]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, A.; Huang, Z.; Elankumaran, S.; Rockemann, D.D.; Samal, S.K. Role of fusion protein cleavage site in the virulence of Newcastle disease virus. Microb. Pathog. 2004, 36, 1–10. [Google Scholar] [CrossRef]
- Rout, S.N.; Samal, S.K. The Large Polymerase Protein Is Associated with the Virulence of Newcastle Disease Virus. J. Virol. 2008, 82, 7828–7836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Terrestrial Manual Chapter 2.3.14 Newcastle Disease (Infection with Newcastle Disease Virus); OIE: Paris, France, 2012; pp. 555–574. [Google Scholar]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Virulence of six heterogeneous-origin Newcastle disease virus isolates before and after sequential passages in domestic chickens. Avian Pathol. 2003, 32, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.; King, D.J.; Seal, B.S. Pathogenesis of Newcastle disease in chickens experimentally infected with viruses of different virulence. Vet. Pathol. 1999, 36, 125–132. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Aldous, E.W.; Mynn, J.K.; Banks, J.; Alexander, D.J. A molecular epidemiological study of avian paramyxovirus type 1 (Newcastle disease virus) isolates by phylogenetic analysis of a partial nucleotide sequence of the fusion protein gene. Avian Pathol. 2003, 32, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect. Genet. Evol. 2016, 39, 22–34. [Google Scholar] [CrossRef] [Green Version]
- King, D.J. Avian Paramyxovirus Type 1 from Pigeons: Isolate Characterization and Pathogenicity after Chicken or Embryo Passage of Selected Isolates. Avian Dis. 1996, 40, 707–714. [Google Scholar] [CrossRef]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Virulence of Pigeon-Origin Newcastle Disease Virus Isolates for Domestic Chickens. Avian Dis. 2001, 45, 906–921. [Google Scholar] [CrossRef] [PubMed]
- Wildlife Health Australia Avian Paramyxoviruses and Australian Wild Birds. Available online: https://wildlifehealthaustralia.com.au/FactSheets.aspx (accessed on 1 November 2019).
- Alexander, D.J.; Wilson, G.W.; Russell, P.H.; Lister, S.A.; Parsons, G. Newcastle disease outbreaks in fowl in Great Britain during 1984. Vet. Rec. 1985, 117, 429–434. [Google Scholar] [CrossRef]
- Alexander, D.J.; Parsons, G. Pathogenicity for chickens of avian paramyxovirus type I isolates obtained from pigeons in great britain during 1983–1985. Avian Pathol. 1986, 15, 487–493. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes; NHMRC: Canberra, Australia, 2013.
- Swayne, D.E.; Senne, D.A.; Beard, C.W. Avian Influenza. In A Laboratory Manual for the Isolation, Identification and Characterization of Avian Pathogens, 5th ed.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2008. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Biancifiori, F.; Fioroni, A. An occurrence of Newcastle disease in pigeons: Virological and serological studies on the isolates. Comp. Immunol. Microbiol. Infect. Dis. 1983, 6, 247–252. [Google Scholar] [CrossRef]
- Spalatin, J.; Hanson, R.P.; Beard, P.D. The hemagglutination-elution pattern as a marker in characterizing Newcastle disease virus. Avian Dis. 1970, 14, 542–549. [Google Scholar] [CrossRef]
- Hanson, R.P.; Spalatin, J. Thermostability of the hemagglutinin of Newcastle disease virus as a strain marker in epizootiologic studies. Avian Dis. 1978, 22, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Ellis, T.M.; Fenwick, S.; Edwards, J.; O’Dea, M.; Parkinson, J. Biological and genetic characterization of a low-pathogenicity avian influenza H6N2 virus originating from a healthy Eurasian coot. Arch. Virol. 2010, 155, 403–409. [Google Scholar] [CrossRef]
- Bu, Y.-W.; Yang, H.-M.; Jin, J.-H.; Zhao, J.; Xue, J.; Zhang, G.-Z. Recombinant Newcastle disease virus (NDV) La Sota expressing the haemagglutinin–neuraminidase protein of genotype VII NDV shows improved protection efficacy against NDV challenge. Avian Pathol. 2019, 48, 91–97. [Google Scholar] [CrossRef]
- Bergfeld, J.; Meers, J.; Bingham, J.; Harper, J.; Payne, J.; Lowther, S.; Marsh, G.; Tachedjian, M.; Middleton, D. An Australian Newcastle Disease virus with a virulent fusion protein cleavage site produces minimal pathogenicity in chickens. Vet. Pathol. 2017, 54, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Kattenbelt, J.A.; Stevens, M.P.; Gould, A.R. Sequence variation in the Newcastle disease virus genome. Virus Res. 2006, 116, 168–184. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, G.; Quan, P.L.; Jabado, O.J.; Conlan, S.; Hirschberg, D.L.; Liu, Y.; Zhai, J.; Renwick, N.; Hui, J.; Hegyi, H.; et al. Panmicrobial oligonucleotide array for diagnosis of infectious diseases. Emerg. Infect. Dis. 2007, 13, 73–81. [Google Scholar] [CrossRef]
- Zhao, W.; Hu, H.; Zsak, L.; Yu, Q.; Yang, Z. HN gene C-terminal extension of Newcastle disease virus is not the determinant of the enteric tropism. Virus Genes 2013, 47, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Hattori, S.; Ishida, N.; Imamura, Y.; Kawakita, M. Nucleotide sequence of the hemagglutinin-neuraminidase gene of Newcastle disease virus avirulent strain D26: Evidence for a longer coding region with a carboxyl terminal extension as compared to virulent strains. Virus Res. 1987, 8, 217–232. [Google Scholar] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Dortmans, J.; Koch, G.; Rottier, P.; Peeters, B. Virulence of Newcastle disease virus: What is known so far? Vet. Res. 2011, 42, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, H.L.; Taylor, T.L.; Dimitrov, K.M.; Sabra, M.; Afonso, C.L.; Suarez, D.L. Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet. Microbiol. 2019, 235, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.L. Virulence during Newcastle Disease Viruses Cross Species Adaptation. Viruses 2021, 13, 110. [Google Scholar] [CrossRef]
- Gould, A.R.; Kattenbelt, J.A.; Selleck, P.; Hansson, E.; Della-Porta, A.; Westbury, H.A. Virulent Newcastle disease in Australia: Molecular epidemiological analysis of viruses isolated prior to and during the outbreaks of 1998–2000. Virus Res. 2001, 77, 51–60. [Google Scholar] [CrossRef]
- Animal Health Australia. Newcastle Disease Management. Available online: https://www.animalhealthaustralia.com.au/what-we-do/endemic-disease/newcastle-disease/ (accessed on 13 February 2021).
- Gelb, J.; Fries, P.A.; Peterson, F.S. Pathogenicity and Cross-Protection of Pigeon Paramyxovirus-1 and Newcastle Disease Virus in Young Chickens. Avian Dis. 1987, 31, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Barbezange, C.; Jestin, V. Quasispecies nature of an unusual avian paramyxovirus type-1 isolated from pigeons. Virus Genes 2005, 30, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Meulemans, G.; Van den Berg, T.P.; Decaesstecker, M.; Boschmans, M. Evolution of pigeon Newcastle disease virus strains. Avian Pathol. 2002, 31, 515–519. [Google Scholar] [CrossRef]
- Collins, M.S.; Alexander, D.J.; Brockman, S.; Kemp, P.A.; Manvell, R.J. Evaluation of mouse monoclonal antibodies raised against an isolate of the variant avian paramyxovirus type 1 responsible for the current panzootic in pigeons. Arch. Virol. 1989, 104, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Stone, H.D. Efficacy of Oil-Emulsion Vaccines Prepared with Pigeon Paramyxovirus-1, Ulster, and La Sota Newcastle Disease Viruses. Avian Dis. 1989, 33, 157–162. [Google Scholar] [CrossRef]
- Hanson, R.P.; Upton, E. Heat stability of hemagglutinin of various strains of Newcastle disease virus. Proc. Soc. Exp. Biol. Med. 1949, 70, 283–287. [Google Scholar] [CrossRef]
- Ide, P.R. Virological studies of Paramyxovirus type 1 infection of pigeons. Can. Vet. J. 1987, 28, 601–603. [Google Scholar] [PubMed]
- Shirai, J.; Tsukamoto, K.; Hihara, H. Newcastle disease viruses isolated from racing pigeons in Japan. Jpn. J. Vet. Sci. 1986, 48, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Rott, R.; Klenk, H.D. Molecular basis of infectivity and pathogenicity of Newcastle disease virus. In Newcastle Disease; Alexander, D.J., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 98–112. [Google Scholar]
- Reeve, P.; Poste, G. Studies on the Cytopathogenicity of Newcastle Disease Virus: Relation between Virulence, Polykaryocytosis and Plaque Size. J. Gen. Virol. 1971, 11, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Dortmans, J.; Rottier, P.; Koch, G.; Peeters, B. The viral replication complex is associated with the virulence of Newcastle disease virus. J. Virol. 2010, 84, 10113–10120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Samal, S.K. Role of intergenic sequences in newcastle disease virus RNA transcription and pathogenesis. J. Virol. 2008, 82, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Peeters, B.P.; de Leeuw, O.S.; Koch, G.; Gielkens, A.L. Rescue of Newcastle disease virus from cloned cDNA: Evidence that cleavability of the fusion protein is a major determinant for virulence. J. Virol. 1999, 73, 5001–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortmans, J.; Koch, G.; Rottier, P.; Peeters, B. Virulence of pigeon paramyxovirus type 1 does not always correlate with the cleavability of its fusion protein. J. Gen. Virol. 2009, 90, 2746–2750. [Google Scholar] [CrossRef]
- Pearson, A.J.E.; Senne, D.A.; Alexander, D.J.; Taylor, W.D.; Peterson, L.A.; Russell, H. Characterization of Newcastle Disease Virus (Avian Paramyxovirus-1) Isolated from Pigeons. Avian Dis. 1987, 31, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.S.; Strong, I.; Alexander, D.J. Evaluation of the molecular basis of pathogenicity of the variant Newcastle disease viruses termed “pigeon PMV-1 viruses”. Arch. Virol. 1994, 134, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Dortmans, J.; Rottier, P.; Koch, G.; Peeters, B. Passaging of a Newcastle disease virus pigeon variant in chickens results in selection of viruses with mutations in the polymerase complex enhancing virus replication and virulence. J. Gen. Virol. 2011, 92, 336–345. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Pass No. | Inoculum | Number of Dead Eggs Post Inoculation (PI) | Number of Live Eggs at the End of Culture | ||||
|---|---|---|---|---|---|---|---|
| Dilution | Egg No. | D2 | D3 | D4 | D5 | ||
| 1 | NA | 3 | 1 (0) * | 1 (2) | 1 (16) (C5 **) | ||
| 2 | neat | 3 | 1 (0) | 1 (16) | 1 (64) | ||
| 3 | 10−3 | 21 | 1 (0) | 12(16 †), 6 (32 †) | 2 (64 †) (C4) | ||
| 4 | 10−3 | 36 | 7 (64 †) | 28 (128 †) | 1 (128) (C3) | ||
| 5 | 10−3 | 6 | 1 (32) | 3 (128 †) | 1 (128), 1 (256) | ||
| Isolates | Virulence | MAbs | Polyclonal Antisera | ||
|---|---|---|---|---|---|
| 617/161 | U85 | P/Aus/3/11 | V4 | ||
| P/Aus/3/11 | Unknown | 1024 | 4 | 256 | 512 |
| Texas GB | Neurotropic velogenic | <2 | 32 | 8 | 128 |
| Herts 33 | Viscerotropic velogenic | <2 | 16 | 16 | 512 |
| Komarov | Mesogenic | <2 | 512 | 64 | 1024 |
| Beaudette C | Mesogenic | <2 | 1024 | 64 | 1024 |
| B1 | Lentogenic | 2 | 256 | 32 | 512 |
| V4 | Lentogenic | <2 | 4 | 32 | 1024 |
| Strain | Original HA Titre | HA-Elution after | Elution Pattern | ||
|---|---|---|---|---|---|
| 24 h | Resuspension | 120 h | |||
| P/Aus/3/11 | 128 | 32 | 128 | <2 | Slow |
| Texas GB | 64 | 32 | 128 | <2 | Slow |
| Herts 33 | 128 | 8 | 256 | <2 | Slow |
| Komarov | 256 | 256 | 512 | <2 | Slow |
| Beaudette C | 256 | 64 | 128 | 32 | Slow |
| B1 | 512 | <2 | <2 | <2 | Rapid |
| V4 | 256 | 256 | 512 | 256 | Slow |
| Original HA Titre | Haemagglutinin Thermostability after (min) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 5 | 10 | 15 | 20 | 25 | 30 | 45 | 60 | 90 | 120 | ||
| P/Aus/3/11 | 128 | 128 | 64 | 64 | 64 | 32 | 32 | 8 | <2 | − | ||
| Texas GB | 64 | 64 | 32 | 32 | 32 | 16 | 16 | 16 | 4 | 4 | <2 | − |
| Herts 33 | 128 | 128 | 128 | 128 | 128 | <2 | <2 | − | ||||
| Komarov | 256 | 128 | 128 | 8 | <2 | − | ||||||
| Beaudette C | 256 | 256 | 256 | 128 | 64 | 64 | 64 | 64 | 64 | 64 | 32 | 32 |
| B1 | 512 | 512 | 256 | − | ||||||||
| V4 | 256 | 256 | 256 | 256 | 256 | 256 | 256 | 256 | 128 | 128 | 64 | 32 |
| Bird No. | Virus Detection (Ct) on Day Post Infection (Cloacal/Choanal Swabs) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 12 | 14 | 21 | |
| 1 | -*/31.6 ± | -/34.8 | -/36.9 | -/36.7 x | |||||||||
| 2 | -/34.0 | 35.9/34.9 | -/31.2 | -/33.6 | -/35.2 | -/- | -/- | -/- | -/- | -/- | -/- | -/38.1 | ND |
| 3 | -/32.9 | -/34.7 x | |||||||||||
| 4 | -/32.4 | -/34.0 | -/32.9 | -/33.9 x | |||||||||
| 5 | -/28.1 | -/32.7 | -/29.9 | -/28.4 | -/30.6 | -/- | 36.9/- | 37.7/- | -/- | 36.2/- | 34.2/- | 33.3/- | -/- |
| 6 | -/29.4 | -/34.3 | -/33.5 | 36.7/28.1 | -/35.6 | -/37.7 x | |||||||
| 7 | -/29.5 | -/34.4 x | |||||||||||
| 8 | -/27.4 | -/32.8 | -/32.4 | -/33.3 | -/31.9 | -/32.9 | -/30.6 | -/37.4 x | |||||
| 9 | -/32.5 | -/34.1 | -/33.0 | -/33.1 | -/36.8 | -/- | -/37.6 | -/38.1 | -/- | -/- | -/- | -/- | ND |
| 10 | -/33.3 | -/31.8 | -/35.9 | -/33.5 | -/31.7 | -/36.5 | 36.8/- | 33.4/34.8 | 33.5/36.7 | 31.7/36.6 | 38.0/- | -/- | ND |
| 11 | -/30.8 | -/35.2 | -/34.3 | -/29.8 | -/28.7 | -/31.1 x | |||||||
| 12 | -/32.5 | 37.7/34.0 | -/31.6 | -/32.7 | -/34.9 | 36.2/37.7 | -/- | -/36.9 | -/- | 37.3/- | -/- | 29.8/- | ND/- |
| 13 | -/30.4 | -/31.0 | 37.6/33.5 | -/29.2 | -/33.2 | 35.6/37.7 | -/37.0 | 36.9/− x | |||||
| +ve% | 0/100 | 15.4/100 | 9.1/100 | 9.1/100 | 0/100 | 22.2/66.7 | 28.6/42.9 | 42.9/57.1 | 20/20 | 60/20 | 40/0 | 60/20 | ND |
| Strain | F Cleavage Site | HN-Extension (Length: Sequence) | Classification | Genbank Acc. No. |
|---|---|---|---|---|
| P/Aus/3/2011 | RRQKRF | 0 | VI.2.1.1.2.2 | MN462666 |
| Aus-V4/66 | GKQGRL | 45: REARSSRLSQLREGWKDDIVSPIFCDAKNQTEYRRELESYAASWP | I.1.1 | JX524203 |
| Aus/98-1252 | RRQRRF | 9: REARSSRLS | I.1.1 | AY935493 |
| Belgium/11-08304 | RRQKRF | 0 | VI.2.1.1.2.2 | JX901123 |
| China/BJ-2013 | RRQKRF | 0 | VI.2.1.1.2.2 | KJ808819 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, S.; Bruce, K.; Stevens, V.; Wong, F.Y.K.; Wang, J.; Johnson, D.; Middleton, D.; O’Riley, K.; McCullough, S.; Williams, D.T.; et al. In Vitro and In Vivo Characterization of a Pigeon Paramyxovirus Type 1 Isolated from Domestic Pigeons in Victoria, Australia 2011. Viruses 2021, 13, 429. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030429
Shan S, Bruce K, Stevens V, Wong FYK, Wang J, Johnson D, Middleton D, O’Riley K, McCullough S, Williams DT, et al. In Vitro and In Vivo Characterization of a Pigeon Paramyxovirus Type 1 Isolated from Domestic Pigeons in Victoria, Australia 2011. Viruses. 2021; 13(3):429. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030429
Chicago/Turabian StyleShan, Songhua, Kerri Bruce, Vittoria Stevens, Frank Y. K. Wong, Jianning Wang, Dayna Johnson, Deborah Middleton, Kim O’Riley, Sam McCullough, David T. Williams, and et al. 2021. "In Vitro and In Vivo Characterization of a Pigeon Paramyxovirus Type 1 Isolated from Domestic Pigeons in Victoria, Australia 2011" Viruses 13, no. 3: 429. https://0-doi-org.brum.beds.ac.uk/10.3390/v13030429