
Isolation and Characterization of a Novel Jumbo Phage from Leaf Litter Compost and Its Suppressive Effect on Rice Seedling Rot Diseases

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Phage Isolation and Culture Conditions
2.3. Transmission Electron Microscopy
2.4. Extraction of Phage Genomic DNA and Next-Generation Sequencing
2.5. In Silico Analysis
2.6. Assessment of Disease Suppression Activity Against Rice Seedling Rot
3. Results
3.1. Isolation of FLC6 from Leaf Litter Compost and TEM Observation of Particles
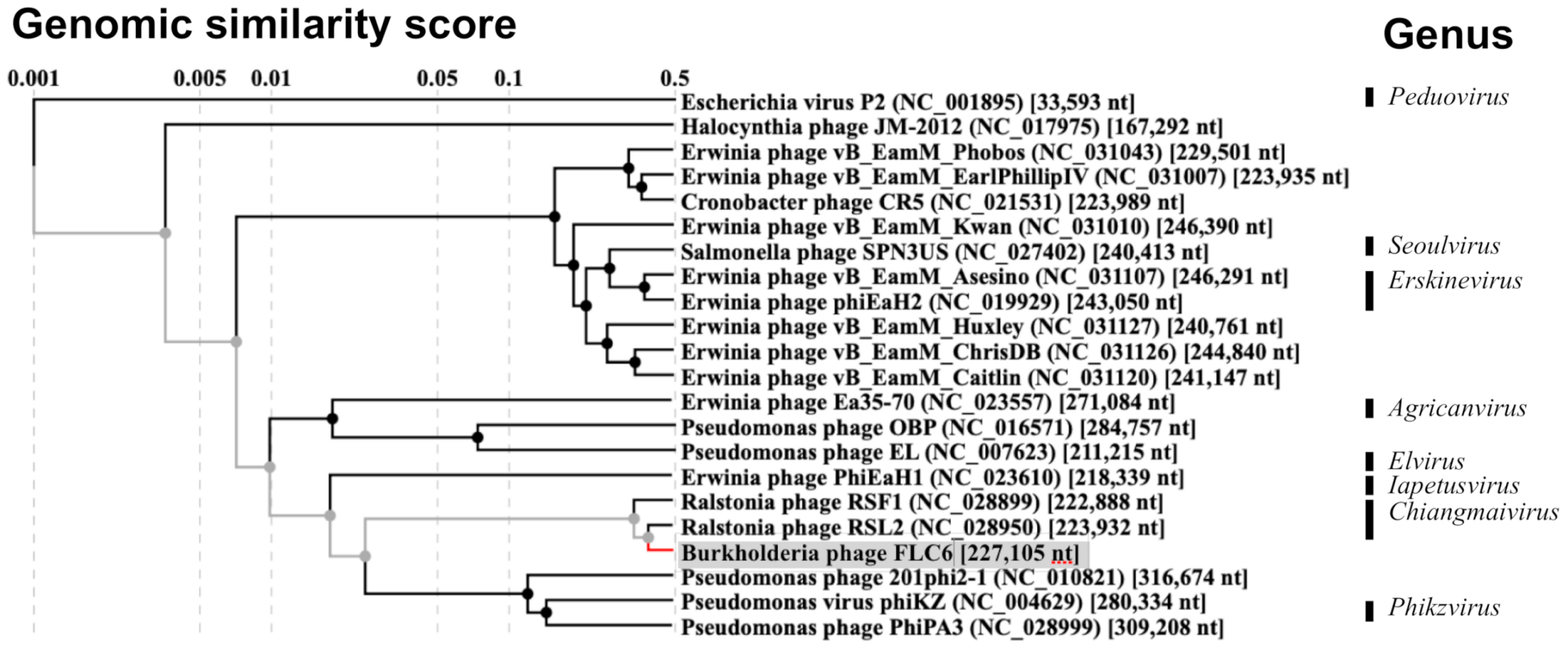
3.2. Genomic Features and Comparison of FLC6 Genome with Other Phages
3.3. Gene Annotation of Predicted Proteins Encoded on FLC6 Genome
3.4. Host Range Analysis
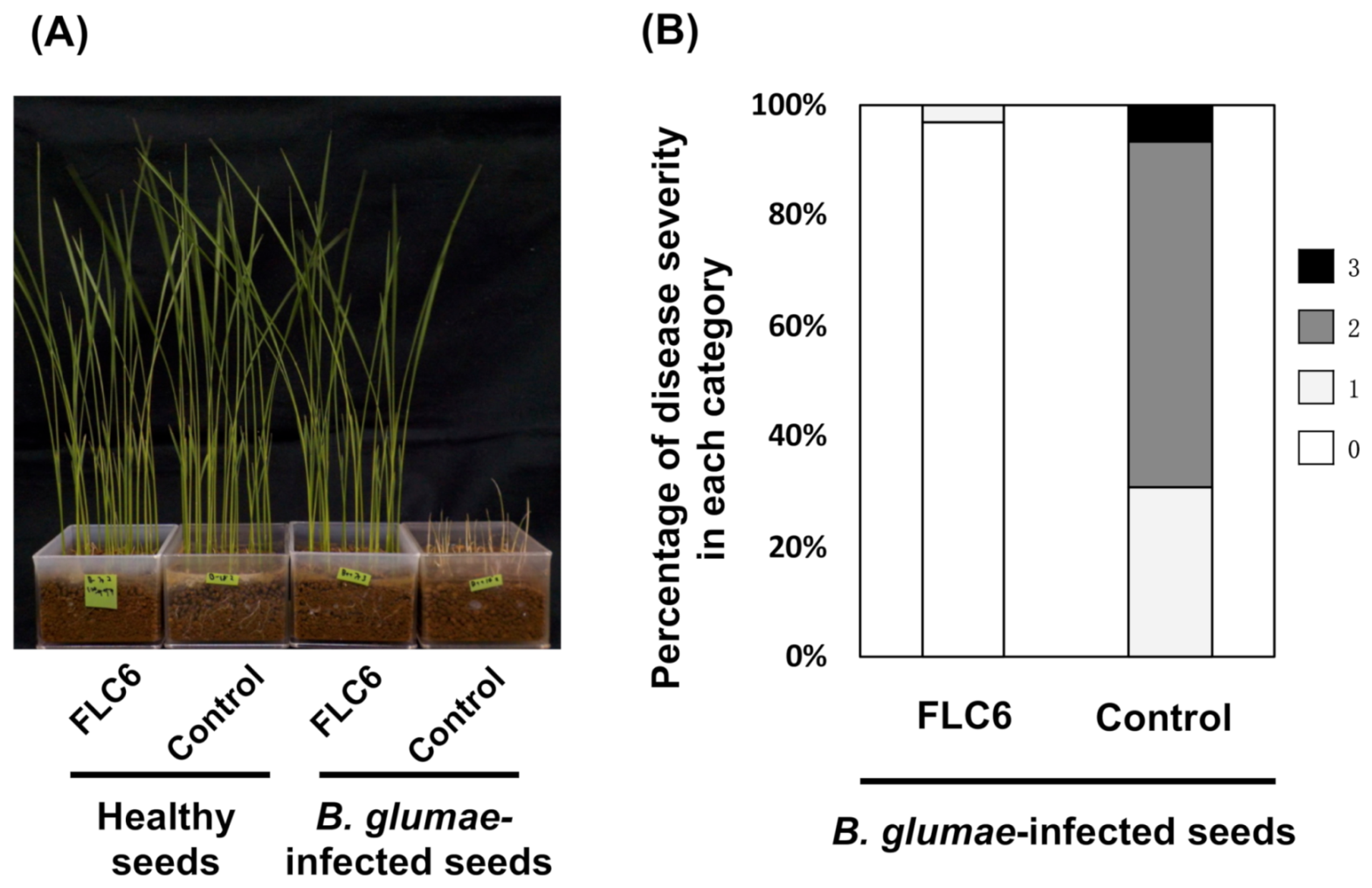
3.5. Disease Suppression Against Rice Seedling Rot by FLC6 Treatment
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orlova, E.V. Bacteriophages and their structural organisation. In Bacteriophages; Kurtböke, I., Ed.; InTech: Rijeka, Croatia, 2012; pp. 3–30. ISBN 978-953-51-0272-4. [Google Scholar]
- Ackermann, H.-W. 5500 Phages examined in the electron microscope. Arch. Virol. 2006, 152, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Genet. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.M. Bacteriophage Diversity. In Viruses of Microorganisms; Caister Academic Press: Poole, UK, 2018; pp. 145–166. [Google Scholar]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Gao, M. Jumbo bacteriophages: An overview. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Attai, H.; Boon, M.; Phillips, K.; Noben, J.-P.; Lavigne, R.; Brown, P.J.B. Larger Than Life: Isolation and Genomic Characterization of a Jumbo Phage That Infects the Bacterial Plant Pathogen, Agrobacterium tumefaciens. Front. Microbiol. 2018, 9, 1861. [Google Scholar] [CrossRef] [Green Version]
- Bhunchoth, A.; Blanc-Mathieu, R.; Mihara, T.; Nishimura, Y.; Askora, A.; Phironrit, N.; Leksomboon, C.; Chatchawankanphanich, O.; Kawasaki, T.; Nakano, M.; et al. Two asian jumbo phages, ϕRSL2 and ϕRSF1, infect Ralstonia solanacearum and show common features of ϕKZ-related phages. Virology 2016, 494, 56–66. [Google Scholar] [CrossRef]
- Day, A.; Ahn, J.; Salmond, G.P.C. Jumbo Bacteriophages Are Represented Within an Increasing Diversity of Environmental Viruses Infecting the Emerging Phytopathogen, Dickeya solani. Front. Microbiol. 2018, 9, 2169. [Google Scholar] [CrossRef]
- Dömötör, D.; Becságh, P.; Rákhely, G.; Schneider, G.; Kovács, T. Complete Genomic Sequence of Erwinia amylovora Phage PhiEaH2. J. Virol. 2012, 86, 10899. [Google Scholar] [CrossRef] [Green Version]
- Frampton, R.A.; Taylor, C.; Moreno, A.V.H.; Visnovsky, S.B.; Petty, N.K.; Pitman, A.R.; Fineran, P.C. Identification of Bacteriophages for Biocontrol of the Kiwifruit Canker Phytopathogen Pseudomonas syringae pv. actinidiae. Appl. Environ. Microbiol. 2014, 80, 2216–2228. [Google Scholar] [CrossRef] [Green Version]
- Malone, L.M.; Warring, S.L.; Jackson, S.A.; Warnecke, C.; Gardner, P.P.; Gumy, L.F.; Fineran, P.C. A jumbo phage that forms a nucleus-like structure evades CRISPR–Cas DNA targeting but is vulnerable to type III RNA-based immunity. Nat. Microbiol. 2019, 5, 48–55. [Google Scholar] [CrossRef]
- Sharma, R.; Pielstick, B.A.; Bell, K.A.; Nieman, T.B.; Stubbs, O.A.; Yeates, E.L.; Baltrus, D.A.; Grose, J.H. A Novel, Highly Related Jumbo Family of Bacteriophages That Were Isolated Against Erwinia. Front. Microbiol. 2019, 10, 1533. [Google Scholar] [CrossRef] [Green Version]
- Yagubi, A.I.; Castle, A.J.; Kropinski, A.M.; Banks, T.W.; Svircev, A.M. Complete Genome Sequence of Erwinia amylovora Bacteriophage vB_EamM_Ea35-70. Genome Announc. 2014, 2, e00413-14. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Satoh, S.; Ishikawa, H.; Fujiwara, A.; Kawasaki, T.; Fujie, M.; Ogata, H. A jumbo phage infecting the phytopathogen Ralstonia solanacearum defines a new lineage of the Myoviridae family. Virology 2010, 398, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, G.; Askora, A.; Blanc-Mathieu, R.; Kawasaki, T.; Li, Y.; Nakano, M.; Ogata, H.; Yamada, T. Xanthomonas citri jumbo phage XacN1 exhibits a wide host range and high complement of tRNA genes. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Yuan, Y.; Gao, M. Characteristics and complete genome analysis of a novel jumbo phage infecting pathogenic Bacillus pumilus causing ginger rhizome rot disease. Arch. Virol. 2016, 161, 3597–3600. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Yoshikawa, G.; Mihara, T.; Chatchawankanphanich, O.; Kawasaki, T.; Nakano, M.; Fujie, M.; Ogata, H.; Yamada, T. Replications of Two Closely Related Groups of Jumbo Phages Show Different Level of Dependence on Host-encoded RNA Polymerase. Front. Microbiol. 2017, 8, 1010. [Google Scholar] [CrossRef]
- Meczker, K.; Dömötör, D.; Vass, J.; Rákhely, G.; Schneider, G.; Kovács, T. The genome of the Erwinia amylovora phage PhiEaH1 reveals greater diversity and broadens the applicability of phages for the treatment of fire blight. FEMS Microbiol. Lett. 2014, 350, 25–27. [Google Scholar] [CrossRef] [Green Version]
- Kannan, V.R.; Bastas, K.K.; Devi, R.S. Scientific and economic impact of plant pathogenic bacteria. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; Kannan, V.R., Bastas, K.K., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 369–392. ISBN 9780429158896. [Google Scholar]
- Azegami, K.; Nishiyama, K.; Watanabe, Y.; Kadota, I.; Ohuch, A.; Fukazawa, C. Pseudomonas plantarii sp. nov., the Causal Agent of Rice Seedling Blight. Int. J. Syst. Bacteriol. 1987, 37, 475. [Google Scholar] [CrossRef] [Green Version]
- Uematsu, T.; Yoshimura, D.; Nishiyama, K.; Ibaragi, T.; Fujii, H. Pathogenic bacterium causing seedling rot of rice. Ann. Phytopathol. Soc. Jpn. 1976, 42, 464–471. [Google Scholar] [CrossRef]
- Adachi, N.; Tsukamoto, S.; Inoue, Y.; Azegami, K. Control of Bacterial Seedling Rot and Seedling Blight of Rice by Bacteriophage. Plant Dis. 2012, 96, 1033–1036. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Miyashita, S.; Ando, S.; Ito, K.; Fukuhara, T.; Kormelink, R.; Takahashi, H. Complete genomic sequence of a novel phytopathogenic Burkholderia phage isolated from fallen leaf compost. Arch. Virol. 2021, 166, 313–316. [Google Scholar] [CrossRef]
- Azegami, K. Burkholderia glumae and Burkholderia plantarii, the pathogens of bacterial grain rot of rice and bacterial seedling blight of rice, respectively. MAFF Microorg. Genet. Resour. Man. 2009, 26, 1–24. [Google Scholar]
- Horita, M.; Tsuchiya, K. Causal agent of bacterial wilt disease, Ralstonia solanacearum. MAFF Microorg. Genet. Resour. Man. 2012, 12, 1–34. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual (3-Volume Set); Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; ISBN 978-087969577-4. [Google Scholar]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.; Korobeynikov, A.; Lapidus, A.; Prjibelsky, A.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling genomes and mini-metagenomes from highly chimeric reads. In Research in Computational Molecular Biology; RECOMB 2013; Lecture Notes in Computer Science; Deng, M., Jiang, R., Sun, F., Zhang, X., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 158–170. ISBN 9783642371943. [Google Scholar]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Staerfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Ando, S.; Ito, T.; Kanno, T.; Kobayashi, T.; Morikawa, T.; Honda, K.-I.; Tsushima, S.; Takahashi, H. Impact of organic crop management on suppression of bacterial seedling diseases in rice. Org. Agric. 2014, 4, 187–196. [Google Scholar] [CrossRef]
- Takahashi, H.; Matsushita, Y.; Ito, T.; Nakai, Y.; Nanzyo, M.; Kobayashi, T.; Iwaishi, S.; Hashimoto, T.; Miyashita, S.; Morikawa, T.; et al. Comparative analysis of microbial diversity and bacterial seedling disease-suppressive activity in organic-farmed and standardized commercial conventional soils for rice nursery cultivation. J. Phytopathol. 2018, 166, 249–264. [Google Scholar] [CrossRef]
- Ando, S.; Takahashi, H. Analysis of the disease-suppressing effects of microorganisms included in nursery soils for organic farming of rice. J. Integr. Agric. 2018, 15, 51–54. [Google Scholar]
- Hikichi, Y.; Egami, H.; Oguri, Y.; Okuno, T. Fitness for Survival of Burkholderia glumae Resistant to Oxolinic Acid in Rice Plants. Jpn. J. Phytopathol. 1998, 64, 147–152. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wei, Z.; Yang, K.; Wang, J.; Jousset, A.; Xu, Y.; Shen, Q.; Friman, V.-P. Phage combination therapies for bacterial wilt disease in tomato. Nat. Biotechnol. 2019, 37, 1513–1520. [Google Scholar] [CrossRef]
- Álvarez, B.; López, M.M.; Biosca, E.G. Biocontrol of the Major Plant Pathogen Ralstonia solanacearum in Irrigation Water and Host Plants by Novel Waterborne Lytic Bacteriophages. Front. Microbiol. 2019, 10, 2813. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, B.; Biosca, E.G. Bacteriophage-Based Bacterial Wilt Biocontrol for an Environmentally Sustainable Agriculture. Front. Plant Sci. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, A.; Fujisawa, M.; Hamasaki, R.; Kawasaki, T.; Fujie, M.; Yamada, T. Biocontrol of Ralstonia solanacearum by Treatment with Lytic Bacteriophages. Appl. Environ. Microbiol. 2011, 77, 4155–4162. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phage | NCBI RefSeq Accession | FLC6 gp175 | FLC6 gp176 | ||
|---|---|---|---|---|---|
| Sequence Identity 1 (%) | Query Coverage 1 (%) | Sequence Identity 1 (%) | Query Coverage 1 (%) | ||
| RSL2 | YP_009213007.1 | 84 | 100 | 92 | 54 |
| RSF1 | YP_009207973.2 | 76 | 100 | 95 | 50 |
| Species | Strain | Sensitivity 2 |
|---|---|---|
| B. glumae | MAFF106619 | + |
| B. glumae | MAFF301169 1 | + |
| B. glumae | MAFF302417 | + |
| B. glumae | MAFF302552 | + |
| B. glumae | MAFF302746 | + |
| B. plantarii | MAFF106727 | + |
| B. plantarii | MAFF302466 | + |
| B. plantarii | MAFF302475 | + |
| B. plantarii | MAFF302909 | + |
| B. plantarii | MAFF302912 | + |
| B. plantarii | MAFF302936 | + |
| R. pseudosolanacearum | MAFF106603 | + |
| R. pseudosolanacearum | MAFF106611 | + |
| R. pseudosolanacearum | MAFF211270 | − |
| R. pseudosolanacearum | MAFF301485 | − |
| R. syzygii subsp. indonesiensis | MAFF211271 | − |
| R. syzygii subsp. indonesiensis | MAFF327032 | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sasaki, R.; Miyashita, S.; Ando, S.; Ito, K.; Fukuhara, T.; Takahashi, H. Isolation and Characterization of a Novel Jumbo Phage from Leaf Litter Compost and Its Suppressive Effect on Rice Seedling Rot Diseases. Viruses 2021, 13, 591. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040591
Sasaki R, Miyashita S, Ando S, Ito K, Fukuhara T, Takahashi H. Isolation and Characterization of a Novel Jumbo Phage from Leaf Litter Compost and Its Suppressive Effect on Rice Seedling Rot Diseases. Viruses. 2021; 13(4):591. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040591
Chicago/Turabian StyleSasaki, Ryota, Shuhei Miyashita, Sugihiro Ando, Kumiko Ito, Toshiyuki Fukuhara, and Hideki Takahashi. 2021. "Isolation and Characterization of a Novel Jumbo Phage from Leaf Litter Compost and Its Suppressive Effect on Rice Seedling Rot Diseases" Viruses 13, no. 4: 591. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040591