
High Heterogeneity of Echoviruses in Brazilian Children with Acute Gastroenteritis

 ,
,  , , , , , ,
, , , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Aspects
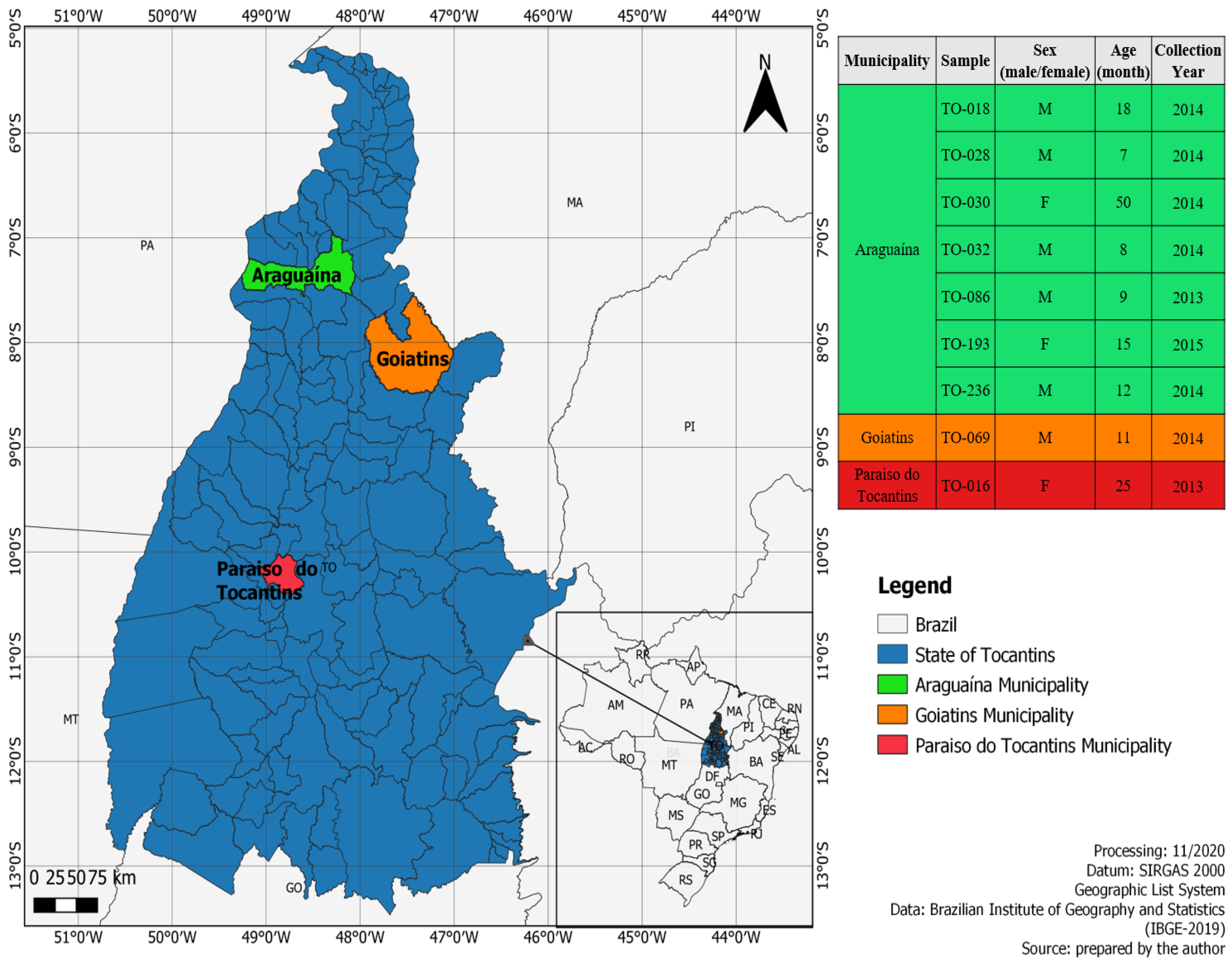
2.2. Study Population and Specimen Collection
2.3. Sample Screening
2.4. Viral Metagenomics
2.5. Molecular Typing of Echovirus
2.6. Alignments and Phylogenetic Analysis
2.7. Evolutionary Distances
2.8. Recombination Analysis
2.9. Accession Numbers
3. Results
3.1. Echovirus Identification
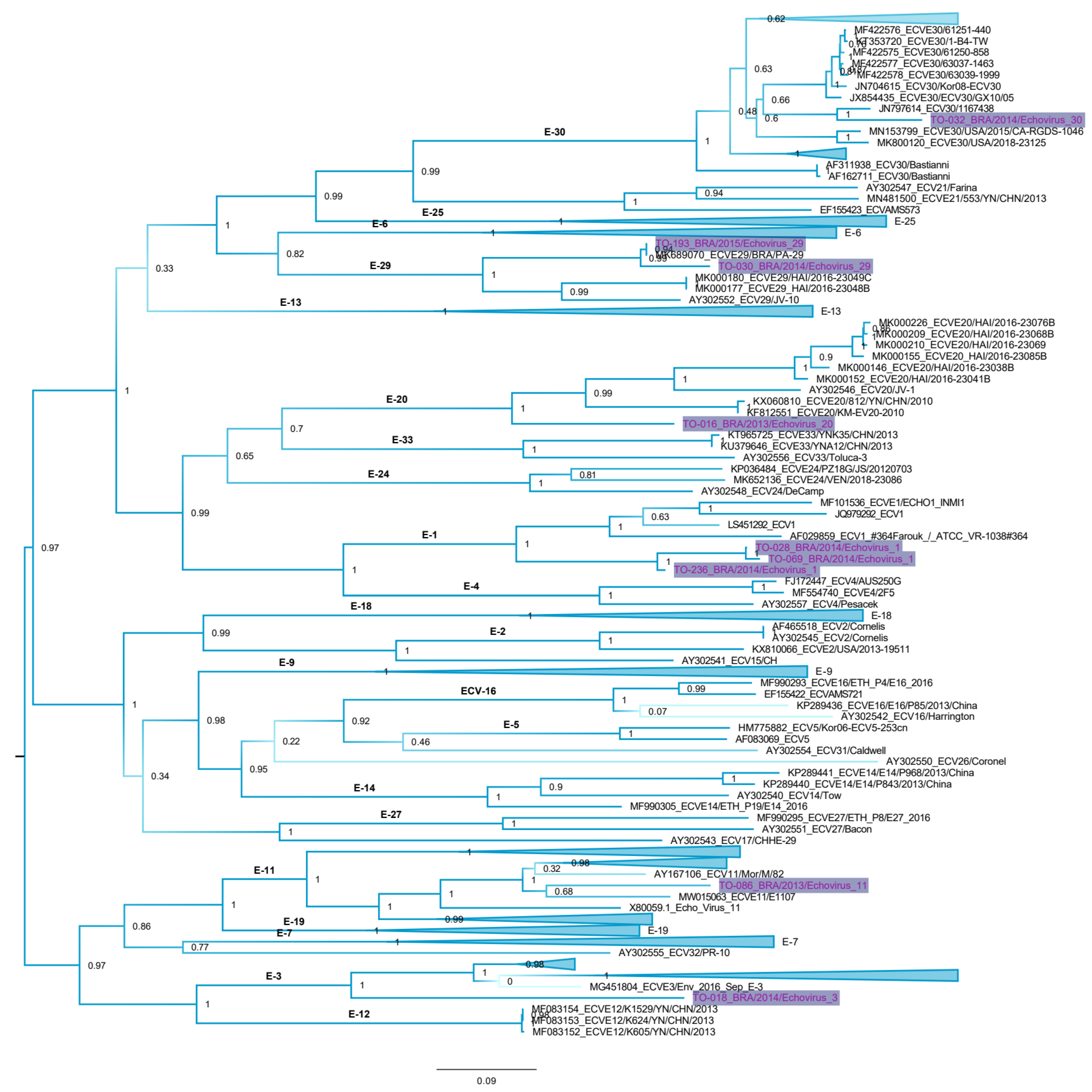
3.2. Echovirus Genotyping
3.3. Genetic Distances and Phylogenetic Inferences
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/picornaviridae/681/genus-enterovirus (accessed on 10 November 2020).
- Committee ON the Echo Viruses Enteric Cytopathogenic Human Orphan (ECHO) Viruses. Science 1955, 122, 1187–1188. [CrossRef]
- Liu, H.; Zhang, J.; Zhao, Y.; Zhang, H.; Lin, K.; Sun, H.; Huang, X.; Yang, Z.; Ma, S. Molecular Characterization of Echovirus 12 Strains Isolated from Healthy Children in China. Sci. Rep. 2018, 8, 11716. [Google Scholar] [CrossRef] [PubMed]
- Maruo, Y.; Nakanishi, M.; Suzuki, Y.; Kaneshi, Y.; Terashita, Y.; Narugami, M.; Takahashi, M.; Kato, S.; Suzuki, R.; Goto, A.; et al. Outbreak of Aseptic Meningitis Caused by Echovirus 30 in Kushiro, Japan in 2017. J. Clin. Virol. 2019, 116, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Faleye, T.O.C.; Adewumi, O.M.; Adeniji, J.A. Genomic Characterization of an Echovirus 7 Isolate of Southeast Asian Ancestry Recovered from a Child in Nigeria with Acute Flaccid Paralysis. Microbiol. Resour. Announc. 2019, 8, e00830-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchs, A.; Leal, E.; Tardy, K.; de Milagres, F.A.P.; Komninakis, S.V.; Brustulin, R.; da Teles, M.A.R.; Lobato, M.C.A.B.S.; das Chagas, R.T.; Abrão, M.F.N.S.; et al. The Rare Enterovirus C99 and Echovirus 29 Strains in Brazil: Potential Risks Associated to Silent Circulation. Mem. Inst. Oswaldo Cruz. 2019, 114, e190160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khetsuriani, N.; Lamonte-Fowlkes, A.; Oberst, S.; Pallansch, M.A. Centers for Disease Control and Prevention Enterovirus Surveillance—United States, 1970–2005. MMWR Surveill. Summ. 2006, 55, 1–20. [Google Scholar]
- Oberste, M.S.; Maher, K.; Kilpatrick, D.R.; Pallansch, M.A. Molecular Evolution of the Human Enteroviruses: Correlation of Serotype with VP1 Sequence and Application to Picornavirus Classification. J. Virol. 1999, 73, 1941–1948. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, N.; Semler, B.L.; Rothberg, P.G.; Larsen, G.R.; Adler, C.J.; Dorner, A.J.; Emini, E.A.; Hanecak, R.; Lee, J.J.; van der Werf, S.; et al. Primary Structure, Gene Organization and Polypeptide Expression of Poliovirus RNA. Nature 1981, 291, 547–553. [Google Scholar] [CrossRef]
- Hyypia, T.; Hovi, T.; Knowles, N.J.; Stanway, G. Classification of Enteroviruses Based on Molecular and Biological Properties. J. Gen. Virol. 1997, 78, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nikolaidis, M.; Mimouli, K.; Kyriakopoulou, Z.; Tsimpidis, M.; Tsakogiannis, D.; Markoulatos, P.; Amoutzias, G.D. Large-Scale Genomic Analysis Reveals Recurrent Patterns of Intertypic Recombination in Human Enteroviruses. Virology 2019, 526, 72–80. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Zhao, Y.; Zhang, H.; Sun, H.; Huang, X.; Yang, Z.; Liu, J.; Ma, S. Identification of a New Recombinant Strain of Echovirus 33 from Children with Hand, Foot, and Mouth Disease Complicated by Meningitis in Yunnan, China. Virol. J. 2019, 16, 63. [Google Scholar] [CrossRef] [Green Version]
- Bujaki, E.; Farkas, Á.; Rigó, Z.; Takács, M. Distribution of Enterovirus Genotypes Detected in Clinical Samples in Hungary, 2010–2018. Acta Microbiol. Immunol. Hung. 2020. [Google Scholar] [CrossRef] [PubMed]
- Thoelen, I.; Lemey, P.; Van Der Donck, I.; Beuselinck, K.; Lindberg, A.M.; Van Ranst, M. Molecular Typing and Epidemiology of Enteroviruses Identified from an Outbreak of Aseptic Meningitis in Belgium during the Summer of 2000. J. Med. Virol. 2003, 70, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Kmetzsch, C.I.; Balkie, E.M.R.; Monteiro, A.; Costa, E.V.; dos Santos, G.P.L.; da ilva, E.E. Echovirus 13 Aseptic Meningitis, Brazil. Emerg. Infect. Dis. 2006, 12, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.C.; Estofolete, C.F.; de Aguiar Milhim, B.H.G.; Augusto, M.T.; Zini, N.; da Silva, G.C.D.; Ferraz-Junior, H.C.; Brienze, V.M.S.; Liso, E.; Cunha, M.S.; et al. Enteric Viruses Circulating in Undiagnosed Central Nervous System Infections at Tertiary Hospital in São José Do Rio Preto, São Paulo, Brazil. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Luchs, A.; Russo, D.H.; Cilli, A.; Costa, F.F.; Morillo, S.G.; Machado, B.C.; Pellini, A.C.G.; de Cássia Compagnoli Carmona, R.; Timenetsky, M. do C.S.T. Echovirus 6 Associated to Aseptic Meningitis Outbreak, in São Joaquim Da Barra, São Paulo, Brazil. Braz. J. Microbiol. 2008, 39, 28–31. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.H.; Luchs, A.; Machado, B.C.; de Carmona, R.C.; do Timenetsky, M.C.S. Echovirus 4 Associated to Hand, Foot and Mouth Disease. Rev. Inst. Med. Trop. Sao Paulo 2006, 48, 197–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, I.P.; Burlandy, F.M.; Lima, S.T.S.; Maximo, A.C.B.; Figueiredo, M.A.A.; Maia, Z.; da Silva, E.E. Echovirus 30 Detection in an Outbreak of Acute Myalgia and Rhabdomyolysis, Brazil 2016–2017. Clin. Microbiol. Infect. 2019, 25, 252.e5–252.e8. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.P.L.; da Costa, E.V.; Tavares, F.N.; da Costa, L.J.; da Silva, E.E. Genetic Diversity of Echovirus 30 Involved in Aseptic Meningitis Cases in Brazil (1998–2008). J. Med. Virol. 2011, 83, 2164–2171. [Google Scholar] [CrossRef]
- Piantadosi, A.; Mukerji, S.S.; Chitneni, P.; Cho, T.A.; Cosimi, L.A.; Hung, D.T.; Goldberg, M.B.; Sabeti, P.C.; Kuritzkes, D.R.; Grad, Y.H. Metagenomic Sequencing of an Echovirus 30 Genome From Cerebrospinal Fluid of a Patient With Aseptic Meningitis and Orchitis. Open Forum Infect. Dis. 2017, 4, ofx138. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-X.; Li, W.; Zhou, J.; Zhang, B.; Feng, Y.; Xu, C.-P.; Lu, Y.-Y.; Holmes, E.C.; Shi, M. High Resolution Metagenomic Characterization of Complex Infectomes in Paediatric Acute Respiratory Infection. Sci. Rep. 2020, 10, 3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, A.S.A.; Luchs, A.; Leal, É.; de Milagres, F.A.P.; Komninakis, S.V.; Gill, D.E.; Lobato, M.C.A.B.S.; Brustulin, R.; das Chagas, R.T.; de Abrão, M.F.N.D.S.; et al. Complete Genome Sequences of Six Human Bocavirus Strains from Patients with Acute Gastroenteritis in the North Region of Brazil. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cilli, A.; Luchs, A.; Leal, E.; Gill, D.; de Milagres, F.A.P.; Komninakis, S.V.; Brustulin, R.; da Teles, M.A.R.; Lobato, M.C.A.B.S.; das Chagas, R.T.; et al. Human Sapovirus GI.2 and GI.3 from Children with Acute Gastroenteritis in Northern Brazil. Mem. Inst. Oswaldo Cruz. 2019, 114, e180574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tahmasebi, R.; Luchs, A.; Tardy, K.; Hefford, P.M.; Tinker, R.J.; Eilami, O.; de Padua Milagres, F.A.; Brustulin, R.; da Teles, M.A.R.; Dos Santos Morais, V.; et al. Viral Gastroenteritis in Tocantins, Brazil: Characterizing the Diversity of Human Adenovirus F through next-Generation Sequencing and Bioinformatics. J. Gen. Virol. 2020, 101, 1280–1288. [Google Scholar] [CrossRef]
- Silva-Sales, M.; Leal, E.; de Milagres, F.A.P.; Brustulin, R.; Morais, V.D.S.; Marcatti, R.; Araújo, E.L.L.; Witkin, S.S.; Deng, X.; Sabino, E.C.; et al. Genomic Constellation of Human Rotavirus A Strains Identified in Northern Brazil: A 6-Year Follow-up (2010–2016). Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e98. [Google Scholar] [CrossRef]
- Charlys da Costa, A.; Thézé, J.; Komninakis, S.C.V.; Sanz-Duro, R.L.; Felinto, M.R.L.; Moura, L.C.C.; de Barroso, I.M.O.; Santos, L.E.C.; de Nunes, M.A.L.; Moura, A.A.; et al. Spread of Chikungunya Virus East/Central/South African Genotype in Northeast Brazil. Emerg. Infect. Dis. 2017, 23, 1742–1744. [Google Scholar] [CrossRef] [Green Version]
- da Costa, A.C.; Leal, E.; Gill, D.; de Milagres, F.A.P.; Komninakis, S.V.; Brustulin, R.; da Teles, M.A.R.; Lobato, M.C.A.B.S.; das Chagas, R.T.; de Abrão, M.F.N.D.S.; et al. Discovery of Cucumis Melo Endornavirus by Deep Sequencing of Human Stool Samples in Brazil. Virus Genes 2019, 55, 332–338. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, G.; Luchs, A.; Milagres, F.; Komninakis, S.; Gill, D.; Lobato, M.; Brustulin, R.; Chagas, R.; Abrão, M.; Soares, C.; et al. Detection and Characterization of Enterovirus B73 from a Child in Brazil. Viruses 2018, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Naccache, S.N.; Ng, T.; Federman, S.; Li, L.; Chiu, C.Y.; Delwart, E.L. An Ensemble Strategy That Significantly Improves de Novo Assembly of Microbial Genomes from Metagenomic Next-Generation Sequencing Data. Nucleic Acids Res. 2015, 43, e46. [Google Scholar] [CrossRef] [Green Version]
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.v.d.; Peñaranda, S.; Oberste, M.S.; Vinjé, J.; Koopmans, M. An Automated Genotyping Tool for Enteroviruses and Noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A Multiple Sequence Alignment Method with Reduced Time and Space Complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, H.A.; Strimmer, K.; Vingron, M.; von Haeseler, A. TREE-PUZZLE: Maximum Likelihood Phylogenetic Analysis Using Quartets and Parallel Computing. Bioinformatics 2002, 18, 502–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Evolutionary Trees from DNA Sequences: A Maximum Likelihood Approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1. [Google Scholar] [CrossRef] [Green Version]
- McVean, G.A.T. The Fine-Scale Structure of Recombination Rate Variation in the Human Genome. Science 2004, 304, 581–584. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Lin, X.; Ji, F.; Xiong, P.; Liu, Y.; Wang, S.; Chen, P.; Xu, Q.; Zhang, L.; Tao, Z.; et al. Prevalence and Bayesian Phylogenetics of Enteroviruses Derived From Environmental Surveillance Around Polio Vaccine Switch Period in Shandong Province, China. Food Environ. Virol. 2020, 12, 321–332. [Google Scholar] [CrossRef]
- de Compagnoli Carmona, R.C.; Caetano Machado, B.; de Sousa, C.A.; Vieira, H.R.; Moraes Alves, M.R.; de Souza, K.A.F.; de Souza Gregório, D.; Costa Vilanova, B.; do Sampaio Tavares Timenetsky, M.C. Distribution of Species Enterovirus B in Patients with Central Nervous System Infections in São Paulo State, Brazil. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- de Castro, C.M.O.; Oliveira, D.S.; Macedo, O.; Lima, M.J.L.; Santana, M.B.; Wanzeller, A.L.M.; da Silveira, E.; de Gomes, M.L.C. Echovirus 30 Associated with Cases of Aseptic Meningitis in State of Pará, Northern Brazil. Mem. Inst. Oswaldo Cruz. 2009, 104, 444–450. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, C.A.; Luchs, A.; Russo, D.H.; de Cassia Compagnoli Carmona, R.; Afonso, A.M.S.; de Oliveira, M.I.; Curti, S.P.; de Moraes, J.C.; Toscano, C.M.; Ciccone, F.H.; et al. Rubella Virus Genotype 1G and Echovirus 9 as Etiologic Agents of Exanthematous Diseases in Brazil: Insights from Phylogenetic Analysis. Arch. Virol. 2014, 159, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, E.; Sousa, I.; Burlandy, F.; Costa, E.; Dias, A.; Serrano, R.; Oliveira, M.; Lopes, R.; Debur, M.; Burger, M.; et al. Identification and Phylogenetic Characterization of Human Enteroviruses Isolated from Cases of Aseptic Meningitis in Brazil, 2013–2017. Viruses 2019, 11, 690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, R.S.; de Sousa, I.P.; Monteiro, J.C.; Ferreira, J.L.; Dos Santos Alves, J.C.; Tavares, F.N. Detection and Identification of Enteroviruses Circulating in Children with Acute Gastroenteritis in Pará State, Northern Brazil (2010–2011). Virol. J. 2020, 17, 156. [Google Scholar] [CrossRef]
- Sousa, I.P.; de Oliveira, M.L.A.; Burlandy, F.M.; Machado, R.S.; Oliveira, S.S.; Tavares, F.N.; Gomes-Neto, F.; da Costa, E.V.; da Silva, E.E. Molecular Characterization and Epidemiological Aspects of Non-Polio Enteroviruses Isolated from Acute Flaccid Paralysis in Brazil: A Historical Series (2005–2017). Emerg. Microbes. Infect. 2020, 9, 2536–2546. [Google Scholar] [CrossRef]
- Dos Santos, G.P.L.; Skraba, I.; Oliveira, D.; Lima, A.A.F.; de Melo, M.M.M.; Kmetzsch, C.I.; da Costa, E.V.; da Silva, E.E. Enterovirus Meningitis in Brazil, 1998–2003. J. Med. Virol. 2006, 78, 98–104. [Google Scholar] [CrossRef]
- Van der Sanden, S.M.G.; Koopmans, M.P.G.; van der Avoort, H.G.A.M. Detection of Human Enteroviruses and Parechoviruses as Part of the National Enterovirus Surveillance in the Netherlands, 1996–2011. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1525–1531. [Google Scholar] [CrossRef]
- Suppiah, J.; Saraswathy, T.S.; Amry, K.; Yusof, A.; Saat, Z. Echovirus Serotypes Circulating in Malaysia from 2002 to 2013. Asian Pac. J. Trop. Med. 2016, 9, 252–255. [Google Scholar] [CrossRef]
- Leendertse, M.; van Vugt, M.; Benschop, K.S.; van Dijk, K.; Minnaar, R.P.; van Eijk, H.W.M.; Hodiamont, C.J.; Wolthers, K.C. Pleurodynia Caused by an Echovirus 1 Brought Back from the Tropics. J. Clin. Virol. 2013, 58, 490–493. [Google Scholar] [CrossRef] [Green Version]
- Bell, E.J.; Grist, N.R. Echoviruses, Carditis, and Acute Pleurodynia. Lancet 1970, 1, 326–328. [Google Scholar] [CrossRef]
- Sekiguchi, J.; Kubo, H.; Kaida, A.; Takakura, K.-I.; Goto, K.; Hase, A.; Iritani, N. High Incidence of Human Echovirus Type 3 among Children in Osaka, Japan during the Summer of 2010. Jpn. J. Infect. Dis. 2011, 64, 533–535. [Google Scholar] [PubMed]
- Pouremamali, A.; Makavndi, M.; Samarbafzadeh, A.; Neisi, N.; Rasti, M.; Shamsizadeh, A.; Teimoori, A.; Haj, M.S.; Nashibi, R.; Salmanzadeh, S.; et al. Human Rhinoviruses A9, A49, B14 and Echovirus 3, 9 among the Patients with Acute Respiratory Infection. Epidemiol. Mikrobiol. Imunol. 2018, 67, 18–23. [Google Scholar] [PubMed]
- Maguire, H.C.; Atkinson, P.; Sharland, M.; Bendig, J. Enterovirus Infections in England and Wales: Laboratory Surveillance Data: 1975 to 1994. Commun. Dis. Public Health 1999, 2, 122–125. [Google Scholar]
- Antona, D.; Lévêque, N.; Chomel, J.J.; Dubrou, S.; Lévy-Bruhl, D.; Lina, B. Surveillance of Enteroviruses in France, 2000–2004. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 403–412. [Google Scholar] [CrossRef]
- Saraswathy, T.S.; Khairullah, N.S.; Sinniah, M.; Fauziah, M.K.; Apandi, M.Y.; Shamsuddin, M. Laboratory Acute Flaccid Paralysis Surveillance in Malaysia: A Decade of Commitment to the WHO Global Polio Eradication Initiative. Southeast Asian J. Trop. Med. Public Health 2004, 35, 421–424. [Google Scholar]
- Bahri, O.; Rezig, D.; Nejma-Oueslati, B.B.; Yahia, A.B.; Sassi, J.B.; Hogga, N.; Sadraoui, A.; Triki, H. Enteroviruses in Tunisia: Virological Surveillance over 12 Years (1992–2003). J. Med. Microbiol. 2005, 54, 63–69. [Google Scholar] [CrossRef]
- Khetsuriani, N.; Kutateladze, T.; Zangaladze, E.; Shutkova, T.; Peñaranda, S.; Nix, W.A.; Pallansch, M.A.; Oberste, M.S. High Degree of Genetic Diversity of Non-Polio Enteroviruses Identified in Georgia by Environmental and Clinical Surveillance, 2002–2005. J. Med. Microbiol. 2010, 59, 1340–1347. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Yan, D.; Chen, L.; Zhang, Y.; Song, Y.; Zhu, S.; Ji, T.; Zhou, W.; Gan, F.; Wang, X.; et al. Multiple Genotypes of Echovirus 11 Circulated in Mainland China between 1994 and 2017. Sci. Rep. 2019, 9, 10583. [Google Scholar] [CrossRef]
- Chen, J.-H.; Chiu, N.-C.; Chang, J.-H.; Huang, F.-Y.; Wu, K.-B.; Lin, T.-L. A Neonatal Echovirus 11 Outbreak in an Obstetric Clinic. J. Microbiol. Immunol. Infect. 2005, 38, 332–337. [Google Scholar]
- Chevaliez, S.; Szendröi, A.; Caro, V.; Balanant, J.; Guillot, S.; Berencsi, G.; Delpeyroux, F. Molecular Comparison of Echovirus 11 Strains Circulating in Europe during an Epidemic of Multisystem Hemorrhagic Disease of Infants Indicates That Evolution Generally Occurs by Recombination. Virology 2004, 325, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.; Koptides, D.; Tryfonos, C.; Christodoulou, C. Molecular Typing of Enteroviruses Associated with Viral Meningitis in Cyprus, 2000–2002. J. Med. Microbiol. 2006, 55, 1035–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrerizo, M.; Echevarria, J.E.; González, I.; de Miguel, T.; Trallero, G. Molecular Epidemiological Study of HEV-B Enteroviruses Involved in the Increase in Meningitis Cases Occurred in Spain during 2006. J. Med. Virol. 2008, 80, 1018–1024. [Google Scholar] [CrossRef]
- Rao, C.D.; Yergolkar, P.; Shankarappa, K.S. Antigenic Diversity of Enteroviruses Associated with Nonpolio Acute Flaccid Paralysis, India, 2007–2009. Emerg. Infect. Dis. 2012, 18, 1833–1840. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Enterovirus Surveillance—United States, 2002–2004. MMWR Morb. Mortal. Wkly. Rep. 2006, 55, 153–156. [Google Scholar]
- Fernandez-Garcia, M.D.; Kebe, O.; Fall, A.D.; Ndiaye, K. Identification and Molecular Characterization of Non-Polio Enteroviruses from Children with Acute Flaccid Paralysis in West Africa, 2013–2014. Sci. Rep. 2017, 7, 3808. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.; Kern, J.; Bell, J.A. Observations on a Group of Viruses (JV-5, JV-6 AND JV-10) Comprising a Newly Recognized Enterovirus Serotype. Am. J. Hyg. 1964, 79, 7–15. [Google Scholar] [CrossRef]
- Oberste, M.S.; Maher, K.; Williams, A.J.; Dybdahl-Sissoko, N.; Brown, B.A.; Gookin, M.S.; Peñaranda, S.; Mishrik, N.; Uddin, M.; Pallansch, M.A. Species-Specific RT-PCR Amplification of Human Enteroviruses: A Tool for Rapid Species Identification of Uncharacterized Enteroviruses. J. Gen. Virol. 2006, 87, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; Casas, I.; Cisterna, D.; Trallero, G.; Tenorio, A.; Freire, C. Molecular Epidemiology of Echovirus 30: Temporal Circulation and Prevalence of SingleLineages. J. Virol. 2002, 76, 4940–4949. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, C.; Hovi, T.; Mulders, M.N. Molecular Epidemiology of Echovirus 30 in Europe: Succession of Dominant Sublineages within a Single Major Genotype. Arch. Virol. 2001, 146, 521–537. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Age (Months) | Gender | Municipality | State | Year | Genotype | Strain | Symptoms | Co-Infections (Viruses) | Length (nt) | BLAST Score | Accession Numbers |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 25 | F | Paraíso do Tocantins | TO | 2013 | E20 | TO-016 | Diarrhea | RVA/NoV/E99 | 7339 | 80% | MW775340 |
| 18 | M | Araguaína | TO | 2014 | E3 | TO-018 | Diarrhea | RVA/HAdV | 7264 | 81% | MW775341 |
| 7 | M | Araguaína | TO | 2014 | E1 | TO-028 | Diarrhea | RVA/HAdV | 7401 | 79% | MW775342 |
| 50 | F | Araguaína | TO | 2014 | E29 | TO-030 | Diarrhea | RVA/HAdV/PeV | 7230 | 79% | MW775343 |
| 8 | M | Araguaína | TO | 2014 | E30 | TO-032 | Diarrhea, vomiting and fever | NoV/HAdV/HAstV | 7562 | 83% | MW775344 |
| 11 | M | Goiatins | TO | 2014 | E1 | TO-069 | Diarrhea and fever | RVA/HAdV | 7418 | 79% | MW775345 |
| 9 | M | Araguaína | TO | 2013 | E11 | TO-086 | Diarrhea, vomiting and fever | RVA/NoV/HAdV | 7427 | 82% | MW775346 |
| 15 | F | Araguaína | TO | 2015 | E29 | TO-193 | Diarrhea | SAFV | 7412 | 80% | MW775347 |
| 12 | M | Araguaína | TO | 2014 | E1 | TO-236 | Diarrhea vomiting and fever | RVA | 7375 | 77% | MW775348 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
do Socorro Fôro Ramos, E.; Rosa, U.A.; de Oliveira Ribeiro, G.; Villanova, F.; de Pádua Milagres, F.A.; Brustulin, R.; dos Santos Morais, V.; Bertanhe, M.; Marcatti, R.; Araújo, E.L.L.; et al. High Heterogeneity of Echoviruses in Brazilian Children with Acute Gastroenteritis. Viruses 2021, 13, 595. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040595
do Socorro Fôro Ramos E, Rosa UA, de Oliveira Ribeiro G, Villanova F, de Pádua Milagres FA, Brustulin R, dos Santos Morais V, Bertanhe M, Marcatti R, Araújo ELL, et al. High Heterogeneity of Echoviruses in Brazilian Children with Acute Gastroenteritis. Viruses. 2021; 13(4):595. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040595
Chicago/Turabian Styledo Socorro Fôro Ramos, Endrya, Ulisses Alves Rosa, Geovani de Oliveira Ribeiro, Fabiola Villanova, Flávio Augusto de Pádua Milagres, Rafael Brustulin, Vanessa dos Santos Morais, Mayara Bertanhe, Roberta Marcatti, Emerson Luiz Lima Araújo, and et al. 2021. "High Heterogeneity of Echoviruses in Brazilian Children with Acute Gastroenteritis" Viruses 13, no. 4: 595. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040595