
Quinine Inhibits Infection of Human Cell Lines with SARS-CoV-2

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Cell Culture
2.3. Determination of the Number of Viral RNA Copies from Released Viruses by qRT-PCR
2.4. Inhibitors
2.5. Infection Experiments
2.6. SDS-PAGE and Western Blotting
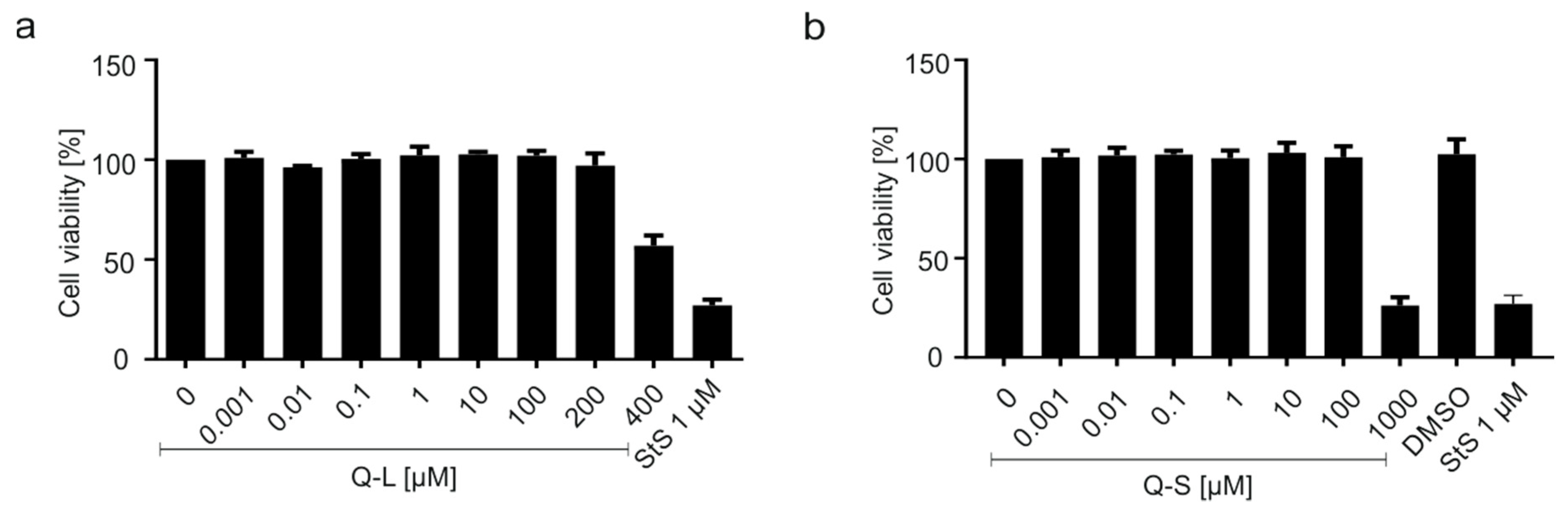
2.7. Assessment of Cell Viability
2.8. Quantitative Real-Time PCR
2.9. Software and Statistics
3. Results
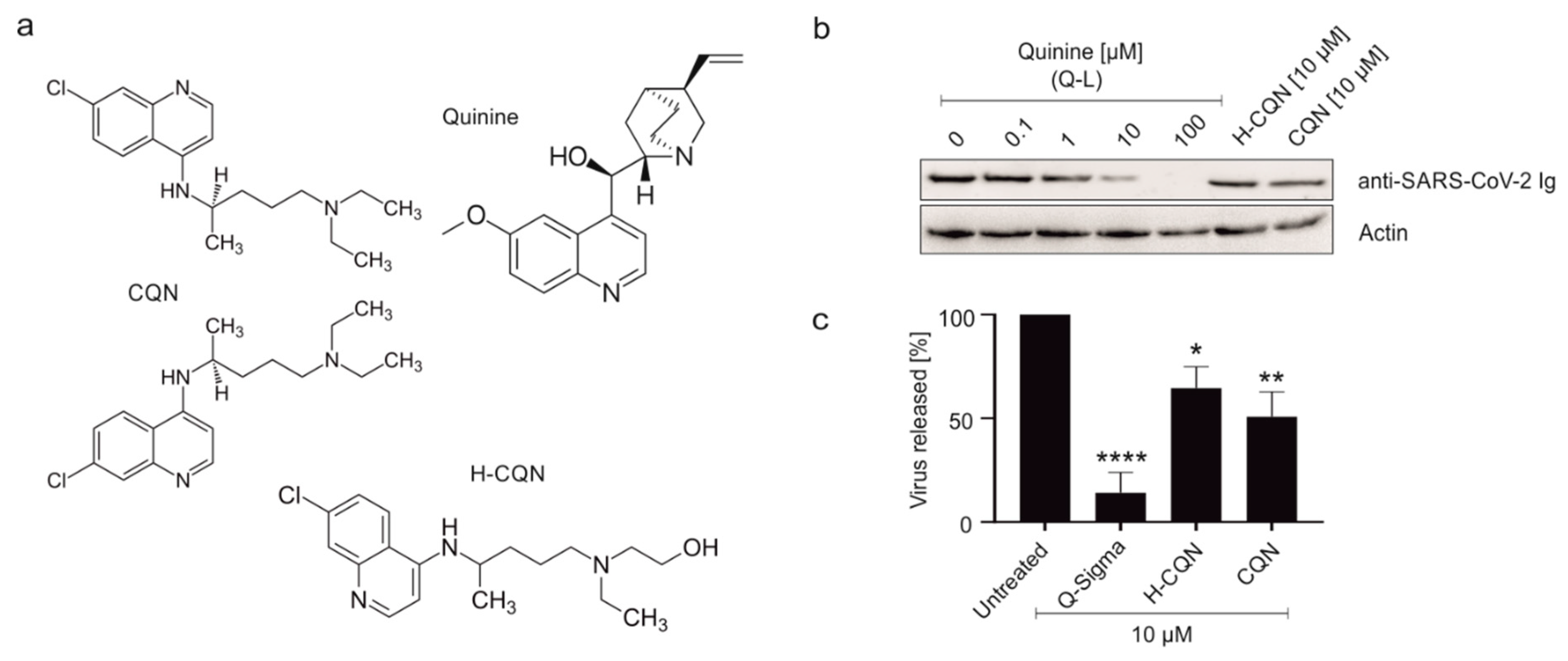
3.1. Quinine Inhibits SARS-CoV-2 More Potently as Compared to H-CQN and CQN in Vero B4 Cells
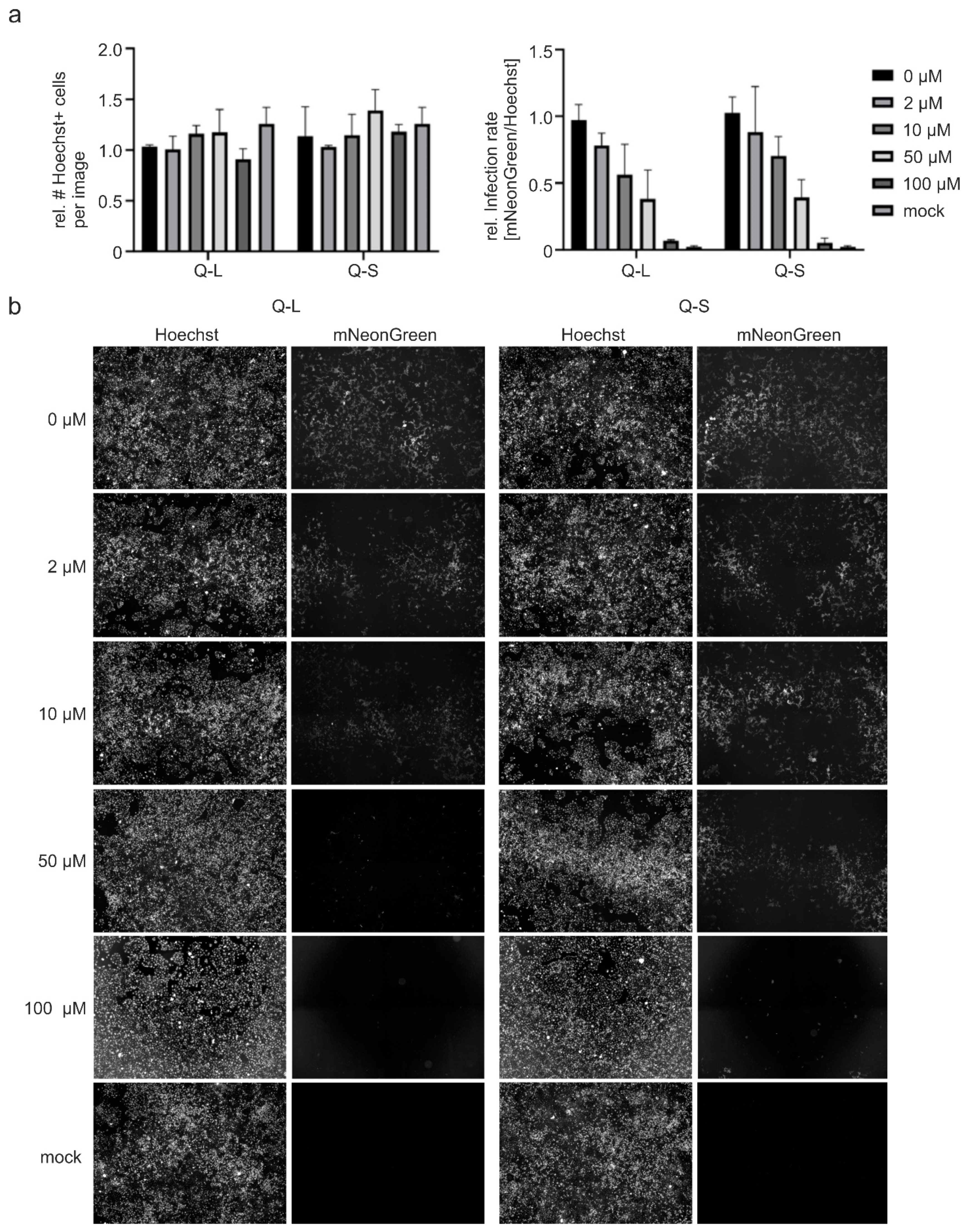
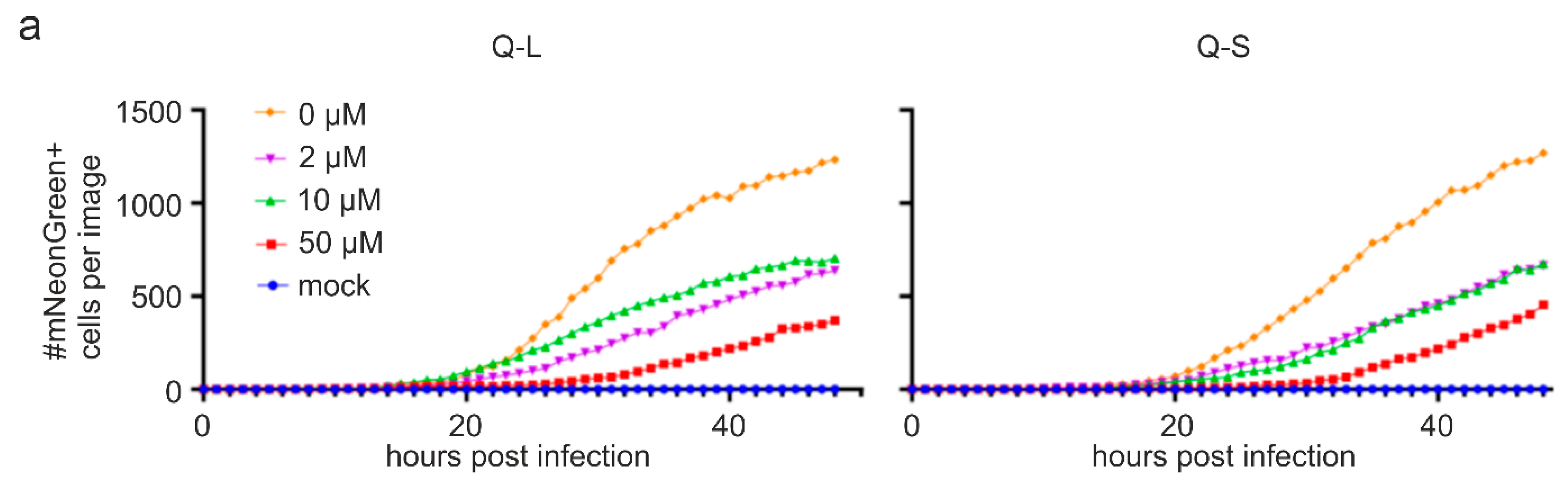
3.2. Quinine Inhibits SARS-CoV-2 Infection and Spread in Human TMPRSS2+ Colon Cells
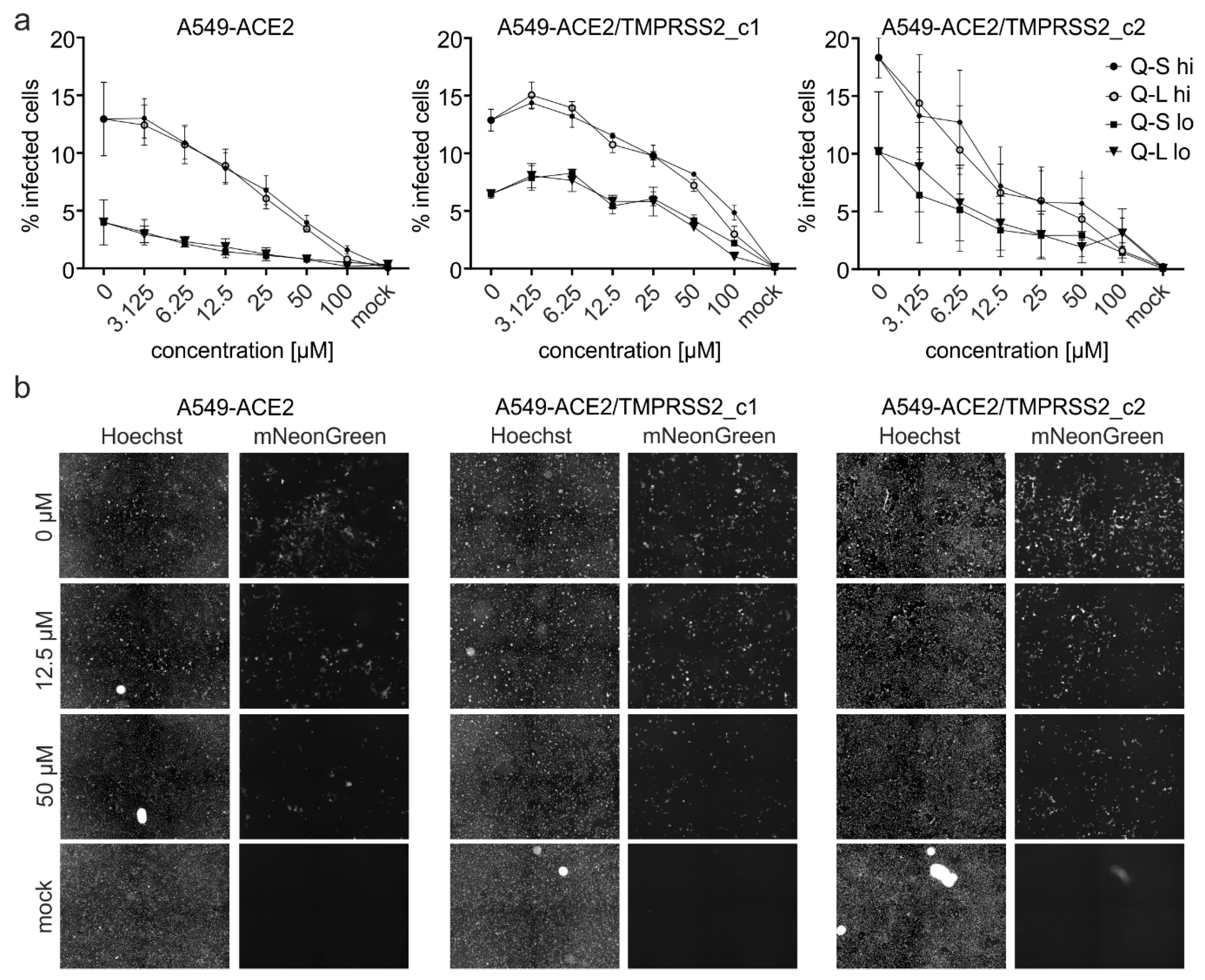
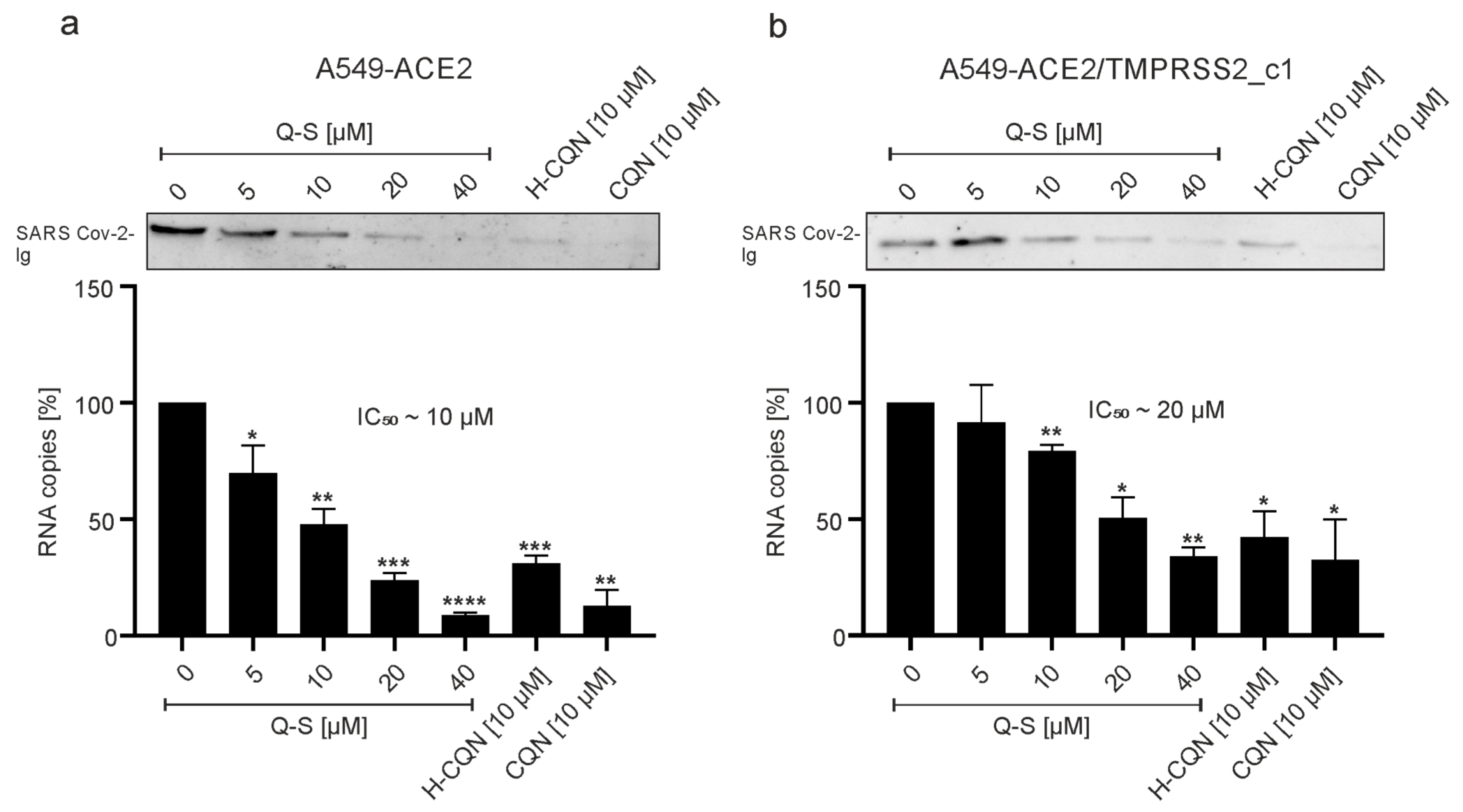
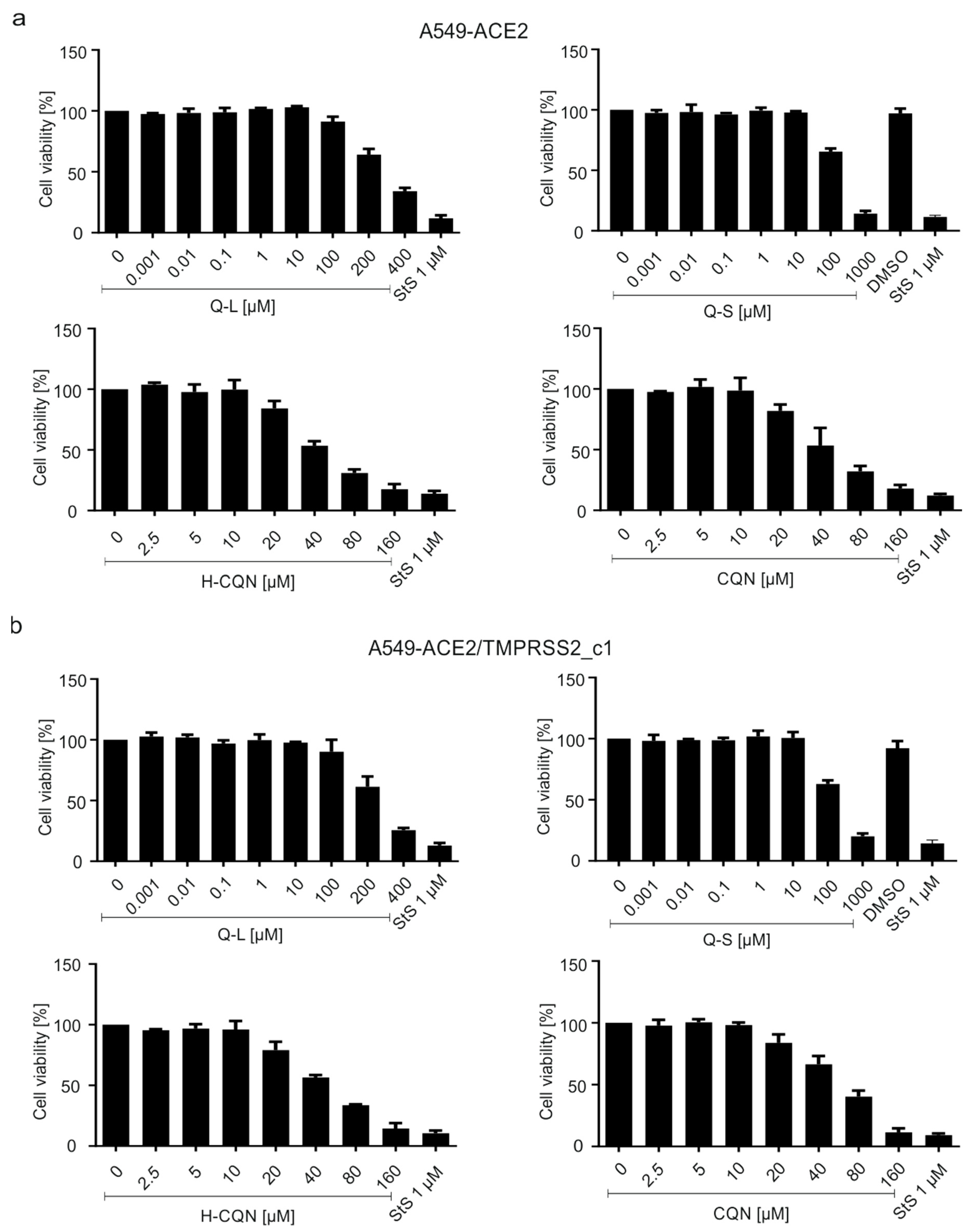
3.3. Quinine Restricts Viral Infection in Human Transgenic Lung Cancer Cells
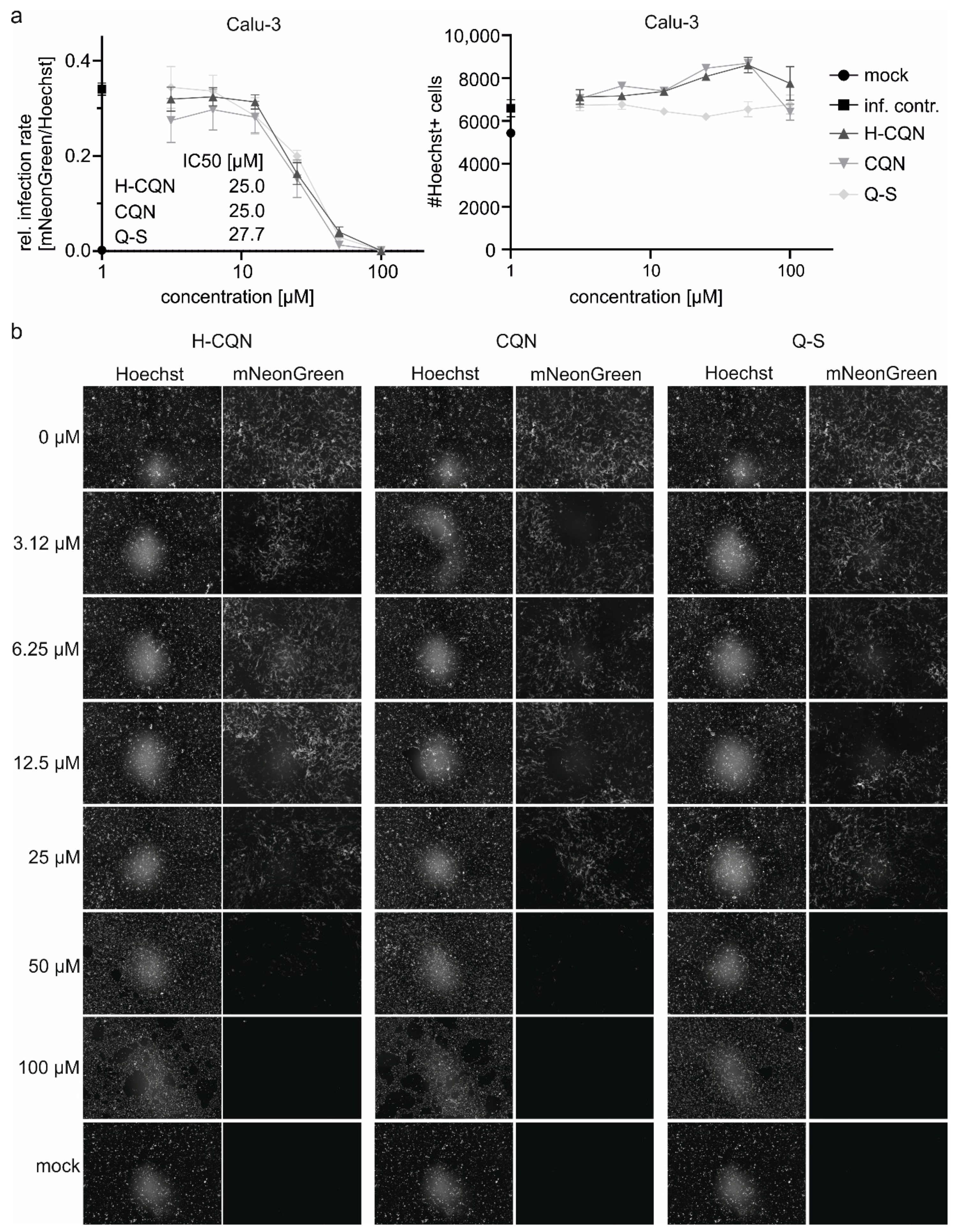
3.4. Quinine Inhibits SARS-CoV-2 Infection in Human Calu-3 Lung Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- World Health Organisation. Q&A: Influenza and COVID-19-Similarities and Differences. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/question-and-answers-hub/q-a-detail/q-a-similarities-and-differences-covid-19-and-influenza (accessed on 19 May 2020).
- Johns Hopkins University. COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University. Available online: https://0-coronavirus-jhu-edu.brum.beds.ac.uk/map.html (accessed on 30 October 2020).
- U. S. Food & Drug Administration. FDA Approves First Treatment for COVID-19. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-treatment-covid-19 (accessed on 30 October 2020).
- Pan, H.; Peto, R.; Henao-Restrepo, A.M.; Preziosi, M.P.; Sathiyamoorthy, V.; Abdool Karim, Q.; Alejandria, M.M.; Hernández García, C.; Kieny, M.P.; Malekzadeh, R.; et al. Repurposed Antiviral Drugs for Covid-19-Interim WHO Solidarity Trial Results. N. Engl. J. Med. 2021, 384, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Wang, D.; Nie, J.; Liang, H.; Gu, J.; Zhao, A.; Xu, L.; Lang, C.; Cui, X.; Guo, X.; et al. The efficacy assessment of convalescent plasma therapy for COVID-19 patients: A multi-center case series. Signal Transduct. Target. Ther. 2020, 5, 219. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Nirula, A.; Heller, B.; Gottlieb, R.L.; Boscia, J.; Morris, J.; Huhn, G.; Cardona, J.; Mocherla, B.; Stosor, V.; et al. SARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, M.; De Giglio, M.A.R.; Roviello, G.N. SARS-CoV-2: Recent Reports on Antiviral Therapies Based on Lopinavir/Ritonavir, Darunavir/Umifenovir, Hydroxychloroquine, Remdesivir, Favipiravir and Other Drugs for the Treatment of the New Coronavirus. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef]
- RECOVERY Collaborative Group. Effect of Hydroxychloroquine in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Rosenberg, E.S.; Dufort, E.M.; Udo, T.; Wilberschied, L.A.; Kumar, J.; Tesoriero, J.; Weinberg, P.; Kirkwood, J.; Muse, A.; DeHovitz, J.; et al. Association of Treatment with Hydroxychloroquine or Azithromycin With In-Hospital Mortality in Patients With COVID-19 in New York State. JAMA 2020, 323, 2493–2502. [Google Scholar] [CrossRef]
- Tang, W.; Cao, Z.; Han, M.; Wang, Z.; Chen, J.; Sun, W.; Wu, Y.; Xiao, W.; Liu, S.; Chen, E.; et al. Hydroxychloroquine in patients with mainly mild to moderate coronavirus disease 2019: Open label, randomised controlled trial. BMJ 2020, 369, m1849. [Google Scholar] [CrossRef]
- Magagnoli, J.; Narendran, S.; Pereira, F.; Cummings, T.H.; Hardin, J.W.; Sutton, S.S.; Ambati, J. Outcomes of Hydroxychloroquine Usage in United States Veterans Hospitalized with COVID-19. Med 2020, 1, 114–127.e3. [Google Scholar] [CrossRef] [PubMed]
- Maisonnasse, P.; Guedj, J.; Contreras, V.; Behillil, S.; Solas, C.; Marlin, R.; Naninck, T.; Pizzorno, A.; Lemaitre, J.; Gonçalves, A.; et al. Hydroxychloroquine use against SARS-CoV-2 infection in non-human primates. Nature 2020, 585, 584–587. [Google Scholar] [CrossRef]
- Kaptein, S.J.F.; Jacobs, S.; Langendries, L.; Seldeslachts, L.; ter Horst, S.; Liesenborghs, L.; Hens, B.; Vergote, V.; Heylen, E.; Barthelemy, K.; et al. Favipiravir at high doses has potent antiviral activity in SARS-CoV-2−infected hamsters, whereas hydroxychloroquine lacks activity. Proc. Natl. Acad. Sci. USA 2020, 117, 26955–26965. [Google Scholar] [CrossRef] [PubMed]
- Ou, T.; Mou, H.; Zhang, L.; Ojha, A.; Choe, H.; Farzan, M. Hydroxychloroquine-mediated inhibition of SARS-CoV-2 entry is attenuated by TMPRSS2. PLoS Pathog. 2021, 17, e1009212. [Google Scholar] [CrossRef]
- Hoffmann, M.; Mösbauer, K.; Hofmann-Winkler, H.; Kaul, A.; Kleine-Weber, H.; Krüger, N.; Gassen, N.C.; Müller, M.A.; Drosten, C.; Pöhlmann, S. Chloroquine does not inhibit infection of human lung cells with SARS-CoV-2. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeier, E.; Dörner, T. Mechanisms of action of hydroxychloroquine and chloroquine: Implications for rheumatology. Nat. Rev. Rheumatol. 2020, 16, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.N.; Wu, Z.X.; Dong, S.; Yang, D.H.; Zhang, L.; Ke, Z.; Zou, C.; Chen, Z.S. Chloroquine and hydroxychloroquine in the treatment of malaria and repurposing in treating COVID-19. Pharmacol. Ther. 2020, 216, 107672. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, S.; Scaccabarozzi, D.; Signorini, L.; Perego, F.; Ilboudo, D.P.; Ferrante, P.; Delbue, S. The Use of Antimalarial Drugs against Viral Infection. Microorganisms 2020, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savarino, A.; Boelaert, J.R.; Cassone, A.; Majori, G.; Cauda, R. Effects of chloroquine on viral infections: An old drug against today’s diseases? Lancet Infect. Dis. 2003, 3, 722–727. [Google Scholar] [CrossRef]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine inhibits autophagic flux by decreasing autophagosome-lysosome fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef] [PubMed]
- Srinivasa, A.; Tosounidou, S.; Gordon, C. Increased Incidence of Gastrointestinal Side Effects in Patients Taking Hydroxychloroquine: A Brand-related Issue? J. Rheumatol. 2017, 44, 398. [Google Scholar] [CrossRef]
- Jorge, A.; Ung, C.; Young, L.H.; Melles, R.B.; Choi, H.K. Hydroxychloroquine retinopathy-implications of research advances for rheumatology care. Nat. Rev. Rheumatol. 2018, 14, 693–703. [Google Scholar] [CrossRef]
- Tönnesmann, E.; Kandolf, R.; Lewalter, T. Chloroquine cardiomyopathy-a review of the literature. Immunopharmacol. Immunotoxicol. 2013, 35, 434–442. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. Bull. Eur. Sur Les Mal. Transm. = Eur. Commun. Dis. Bull. 2020, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Xie, X.; Muruato, A.; Lokugamage, K.G.; Narayanan, K.; Zhang, X.; Zou, J.; Liu, J.; Schindewolf, C.; Bopp, N.E.; Aguilar, P.V.; et al. An Infectious cDNA Clone of SARS-CoV-2. Cell Host Microbe 2020, 27, 841–848.e3. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.; Glowacka, I.; Blazejewska, P.; Soilleux, E.; Allen, P.; Danisch, S.; Steffen, I.; Choi, S.Y.; Park, Y.; Schneider, H.; et al. TMPRSS2 and TMPRSS4 facilitate trypsin-independent spread of influenza virus in Caco-2 cells. J. Virol. 2010, 84, 10016–10025. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Giard, D.J.; Aaronson, S.A.; Todaro, G.J.; Arnstein, P.; Kersey, J.H.; Dosik, H.; Parks, W.P. In vitro cultivation of human tumors: Establishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Inst. 1973, 51, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, J.A.; Tremblay, B.J.; Mansfield, M.J.; Woody, O.; Lobb, B.; Banerjee, A.; Chandiramohan, A.; Tiessen, N.; Cao, Q.; Dvorkin-Gheva, A.; et al. Gene expression and in situ protein profiling of candidate SARS-CoV-2 receptors in human airway epithelial cells and lung tissue. Eur. Respir. J. 2020, 56. [Google Scholar] [CrossRef]
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hu, J.; Zhang, Z.; Jiang, S.; Han, S.; Yan, D.; Zhuang, R.; Hu, B.; Zhang, Z. Efficacy of hydroxychloroquine in patients with COVID-19: Results of a randomized clinical trial. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Cortegiani, A.; Ingoglia, G.; Ippolito, M.; Giarratano, A.; Einav, S. A systematic review on the efficacy and safety of chloroquine for the treatment of COVID-19. J. Crit. Care 2020. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Tian, Z.; Yang, X. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci. Trends 2020, 14, 72–73. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Li, C.; Chen, P.; Zhou, N.; Wang, L.; Li, J.; Jiang, H.; Wang, D.W. Low dose of hydroxychloroquine reduces fatality of critically ill patients with COVID-19. Sci. China Life Sci. 2020. [Google Scholar] [CrossRef]
- Gautret, P.; Lagier, J.C.; Parola, P.; Hoang, V.T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Lagier, J.-C.; Gautret, P.; Colson, P.; Fournier, P.-E.; Amrane, S.; Hocquart, M.; Mailhe, M.; Esteves-Vieira, V.; Doudier, B.; et al. Early treatment of COVID-19 patients with hydroxychloroquine and azithromycin: A retrospective analysis of 1061 cases in Marseille, France. Travel Med. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Mahévas, M.; Tran, V.T.; Roumier, M.; Chabrol, A.; Paule, R.; Guillaud, C.; Fois, E.; Lepeule, R.; Szwebel, T.A.; Lescure, F.X.; et al. Clinical efficacy of hydroxychloroquine in patients with covid-19 pneumonia who require oxygen: Observational comparative study using routine care data. BMJ 2020, 369, m1844. [Google Scholar] [CrossRef]
- Geleris, J.; Sun, Y.; Platt, J.; Zucker, J.; Baldwin, M.; Hripcsak, G.; Labella, A.; Manson, D.; Kubin, C.; Barr, R.G.; et al. Observational Study of Hydroxychloroquine in Hospitalized Patients with Covid-19. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Ip, A.; Berry, D.A.; Hansen, E.; Goy, A.H.; Pecora, A.L.; Sinclaire, B.A.; Bednarz, U.; Marafelias, M.; Berry, S.M.; Berry, N.S.; et al. Hydroxychloroquine and Tocilizumab Therapy in COVID-19 Patients—An Observational Study. PLoS ONE 2020. [Google Scholar] [CrossRef]
- Jomsky, M.G.; Nicholas, A.K. Could Low-Dose Quinine Prevent or Treat Coronavirus Infection? EC Pharmacol. Toxicol. Comment. 2020, 8, 62–64. [Google Scholar] [CrossRef]
- Andriyanto, H. Indonesia Trials Quinine as Covid-19 Treatment. Jakarta Globe. Available online: https://jakartaglobe.id/news/indonesia-trials-quinine-as-covid19-treatment (accessed on 17 May 2020).
- Sputnik News. Pre-Clinical Trials Show Mefloquine Stop Coronavirus Cytopathic Effect, Russian Official Says. Available online: https://sputniknews.com/amp/russia/202005151079316649-pre-clinical-trials-show-mefloquine-stop-coronavirus-cytopathic-effect-russian-official-says/ (accessed on 15 May 2020).
- U.S. National Library of Medicine. GLS-1200Topical Nasal Spray to Prevent SARS-CoV-2 Infection (COVID-19) in Health Care Personnel. Available online: https://clinicaltrials.gov/ct2/show/NCT04408183 (accessed on 30 October 2020).
- Gendrot, M.; Andreani, J.; Boxberger, M.; Jardot, P.; Fonta, I.; Le Bideau, M.; Duflot, I.; Mosnier, J.; Rolland, C.; Bogreau, H.; et al. Antimalarial drugs inhibit the replication of SARS-CoV-2: An in vitro evaluation. Travel Med. Infect. Dis. 2020, 37, 101873. [Google Scholar] [CrossRef]
- Achan, J.; Talisuna, A.O.; Erhart, A.; Yeka, A.; Tibenderana, J.K.; Baliraine, F.N.; Rosenthal, P.J.; D’Alessandro, U. Quinine, an old anti-malarial drug in a modern world: Role in the treatment of malaria. Malar. J. 2011, 10, 144. [Google Scholar] [CrossRef] [Green Version]
- Eyal, S. The Fever Tree: From Malaria to Neurological Diseases. Toxins 2018, 10, 491. [Google Scholar] [CrossRef] [Green Version]
- Kunte, R.; Kunwar, R. WHO Guidelines for the treatment of malaria. Med. J. Armed. Forces India 2011, 67, 376. [Google Scholar] [CrossRef] [Green Version]
- Humphries, W. Just the tonic … gin becomes nation’s favourite spirit. The Times, 16 December 2017. [Google Scholar]
- European Union Law. Regulation (EC) No 1334/2008 of the European Parliament and of the Council of 16 December 2008 on Flavourings and Certain Food Ingredients with Flavouring Properties for Use in and on Foods and Amending Council Regulation (EEC) No 1601/91, Regulations (EC) No 2232/96 and (EC) No 110/2008 and Directive 2000/13/EC. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:02008R1334-20190521 (accessed on 26 May 2020).
- Malakar, S.; Sreelatha, L.; Dechtawewat, T.; Noisakran, S.; Yenchitsomanus, P.T.; Chu, J.J.H.; Limjindaporn, T. Drug repurposing of quinine as antiviral against dengue virus infection. Virus Res. 2018, 255, 171–178. [Google Scholar] [CrossRef]
- Wolf, R.; Baroni, A.; Greco, R.; Corrado, F.; Ruocco, E.; Tufano, M.A.; Ruocco, V. Quinine sulfate and HSV replication. Dermatol. Online J. 2003, 9, 3. [Google Scholar]
- Marois, I.; Cloutier, A.; Meunier, I.; Weingartl, H.M.; Cantin, A.M.; Richter, M.V. Inhibition of influenza virus replication by targeting broad host cell pathways. PLoS ONE 2014, 9, e110631. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Fröba, M.; Graf, P.; Große, M.; Grassauer, A.; Auth, J.; Schubert, U.; Prieschl-Grassauer, E. Iota-carrageenan neutralizes SARS-CoV-2 and inhibits viral replication in vitro. PLoS ONE 2021, 16, e0237480. [Google Scholar] [CrossRef]
- Jang, Y.; Shin, H.; Lee, M.K.; Kwon, O.S.; Shin, J.S.; Kim, Y.-I.; Kim, C.W.; Lee, H.-R.; Kim, M. Antiviral activity of lambda-carrageenan against influenza viruses and severe acute respiratory syndrome coronavirus 2. Sci. Rep. 2021, 11, 821. [Google Scholar] [CrossRef]
- Schütz, D.; Conzelmann, C.; Fois, G.; Groß, R.; Weil, T.; Wettstein, L.; Stenger, S.; Zelikin, A.; Hoffmann, T.K.; Frick, M.; et al. Carrageenan containing over-the-counter nasal and oral sprays inhibit SARS-CoV-2 infection of airway epithelial cultures. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2021. [Google Scholar] [CrossRef]
- Efficacy of a Nasal Spray Containing Iota-Carrageenan in the Prophylaxis of COVID-19 Disease in Health Personnel Dedicated to Patients Care With COVID-19 Disease (CARR-COV-02). 2020. Available online: https://clinicaltrials.gov/ct2/show/NCT04521322 (accessed on 27 November 2020).
- ICT Milstein. La Eficacia del Spray Nasal Con Carragenina Para la Prevención del COVID-19 ha Dado Resultados Positivos. Available online: https://milstein.conicet.gov.ar/la-eficacia-del-spray-nasal-con-carragenina-para-la-prevencion-del-covid-19-ha-dado-resultados-positivos/ (accessed on 16 March 2021).
- Hall, A.P.; Czerwinski, A.W.; Madonia, E.C.; Evensen, K.L. Human plasma and urine quinine levels following tablets, capsules, and intravenous infusion. Clin. Pharmacol. Ther. 1973, 14, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Soyinka, J.O.; Onyeji, C.O.; Omoruyi, S.I.; Owolabi, A.R.; Sarma, P.V.; Cook, J.M. Effects of concurrent administration of nevirapine on the disposition of quinine in healthy volunteers. J. Pharm. Pharmacol. 2009, 61, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, A.L.; Koskiahde, V.; Juva, K. Dosage of antimalarial drugs for children with juvenile rheumatoid arthritis and systemic lupus erythematosus. A clinical study with determination of serum concentrations of chloroquine and hydroxychloroquine. Scand. J. Rheumatol. 1974, 3, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Fiechtner, J.J.; Carpenter, J.R.; Brown, R.R.; Stroshane, R.M.; Stecher, V.J. Plasma hydroxychloroquine concentrations and efficacy in rheumatoid arthritis. Arthritis Rheum. 1987, 30, 567–571. [Google Scholar] [CrossRef]
- Compendium.ch. “CHININSULFAT Hänseler Drag 250 mg”, Package Insert. Available online: https://compendium.ch/product/1105690-chininsulfat-hanseler-drag-250-mg/mpro (accessed on 29 May 2020).
- Vanguard. COVID-19: Nigerian Study Finds Chloroquine, Hydroxychloroquine Effective as Prophylaxis. Available online: https://www.vanguardngr.com/2020/06/covid-19-nigerian-study-finds-chloroquine-hydroxychloroquine-effective-as-prophylaxis/amp/ (accessed on 30 October 2020).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence |
|---|---|
| qRT hGAPDH_fwd | tgc acc acc aac tgc tta gc |
| qRT hGAPDH_rev | ggc atg gac tgt ggt cat gag |
| qRT_hACE2 fwd | gat gcc tcc ctg ctc att tg |
| qRT_hACE2 rev | aac ttc tcg gcc tcc ttg aa |
| qRT_hTMPRSS2 fwd | agg acg aga atc ggt gtg tt |
| qRT_hTMPRSS2_ rev | gga tcc gct gtc atc cac ta |
| Compound IC50 [µM] | A549-ACE2 | A549-ACE2/TMPRSS2_c1 | A549-ACE2/hTMPRSS2_c2 | |||
|---|---|---|---|---|---|---|
| hi | lo | hi | lo | hi | lo | |
| Q-S | 29.07 | 5.98 | 50.02 | 52.82 | 7.48 | 3.75 |
| Q-L | 24.42 | 13.35 | 55.82 | 52.86 | 7.50 | 5.58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Große, M.; Ruetalo, N.; Layer, M.; Hu, D.; Businger, R.; Rheber, S.; Setz, C.; Rauch, P.; Auth, J.; Fröba, M.; et al. Quinine Inhibits Infection of Human Cell Lines with SARS-CoV-2. Viruses 2021, 13, 647. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040647
Große M, Ruetalo N, Layer M, Hu D, Businger R, Rheber S, Setz C, Rauch P, Auth J, Fröba M, et al. Quinine Inhibits Infection of Human Cell Lines with SARS-CoV-2. Viruses. 2021; 13(4):647. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040647
Chicago/Turabian StyleGroße, Maximilian, Natalia Ruetalo, Mirjam Layer, Dan Hu, Ramona Businger, Sascha Rheber, Christian Setz, Pia Rauch, Janina Auth, Maria Fröba, and et al. 2021. "Quinine Inhibits Infection of Human Cell Lines with SARS-CoV-2" Viruses 13, no. 4: 647. https://0-doi-org.brum.beds.ac.uk/10.3390/v13040647